


記住我
African swine fever (ASF), caused by the African swine fever virus (ASFV), is a severe animal disease, with mortality approaching 100% for virulent strains. ASFV, the only known DNA arbovirus (Hakizimana et al., 2020), is also the only member of the ASFV genus of the family Asfarviridae (Mebus, 1988).
ASF has existed for over 100 years, and regional outbreaks have been reported (Mighell and Ward, 2021; Penrith and Kivaria, 2022). ASFV was first reported in Kenya in 1921. Subsequently, it spread to countries in Western Europe, Latin America, and Eastern Europe (Gaudreault et al., 2020). By the mid-1990s, ASFV had been eradicated in the Americas and Europe, but Sardinia remained endemic (Costard et al., 2009; Gaudreault et al., 2020). In 2014, ASFV spread into the Russian Federation and Eastern Europe, where it continues to circulate (Vergne et al., 2016). An outbreak of ASF has occurred in Asia. In August 2018, the first ASF case in China was identified in Shenyang, Liaoning Province. Subsequently, outbreaks were reported in other provinces in China, thus causing extensive economic losses (Tao et al., 2020). Currently, the spread of the ASF continues with outbreaks in Southeast Asia including Thailand, the Philippines, and other countries (Cooper et al., 2021; Cadenas-Fernández et al., 2022; Iscaro et al., 2022; Ramirez-Medina et al., 2022a). ASFV infections of wild boar are more serious than infections of domestic pigs (O’Neill et al., 2020; Sauter-Louis et al., 2021). Because no effective ASF vaccine is available, the prevention and control of ASF is primarily at the level of biosecurity, employing multiple lines of defense that block the spread of ASF (Liu et al., 2021). These approaches are less than optimal and improvements in prevention and control of the virus, as well as the development of an ASF vaccine, are essential. It is therefore necessary to identify the structure and function of ASFV proteins.
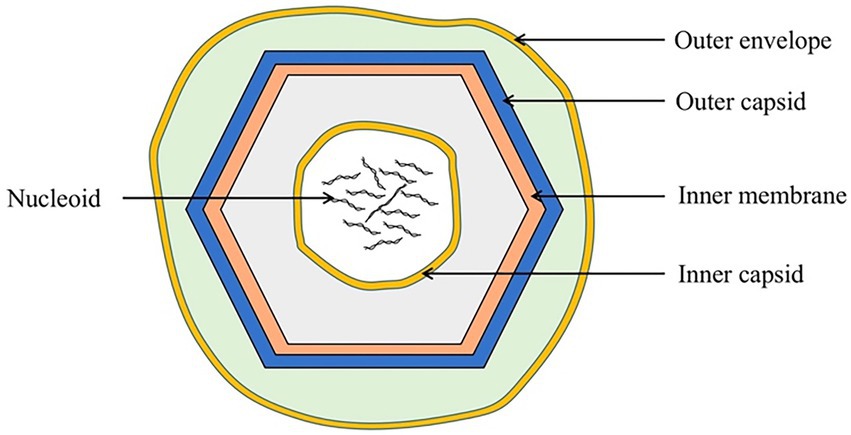
2. A brief description of ASFVASFV is a double-stranded DNA virus (Hakobyan et al., 2018) with a diameter of approximately 260–300 nm. It includes an envelope, capsid, inner membrane, core shell, and genome (Figure 1; Revilla et al., 2018).
Figure 1. Schematic diagram of the ASFV structure. ASFV is composed of an external envelope, outer capsid, inner membrane, inner capsid, and nucleoid.
The external envelope, the outermost structure of ASFV, may be involved in viral attachment and endocytosis. ASFV enters host cells through clathrin-mediated endocytosis (CME) and macropinocytosis (Mercer et al., 2010; Galindo et al., 2015; Zhang et al., 2021). CD2v, an envelope protein, is composed of a signal peptide, transmembrane region, and two immunoglobulin-like domains. CD2v has a variable number of proline-rich repeats within the cytoplasmic domain that can interact with the actin-binding adaptor protein, SH3P7, to promote vesicle transport and signal transduction (Goatley and Dixon, 2011; Jia et al., 2017; Alejo et al., 2018).
The capsid has an icosahedral structure and is adjacent to the external envelope membrane, which protects the virus from nucleases or other physical and chemical factors within the environment. The major capsid protein, p72, is the most dominant structural component of the virion and constitutes 31%–33% of the total mass of the virion (Andrés et al., 2020). This major viral protein induces an antibody response after viral infection. Owing to its conservation and immunogenicity, it is widely used as an antibody detection target for ASFV infection (Geng et al., 2022; Wang C. et al., 2022).
The inner membrane contains an icosahedral capsid that encloses the core shell and the nucleoid. Inside the inner envelope are p17, pE183L, p12, pE248R, and pH108R (Alejo et al., 2018). The inner membrane protein, p17, is an essential and highly abundant protein required for the assembly of the capsid and for icosahedral morphogenesis (Liu et al., 2019; Andrés et al., 2020).
The correct assembly of the core shell depends on the assembly of the outer capsid and the inner membrane, but not vice versa. The core shell may serve to confine and protect the viral genome from host nucleases as well as from host dsDNA sensors, inhibiting the innate immune response during initial infection (Andrés et al., 2020).
The viral genome is enclosed by a core shell, approximately 170 to 194 kbp in length, ending in hairpin loops, comprised variable regions that contain tandem repeats and multigene families (Shimmon et al., 2021). After ASFV endocytosis, the viral genome is delivered into the cytosol, wherein transcription of the early viral genes is initiated (Cackett et al., 2020).
The ASFV genome encodes over 60 structural proteins and 100 non-structural proteins (Jia et al., 2017). Of these, approximately 50 genes encode viral structural proteins with known functions (Sun et al., 2021), although the function of many other proteins is unknown. The mechanism of ASFV infection is not completely understood, which makes vaccine development difficult. Structural and functional analysis of these proteins is essential to the future development of antiviral drugs and vaccines. In this review, recent analysis of ASFV protein structure and function is summarized.
3. ASFV protein structure and function 3.1. Proteins involved in invasionThe entry of ASFV into host cells occurs through receptor-mediated endocytosis; however, the cellular receptors for binding and entry are unknown. Clathrin-mediated endocytosis (CME) or macropinocytosis is the most likely route of endocytosis (Mercer et al., 2010; Galindo et al., 2015; Zhang et al., 2021). This process requires dynein. Viruses use dynein to facilitate internalization and intracellular transport (Hernáez et al., 2010). Previous studies have shown that p54 interacts with dynein and consequently participates in viral invasion (Alonso et al., 2001). Moreover, pE248R participates in membrane fusion, thus promoting ASFV infection; however, the mechanism is currently unclear (Rodríguez et al., 2009). Therefore, the structural analysis of p54 and pE248R is expected to provide a deeper understanding of ASFV invasion.
3.1.1. p54p54, encoded by the E183L gene, is a transmembrane protein located on the external envelope (Rodriguez et al., 1994; Rodríguez et al., 2004). It contains a putative transmembrane domain and disulfide-linked homodimers (Rodríguez et al., 2004). The 13-amino-acid domain of p54 binds LC8, which is part of the microtubular motor complex, and subsequently hijacks the microtubule motor complex cellular transport machinery (Alonso et al., 2001). After this binding, p54 transports the virus intracellularly along microtubules. The virus rapidly reaches the perinuclear area via transport through the cytosol. Subsequently, viral replication occurs. In this process, the disruption of the function of the microtubule motor dynein inhibits the transport of viruses (Hernáez et al., 2010). This interaction is critical during viral internalization and transport to factory sites (Hernáez et al., 2004). In addition, the interaction of the microtubule motor protein LC8 with p54 protein may allow viruses to use host cells to promote viral transport and replication (Alonso et al., 2001). However, owing to the complexity of the system, the mechanism of this interaction remains elusive (Hernáez et al., 2010). Therefore, analysis of the structure of p54 may facilitate understanding of this mechanism of interaction, and enable blocking of the critical early stage of ASFV infection.
3.1.2. pE248RpE248R, encoded by the E248R gene, is localized at the inner envelope and contains intramolecular disulfide bonds, a putative myristoylation site, and a hydrophobic transmembrane region near its carboxy terminus (Rodríguez et al., 2009). pE248R is associated with the infectivity of ASFV. A lack of pE248R decreases the infectivity of ASFV. In addition, pE248R is required for cell entry and membrane fusion (Rodríguez et al., 2009). pE248R shows high amino acid sequence similarity with VACV protein L1—a component of the poxviral multiprotein entry/fusion complex, which is required for proper membrane fusion and core penetration (Moss, 2012; Hernáez et al., 2016). Membrane fusion may occur during the passage of virions through the cytoplasmic vacuolar membrane and subsequently deliver viral “cores” into the cytoplasm (Valdeira et al., 1998; Rodríguez et al., 2009). In addition, ASFV entry relies on a fusion machinery comprising pE248R and pE199L (Matamoros et al., 2020). Further studies on this protein and fusion machinery may reveal the mechanisms of ASFV entry and postentry pathways.
3.2. Proteins involved in replication and transcriptionASFV replication occurs mainly in the cytoplasm 6–8 h after host cell infection. The expression of intermediate and late genes, which encode viral particle structure-related proteins and early transcription factors, starts after 8–16 h (Gaudreault et al., 2020; Wang Y. et al., 2021). The proteins currently known to be involved in ASFV replication and transcription are shown in Table 1.
Table 1. Proteins involved in the replication and transcription of ASFV.
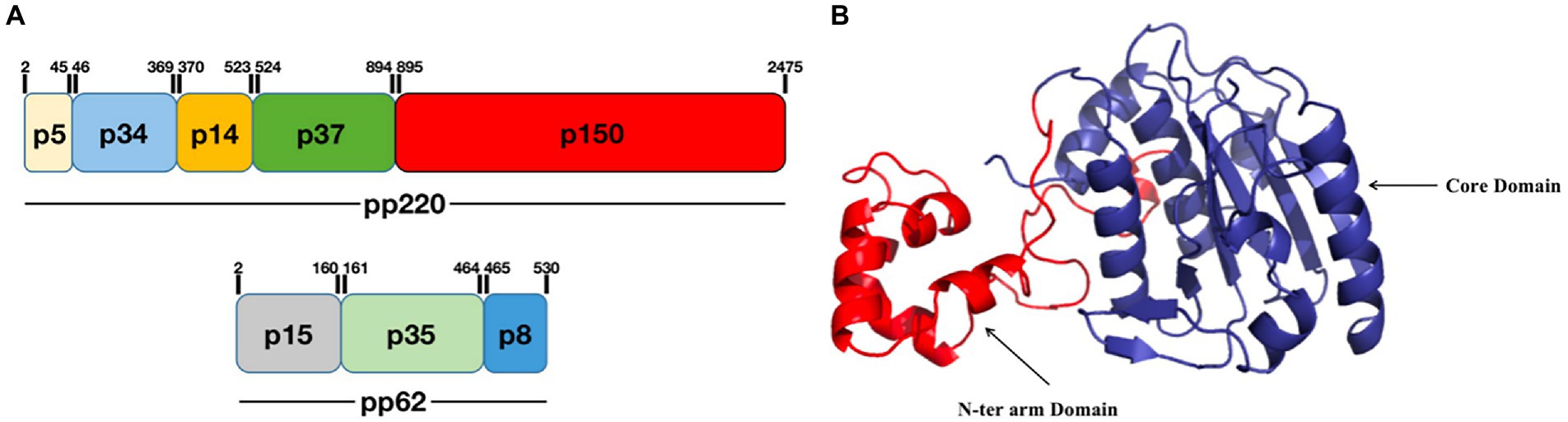
3.2.1. pS273R catalyzes the pp220 polyprotein precursors into p14 and p37 that are involved in replicationpS273R is a specific SUMO-1 cysteine protease that catalyzes the maturation of the pp220 and pp62 polyprotein precursors into the core-shell proteins p5, p34, p14, p37, p150, p15, p35, and p8 (Figure 2A; Andrés et al., 2002a; Alejo et al., 2018). The pS273R protease comprises 12e α-helices, seven β-strands, and one 310-helix, which fold into two close but distinct domains: the core domain and arm domain (Figure 2B). The core domain consists of a sandwich-like fold structure that contains residues N84 to A273 and shares high structural similarity with chlamydial deubiquitinating enzyme, sentrin-specific protease, and adenovirus protease. The arm domain is unique to ASFV, and consists of five α-helices (α1 to α5) and one 310 helix; residues M1 to N83 of pS273R are necessary for enzymatic activity. The pS273R protease has a catalytic triad, C232-H168-N187, which catalyzes ubiquitin or ubiquitin-like proteins such as ISG15 or SUMO (Báez-Santos et al., 2015; Li et al., 2020). The pS273R protease may antagonize the host IFN-I pathway by deubiquitinating specific proteins (Li et al., 2020). Furthermore, targeted inhibitors have been designed for pS273R and include tetrapeptide nitrile compounds (Mac Sweeney et al., 2014; Grosche et al., 2015; Li et al., 2020). Evaluation of the biochemical activity of pS273R has demonstrated that these inhibitors bind the enzyme’s active site, thus providing a reference for the design of antiviral drugs based on the ASFV protein structure.
Figure 2. Schematic diagram of the ASFV pS273R structure. (A) pS273R catalyzes the hydrolysis of pp220 to p5, p34, p14, p37, and p150, and pp62 to p15, p35, and p8. (B) The pS273R protein consists of an “arm domain” (red) and a “core domain” (dark blue; Li et al., 2020).
pp220, encoded by the CP2475L gene, is an N-myristoylated precursor polypeptide that may function as a membrane-anchoring signal that facilitates the binding of the developing core shell to the inner envelope (Andrés et al., 2002a,b). pp220 is essential for core assembly rather than the construction of external viral domains; moreover, it is directly or indirectly involved in the incorporation of other major core components, including the viral genome, into virus particles (Andrés et al., 2002a,b). The processing of pp220 is dependent on the expression of p72. Unprocessed pp220 assembles into aberrant zipper-like elements consisting of an elongated membrane-bound protein structure by the repression of p72 synthesis (Andrés et al., 2002a).
p37 and p14 from the proteolytic processing of pp220 are both core shell proteins (Andrés et al., 2002a). p37, the first nucleocytoplasmic shuttling protein encoded by the ASFV genome, is involved in the nuclear transport of viral DNA during the ASFV replication cycle (Eulálio et al., 2004). Studies have shown that p37 is both imported into the nucleus and exported from the nucleus to the cytoplasm through a process mediated by CRM-1 receptors (Eulálio et al., 2007). p14 is also a nucleocytoplasmic shuttling protein. However, unlike p37, it only enters the nucleus but is not exported to the cytoplasm. p37 and p14 may be involved in the formation of a complex with viral DNA, thus facilitating entry into the nucleus and subsequent initiation of DNA replication (Eulálio et al., 2004).
3.2.2. The viral nucleoid protein p10 and pA104R involved in replicationp10, encoded by the K78R gene, is located in the viral nucleoid and has been shown to exhibit DNA-binding activities that may be related to the characteristic helix-turn-helix structural motif (Nunes-Correia et al., 2008). Both the C-terminal helix rich in lysine residues and the serine-rich residues found in the N-terminal helix of p10 are crucial for the interaction with dsDNA. p10 exploits cellular mechanisms to achieve active nuclear import (Istrate et al., 2022). In ASFV-infected cells, p10 protein strongly accumulates in the nucleus in late stages post-infection and therefore may perform an important function in the nucleus during the late phase of the viral replication cycle (Nunes-Correia et al., 2008). The crystal structure of p10 has not yet been reported. Thus, further study of the structure of p10 may aid in understanding of the binding of p10 to DNA in the viral life cycle, and provide a theoretical basis for the development of an ASF vaccine.
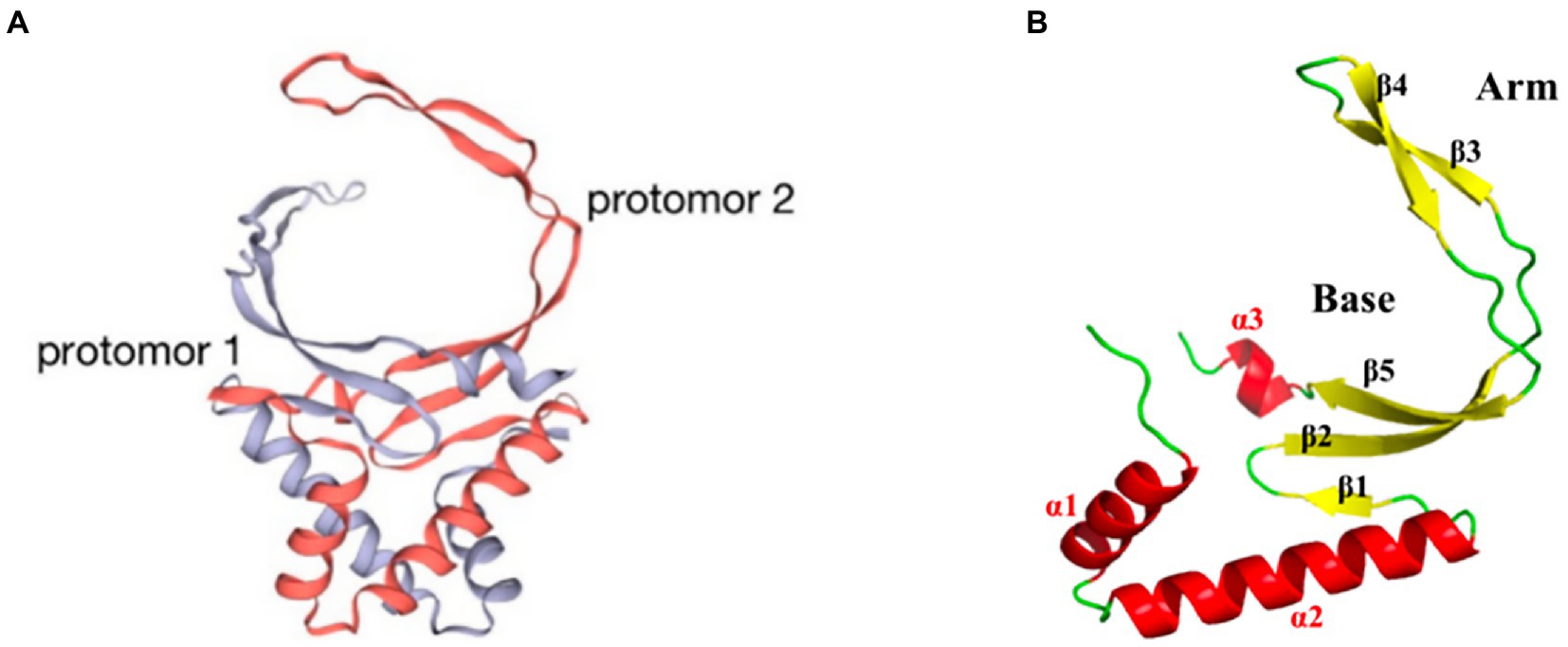
pA104R is a homodimer with two domains: an α-helical region (AHR) and a β-strand DNA-binding region (BDR; Figure 3A). The secondary structural elements of the protomer of the pA104R dimer include two α-helixes (α1 and α2) followed by five β-sheets (β1-5), which end with a final α/310-helix (α3/η1). The β1, β2, and β5 strands form an antiparallel β-sheet, which constitutes the bottom of the BDR, whereas the β3 and β4 strands compose the arms of the BDR (Figure 3B; Liu et al., 2020). The positively charged amino acids K92, R94, K97, R69, H72, K83, and K85 are densely distributed within the BDR base and the BDR arm regions, and serve as potential binding sites for the negatively charged DNA backbone. Residues in the BDR base region are important contributors to DNA interaction (Liu et al., 2020; Urbano and Ferreira, 2020). pA104R has been suggested to bind dsDNA and ssDNA, with a higher affinity for the former. Furthermore, stilbene derivative compounds bind pA104R and inhibit the pA104R-DNA interaction, thus preventing ASFV replication (Liu et al., 2020). pA104R also participates in the modulation of viral DNA topology and genome packaging, and therefore is likely to be involved in viral DNA replication, transcription, and packaging (Frouco et al., 2017). ASFV DNA packaging is a highly dynamic process, and how pA104R and DNA participate in the process is currently unclear. Future studies on pA104R are expected to provide a deeper understanding of ASFV DNA replication and transcription.
Figure 3. Schematic diagram of the ASFV pA104R structure. (A) The dimer structure of the pA104R protein. Gray: protomer 1; light red: protomer 2. (B) Distribution of α-helices and β-sheets in the pA104R protomer.
3.2.3. pP1192R involved in replicationpP1192R is a type II DNA topoisomerase (topoII) encoded by ASFV ORF P1192R, which is located in cytoplasm and accumulating in the viral factories (Coelho et al., 2015). TopoIIs are able to relax supercoils, to resolve knots and tangles in the DNA (Nitiss, 2009). Thus, pP1192R may play an active role in the replication of ASFV genome. And pP1192R can alleviate the topological tension that generated by the binding of pA104R to the DNA (Coelho and Leitão, 2020). In addition, structural analysis of pP1192R and studies with known structures of topoII inhibitors may lead to the discovery of anti-ASFV drugs (Constantinou et al., 1995; Mottola et al., 2013), which can eventually counteract the activity of the viral topoisomerase without hindering the cellular homologs (Coelho and Leitão, 2020).
3.2.4. I73R Protein involved in replicationASFV encodes a 72-amino acid protein, I73R protein (Cackett et al., 2020). I73R protein is expressed in the nucleus in early stages post-infection and gradually translocates from the nucleus to the cytoplasm. This protein has no homology with any known or other functionally identified protein sequence. I73R protein has a classical α/β architecture wherein three α-helices form the core of the domain, and helices 2 and 3 form the helix-turn-helix unit (Sun et al., 2022). I73R protein shares structural similarity with Z-DNA-binding domains (Zα), which are specific for the left-handed conformation of nucleic acid duplexes of DNA, DNA/RNA hybrids, and RNA (Lee et al., 2016). I73R protein also binds CpG repeats DNA duplexes with a high propensity for forming Z-DNA during DNA-binding assays. Key residues of I73R protein are highly conserved in other Zα domain proteins, and behave similarly to those of other Zα domain-containing proteins such as ADAR1, PKZ, and DAI (Ha et al., 2008, 2009; de Rosa et al., 2013). As a member of the Zα family with the ability to bind Z-DNA, I73R protein may be involved in the process of immune evasion (Sun et al., 2022). I73R protein has also been shown to inhibit host gene expression and to play an important role in host-pathogen interactions and nuclear organization during the initial phase of ASFV replication (Sun et al., 2022). However, the mechanism through which I73R protein promotes ASFV infection is unclear, and thus further research is necessary.
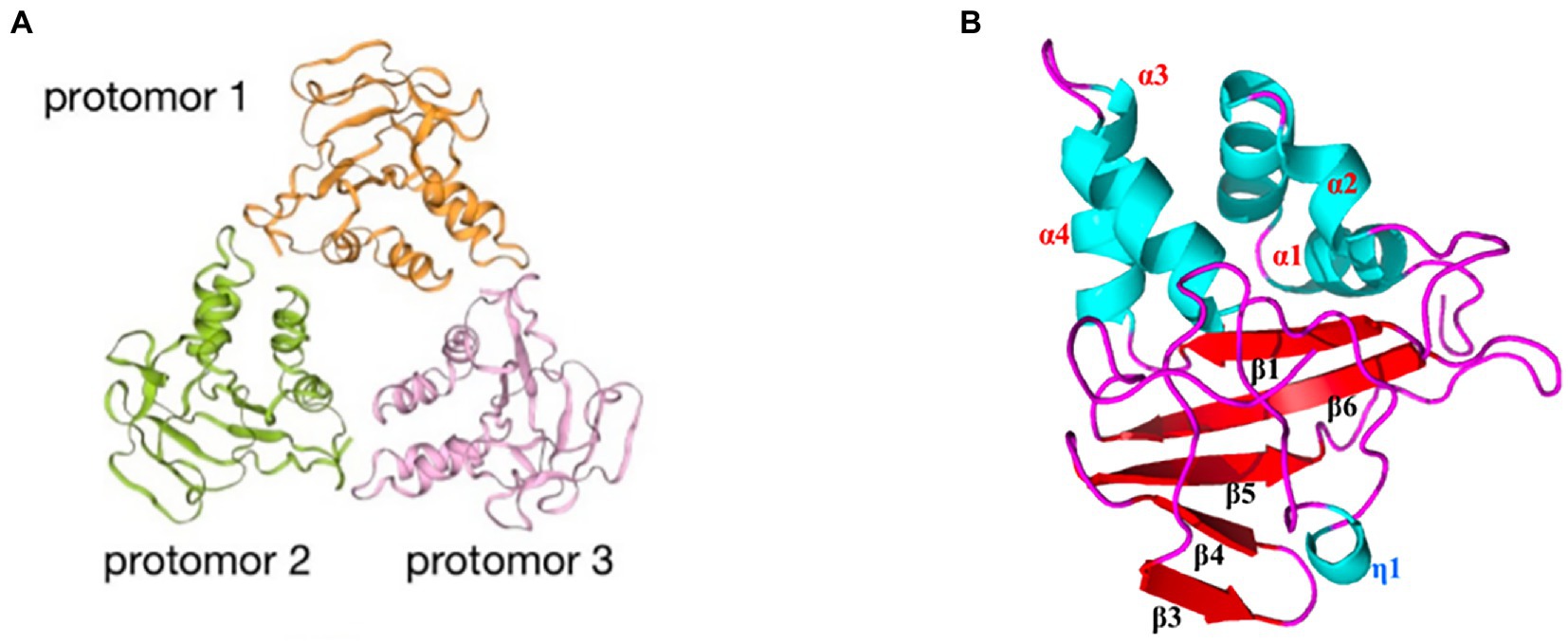
3.2.5. p15 involved in transcriptionASFV p15 is one of the mature products derived from polyprotein pp62 processing at the Gly-Gly-Xaa sequence (Simón-Mateo et al., 1997). The viral polyprotein processing proteinase pS273R plays a key role in this process (Li et al., 2020). p15 exists as several types of oligomers such as dimers and trimers. The dimer (Figure 4A) of p15 arises from disulfide bond formation between the Cys9 and Cys30 residues (Guo et al., 2021). These disulfide bonds are critical for the formation of the p15 structure. The structure of the p15 protomer consists of four α-helixes and six β-sheets (Figure 4B). p15 has been suggested to bind nucleic acids, and to have a stronger affinity for ssDNA than dsDNA. Lys10 and Lys39 are essential for the affinity of DNA binding (Guo et al., 2021; Wang G. et al., 2021). p15’s binding to ssDNA suggests its potential involvement in viral transcription and DNA replication during the ssDNA phase of the viral life cycle (Guo et al., 2021).
Figure 4. Schematic diagram of the ASFV p15 structure. (A) The trimeric structure of the p15 protein. Orange: protomer 1; green: protomer 2; pink: protomer 3. (B) Distribution of α-helices and β-sheets in the p15 proteome.
3.2.6. pQP509L and pQ706L involved in transcriptionpQP509L and pQ706L are both the SF2 RNA helicases that expressed by ASFV (Freitas et al., 2019). pQP509L was detected from 12 h post infection (hpi) within viral factories and host nucleus, and able to assist termination and release of late viral transcripts, which involved in viral transcription. At later times of infection, it can be also related to the modulation of antiviral responses (Freitas et al., 2019). In addition, pQP509L may be associated with ASFV virulence. The deletion of the QP509L and QP383R genes from ASFV is highly attenuated (Li et al., 2022). But subsequent studies have shown that deleting QP509L gene from the highly virulent Georgia 2010 strain is unable to affect the replication or virulence of ASFV (Ramirez-Medina et al., 2022b). pQ706L was detected only at viral factories from 12 hpi onward, and regulates elongation and release of late viral transcripts to involve the transcription of ASFV (Freitas et al., 2019). However, the mechanism of pQP509L and pQ706L is still unclear. Therefore, the research on their structures and functions is expected to enable us to understand these proteins in depth and is crucial for the ASF vaccine design and responding to emerging ASFV strains.
3.3. pNP868R involved in post-transcriptional modificationpNP868R consists of an N-terminal triphosphatase (TPase) domain, a central guanylyltransferase (GTase) domain, and a C-terminal methyltransferase (MTase) domain. The TPase and GTase domains are members of the triphosphate tunnel metalloenzyme (TTM) and covalent nucleotidyltransferase (NTase) superfamilies, respectively (Pena et al., 1993). The MTase domain exhibits the characteristic core fold of class I family of S-adenosyl-l-methionine (AdoMet)-dependent MTases (Cong and Shuman, 1995). The 5′-triphosphate end of the pre-mRNA is first hydrolyzed to a diphosphate by RNA 5′-TPase. The diphosphate RNA is then capped with GMP by RNA GTase forming GpppRNA. Finally, the GpppRNA cap is converted to a 7-methylguanosine RNA cap (m7GpppRNA) by RNA (guanine-N7)-MTase (Figure 5A; Shuman, 2001; Decroly et al., 2011). The process of mRNA 5′-end capping involved by pNP868R contributes to mRNA stability and efficient translation (Du et al., 2020).
Figure 5. (A) The capping mechanism of mRNA involved in pNP868R. First, the triphosphatase (TPase) hydrolyses the γ-phosphate of the nascent RNA (pppNp-RNA, in which N denotes the first transcribed nucleotide and p denotes a phosphate group) to yield a diphosphate RNA (ppNp-RNA) and inorganic phosphate (Pi). Then, guanylyltransferase (GTase) reacts with the α-phosphate of GTP (Gppp), releasing pyrophosphate (PPi) and forming a covalent enzyme–guanylate intermediate (Gp–GTase). The GTase then transfers the GMP molecule (Gp) to the 5′-diphosphate RNA to create GpppNp-RNA. In the final step (guanine-N7)-methyltransferase (N7MTase) transfers the methyl group from S-adenosyl-l-methionine (AdoMet) to the cap guanine to form the cap-0 structure, with the 7-methyl-GpppNp (m7GpppNp) releasing S-adenosyl-l-homocysteine (AdoHcy) as a by-product (Decroly et al., 2011). (B) Schematic diagram of the ASFV pNP868R protein structure, showing α-helices and β-sheets within the structure (Du et al., 2020).
The MTase domain of pNP868R has been resolved and displays a typical class I MTase fold comprised a seven-strand β-sheet (described as β4, β3, α2, β5, β6, β12, and β11) with six helices (α1, α2, α3, α4, α5, and α7). Based on these structures, there is an N-terminal extended β-sheet (β1), followed by a 9-residue β1-α1 loop, and a flap domain between β6 and α7 that contains a four-strand antiparallel β-sheet (β7, β8, β9, and β10), which is flanked by four extra helices (α6, α8, α9, and α10; Figure 5B). The N-terminal extension contains R592 and R597 residues that may be close to the active site upon substrate binding, playing an essential role in substrate recognition as judged by in vitro MTase analysis (Du et al., 2020). Thus, the MTase domain plays a particularly important role in the function of pNP868R.
Future analysis of pNP868R substrate recognition and catalysis are expected to provide a direction for the development of effective biologics targeting ASFV-related enzymes.
3.4. Proteins involved in ASFV translationASFV has self-sufficient replication and transcription mechanisms, but relies on the host translational mechanism for protein synthesis (Zhong et al., 2022). ASFV regulates the DNA translation pathway in a variety of ways, and its products undergo post-translational modifications (Wang Y. et al., 2021). Previous studies have shown that g5Rp, pI215L and pDP71L participate in these processes (Zhang et al., 2010; Dixon et al., 2017; Freitas et al., 2018; Yang et al., 2022), and the structure of g5Rp has been resolved.
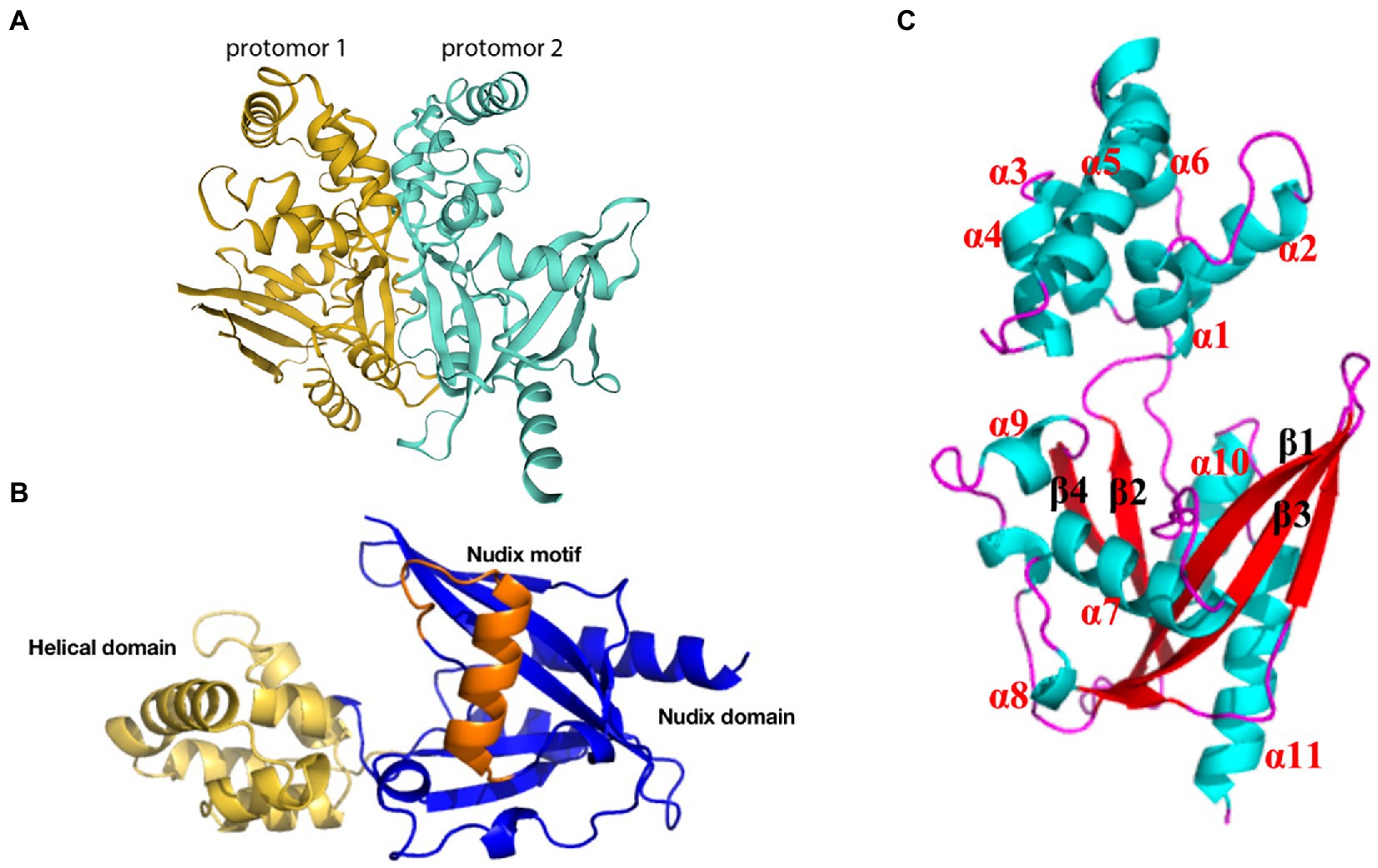
3.4.1. g5Rpg5Rp, the only viral mRNA-decapping enzyme, is expressed in the endoplasmic reticulum at the earliest stage of infection and accumulates throughout the ASFV infection process; this enzyme plays an essential role in the machinery assembly of mRNA regulation and translation initiation. g5Rp forms a stable symmetric dimer of g5Rp crystal packaging, with the dimer composed of two protomers (Figure 6A). Each protomer is composed of a unique N-terminal helical domain and a C-terminal classic Nudix domain (Figure 6B). The helical domain forms a globin-fold-like feature composed of six α-helices (α1 to α6) that connects to the Nudix domain by two hinge linkers. The Nudix domain consists of a central curved β-sheet (β1, β2, β3, β4) surrounded by five α-helices (α7 to α11) and several loops, thereby forming a classic α-β-α sandwich structure (Figure 6C). The helical domain is the major mediator of RNA interaction, with positively charged regions likely contributing to g5Rp binding with RNA (Yang et al., 2022). The g5Rp Nudix domain interacts with RNA, with the g5Rp C-terminal exhibiting substrate selectivity at the RNA binding step (Yang et al., 2022). K8, K95, K133, and R221 are the key residues that mediate g5Rp-RNA interaction and are also important in g5Rp-related cellular RNA degradation in vivo (Yang et al., 2022). g5Rp is the only Nudix hydrolase that targets mRNAs of the host cell. After g5Rp binds to RNA of the host cell, it robustly cleaves the mRNA 5′ cap attached to an RNA moiety dependent upon the Nudix motif (Parrish et al., 2009; Yang et al., 2022). g5Rp has a broader range of nucleotide substrate specificities, including a variety of guanine and adenine nucleotides and dinucleotide polyphosphates (Cartwright et al., 2002). Removal of the 5′ cap on host cellular mRNAs by g5Rp is beneficial to viral gene expression during the early stages of infection (Dixon et al., 2013; Sánchez et al., 2013). As originally described, g5Rp dephosphorylates 5-PP-InsP5 (InsP7) to produce InsP6. InsP6 inhibits the mRNA-decapping activity of g5Rp by competing for the substrate mRNA-binding surface of g5Rp.
Figure 6. Schematic diagram of ASFV g5Rp structure. (A) g5Rp consists of two protomers “back to back,” protomer 1 is in dark yellow and protomer 2 is in cyan (Yang et al., 2022). (B) Nudix domain (dark blue), the helical domain (yellow), and the Nudix motif of g5Rp (orange). (C) The α-helices and β-sheets of the g5Rp protomer.
Although the crystal structure of g5Rp has been resolved, the complex structure formed by the interaction between g5Rp and mRNA is unclear. Therefore, the future resolution of this issue is expected to deepen understanding of the structural basis for g5Rp protease activity, and may have far-reaching significance for the development of new drugs and vaccines against ASFV.
3.4.2. pI215LpI215L, encoded by ASFV I215L gene, localizes in viral factories and host cell nucleus (Freitas et al., 2018), and the only known virus-encoded E2-ubiquitin conjugating enzyme (UBCv1; Barrado-Gil et al., 2020). UBCv1 has high activity and has a catalytic domain that could associate with several classes of polyubiquitin chains. And the catalytic site Cys85 in its catalytic domain plays an important functional role (Freitas et al., 2018; Barrado-Gil et al., 2020). UBCv1 interact with the 40s ribosomal protein S23 (RPS23), which prevents potential mRNAs or other factors from binding to it to influence the translation (Barrado-Gil et al., 2020). UBCv1 is also able to bind to the eukaryotic initiation factor 4E (eIF4E) and induces this factor overexpression, which result in increased protein synthesis (Barrado-Gil et al., 2020; Romagnoli et al., 2021). Besides, the UBCv1 could bind to the E3 ligase Cullin4B (Cul4B) that has an important role in regulating TSC2 and mTOR signaling and degrading 4E-BP2 eukaryotic translation initiation factor (Wang et al., 2013; Kouloulia et al., 2019; Barrado-Gil et al., 2020). The further study on the structure and function of pI215L is expected to the potential use in the drug against ASF.
3.4.3. pDP71LpDP71L exists in either a short form of 70 to 72 amino acids or a long form of approximately 184 amino acids in all ASFV isolates, and is expressed late during the replication cycle (Zhang et al., 2010). It binds all three isoforms (α, β, and γ) of the protein phosphatase 1 catalytic subunit (PP1c) (Rivera et al., 2007; Zhang et al., 2010), and the residues V16 and F18A in pDP71L were critical for binding to PP1c (Barber et al., 2017). DP71L decreases the amount of phosphorylated eIF2α by recruiting PP1c, which dephosphorylates eIF2α, and also inhibits the induction of ATF4 and its downstream target CHOP (Zhang et al., 2010). The phosphorylation of eIF2α is a key rate-limiting step in the control of protein synthesis; therefore, the virus can affect protein expression by decreasing the phosphorylation of eIF2α. And CHOP is essential in causing apoptosis in cells with irrecoverable ER stress (Marciniak et al., 2004; Zhang et al., 2010). Thus, pDP71L inhibits apoptosis by inhibiting CHOP induction and activation (Dixon et al., 2017). However, DP71L is not the only factor required by the virus to control the phosphorylation level of eIF2α during infection (Zhang et al., 2010). Therefore, further studies on the mechanisms affecting the phosphorylation level of eIF2α and the synthesis of protein would deepen understanding of the translation process of ASFV.
3.5. A224L and A179L proteins inhibit the apoptosis of host cells and consequently promote viral proliferation 3.5.1. A224L proteinA224L protein, a late protein in the viral cycle, is a member of the inhibitor of apoptosis (IAP) family that contains a single BIR motif at the NH2 terminus as well as a sequence that may constitute a zinc finger domain of the 4-cysteine type at the C-terminal region. BIRs are involved in all known interactions between IAPs and other proteins and are required for the antideath activity of IAPs (Deveraux et al., 1997; Harvey et al., 1997). However, A224L protein differs in that it lacks the typical RING motif found in other IAP proteins. Therefore, A224L protein may not have ubiquitin ligase activity (Nogal et al., 2001). It has been suggested that A224L protein promotes cell survival. A224L protein substantially inhibits caspase 3 activity and cell death induced by treatment with tumor necrosis factor α (Nogal et al., 2001). The means by which A224L inhibits cell death was suggested from studies indicating that transient expression of A224L protein activates the NF-kB-dependent reporter C-rel. This NF-kB inducing activity was abrogated by an IKK-2-dominant negative mutant and enhanced by expression of TNF receptor-associated factor 2 (TNF-R2) (Rodríguez et al., 2002). The activation of NF-kB mediated by TNF-R2 can inhibit apoptotic cell death by activating transcription of a number of anti-apoptotic genes including IAP and Bcl-2 family members (Rodríguez et al., 2002; Dixon et al., 2017).
3.5.2. A179L proteinA179L protein, similarly to pDP71L and A224L proteins, suppresses ASFV infection-induced apoptosis. However, among these proteins, only the crystal structure of A179L protein has been solved (Nogal et al., 2001; Banjara et al., 2017; Dixon et al., 2017).
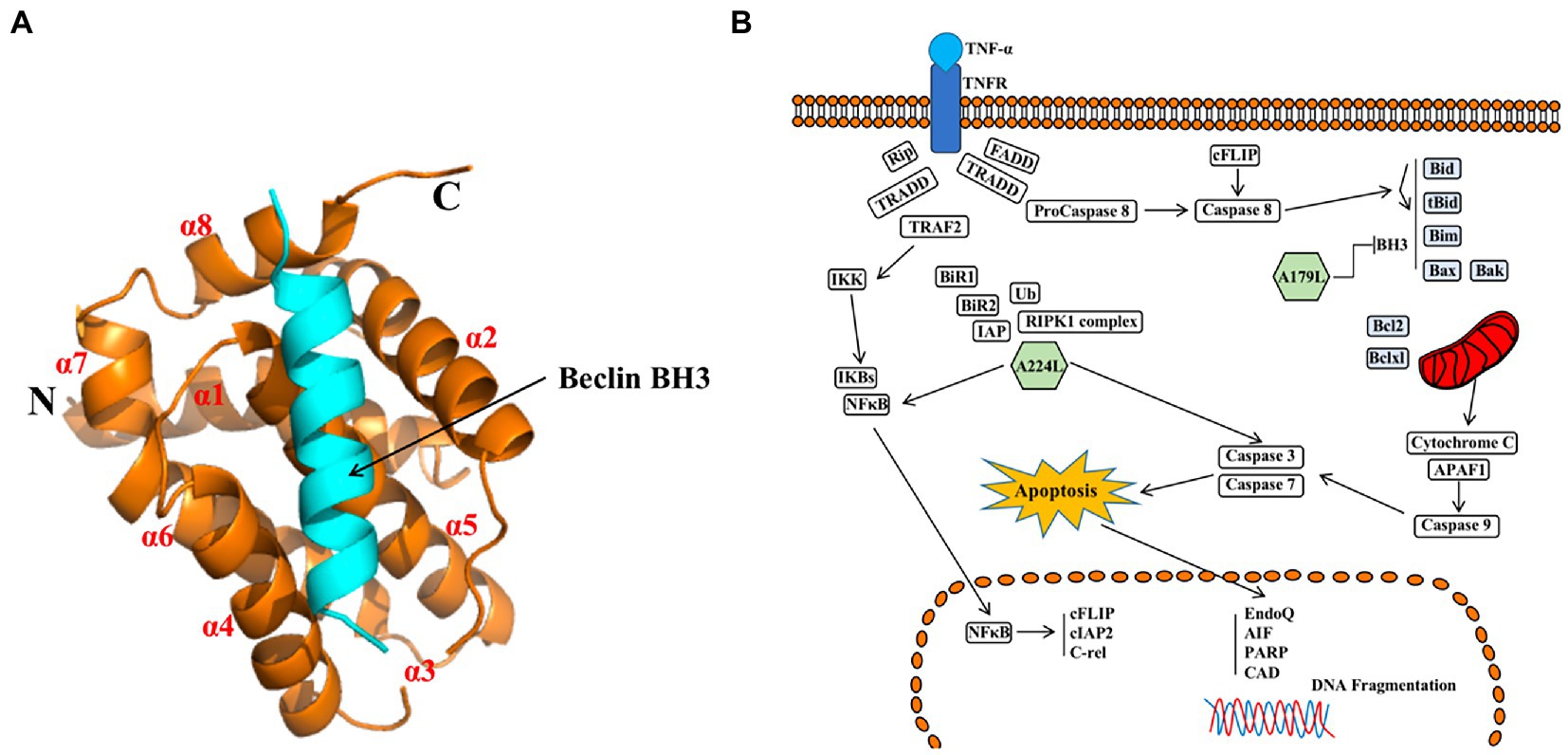
A179L protein, a potent inhibitor of apoptosis, is a member of the Bcl-2 family (Brun et al., 1996; Galindo et al., 2008), which is localized in mitochondria and the endoplasmic reticulum (Banjara et al., 2017). A179L protein adopts a Bcl-2 fold featuring eight α-helices arranged in a globular helical bundle fold. The canonical ligand-binding groove is formed by α-helices 2–5 and engages the BH3 motif of the pro-apoptotic protein Bcl-2 (Kvansakul and Hinds, 2013; Banjara et al., 2017, 2019; Figure 7A). Ionic interactions formed by binding of the E76 residue of A179L protein to the Bid R81 ligand, the D80 residue of A179L protein to the Bax K64 ligand, and the K79 residue of A179L protein to the Bax E61 ligand demonstrate the specificity of A179L protein (Banjara et al., 2017). In addition, A179L protein inhibits apoptosis signaling by binding the porcine pro-apoptotic Bcl-2 proteins Bax, Bak, Bim, Bid, Bad, Bik, Bmf, Hrk, Noxa, and Puma (Banjara et al., 2019). A179L protein also binds full length the autophagy regulator Beclin as well as its BH3 motif, thus demonstrating the ability of A179L protein to interfere with both host apoptosis and autophagy signaling (Revilla et al., 1997; Banjara et al., 2017, 2019).
Figure 7. (A) Schematic diagram of ASFV A179L and BH3 complex structures (Banjara et al., 2019). (B) A224L and A179L of ASFV inhibit apoptosis of infected cells (Dixon et al., 2017). ASFV A224L and A179L are shown as green hexagons. ASFV A179L Bcl-2 family proteins bind to and inhibit several pro-apoptotic proteins with a BH3 motif. A224L IAP family proteins bind to and inhibit caspase 3 and activate the nuclear factor Kappa-light chain enhancer of activated B cell (NF-κB) signaling, thereby increasing the expression of anti-apoptotic genes including cFLIP, cIAP2, and C-rel (Dixon et al., 2017).
In conclusion, A224L and A179L proteins suppress ASFV infection-induced apoptosis. By inhibiting apoptosis, the virus avoids being exposed to the immune system by host cells, thus facilitating immune escape and viral proliferation (Wang Z. et al., 2022). In addition, studies of A224L and A179L proteins have demonstrated that apoptosis and the survival of ASFV-infected host cells are tightly regulated processes (Figure 7B). Apoptotic pathways in host cells proceed primarily through the extrinsic apoptotic pathway (EAP), also known as the death receptor pathway. The alternative pathway is the intrinsic apoptotic pathway, also known as the mitochondrial apoptotic pathway. Further, analysis of the structure, function, and mechanism of action for these proteins may provide for an understanding of viral proliferation and evasion of immune responses (Dixon et al., 2017).
3.6. Proteins involved in ASFV assemblyASFV contains an envelope, capsid, inner capsule membrane, core shell, and inner core (Wang et al., 2019; Andrés et al., 2020). These five components are assembled into a whole virion. In the assembly process, the components are connected and supported by each other, with structural proteins playing a key role. Some non-structural proteins interact with structural proteins to promote protein structure formation and as such are equally important.
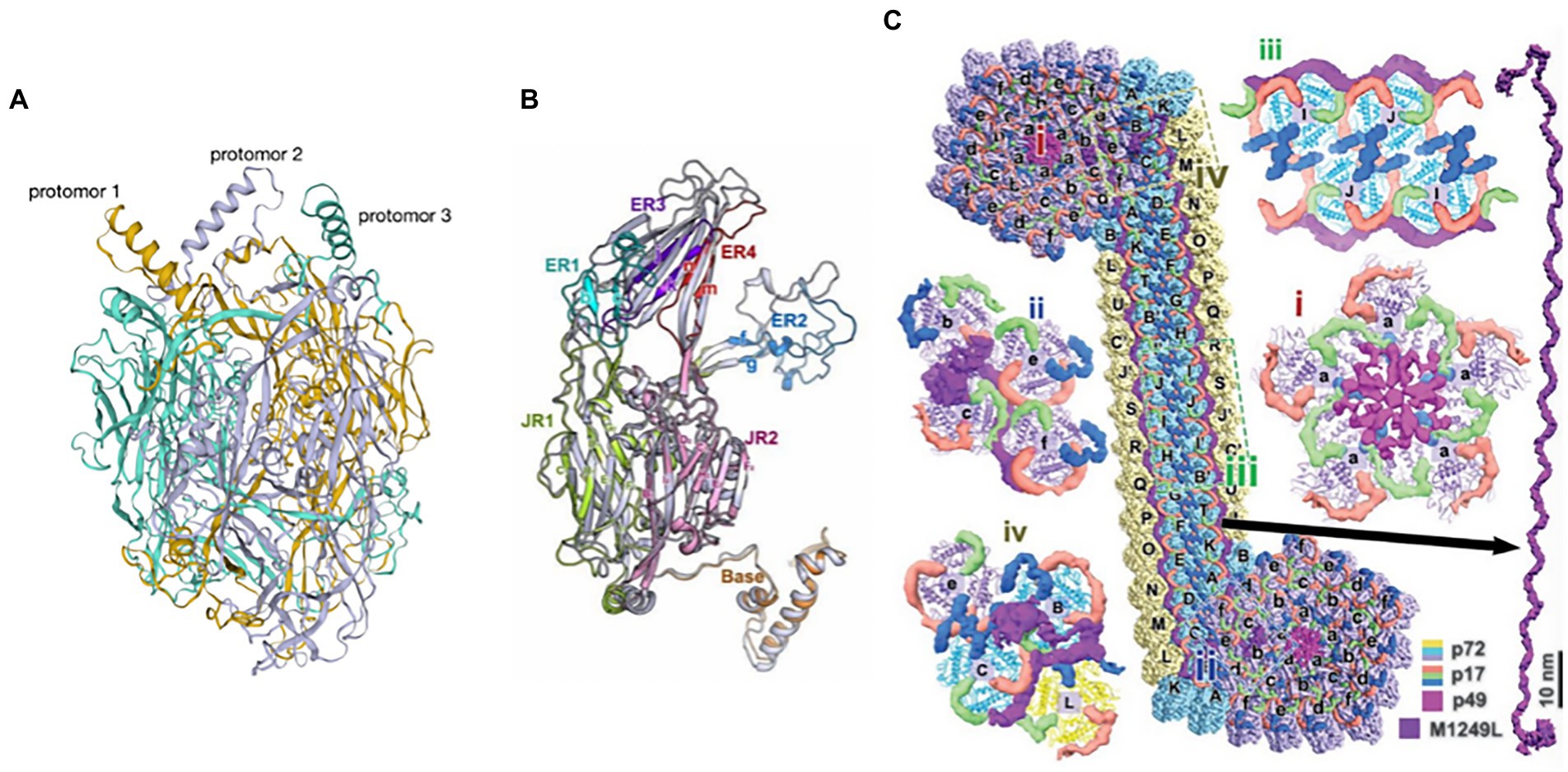
3.6.1. p72 and its molecular chaperone pB602Lp72 is one of the key protective antigens recognized by the immune system in response to ASFV and also the ASFV major capsid protein (Borca et al., 1994; Escribano et al., 2013). The ASFV capsid is constructed of pseudo-hexameric capsomers and pentameric capsomers. Each pseudo-hexameric capsomer is composed of three p72 molecules, and each pentameric capsomer is composed of five penton proteins (H240R). p72 forms a homotrimer (Figure 8A), with each monomer (Figure 8B) adopting a double jelly-roll structure that makes up pseudo-hexameric capsomers (p72 capsomers; Wang et al., 2019; Meng et al., 2022). ASFV capsid assembly is a gradual process involving p72. First, the penton complex docks with the inner membrane, where it recruits p72 capsomers, which form the penton core and initiate assembly. The skeleton unit M1249L then attaches to the penton core, and p72 capsomers and p17 contribute to the formation of zippers. The zippers connect neighboring penton cores and gradually construct a polyhedral framework. Finally, accompanying the formation of the polyhedral framework, p72 capsomers fill in the trisymmetrons and complete the capsid assembly (Wang et al., 2019; Meng et al., 2022). p72 may have potential neutralizing epitopes. Exposed region 1 (ER1) and ER2 form the crown of the p72 capsomer orienting toward the outside of the capsid, which may contribute to a conformational epitope. The β strands of ER3 and ER4 constitute a four-stranded β-sheet that shapes the head of the p72 capsomer and may be another conformational epitope. ER3 may link these two conformational epitopes, and these four ERs probably define the neutralizing epitopes (Andrés et al., 2020).
Figure 8. (A) The trimeric structure of the p72 protein (Wang et al., 2019). Deep yellow: protomer 1; lavender: protomer 2; cyan: protomer 3. (B) Diagram of the p72 monomer. The domains base, JR1 (jelly roll 1), JR2 (jelly roll 2), and ER1 to ER4 are shown (Wang et al., 2019). (C) Diagram of the interaction of p72 with H240R, M1249L, p17, and p49 (Wang et al., 2019).
p72 also has a molecular chaperone, pB602L, which promotes correct folding of p72. The viral assembly process is severely altered in the absence of pB602L, with the generation of aberrant “zipper-like” structures instead of icosahedral virus particles. Further, repression of pB602L synthesis affects the proteolytic processing of polyproteins, pp220 and pp62, and reduces the expression level of p72 (Epifano et al., 2006). However, the mechanism through which pB602L affects p72 is unclear, and this interaction may be direct. Understanding the structure and function of pB602L may provide valuable insight into the structure of p72.
3.6.2. Interaction of p72 with H240R, M1249L, p17, and p49H240R is a capsid protein of ASFV that is a penton protein found in the cytoplasm. H240R as an uncharacterized but essential virion protein that exhibits a single jelly roll and a globular cap that is different from, yet homologous to p72 (Wang et al., 2019; Andrés et al., 2020). H240R is encapsulated by a p72 shell that fills the pentameric capsomer and forms an apex to facilitate the assembly of the entire capsid. However, H240R is not involved in viral attachment or entry into pulmonary alveolar macrophages (PAMs) (Zhou et al., 2022). H240R, p17, p49, and M1249L form a complicated network immediately below the outer capsid shell, stabilizing the whole capsid (Figure 8C; Wang et al., 2019; Andrés et al., 2020).
M1249L is a skeleton protein with multiple helices and a fiber-like configuration with two terminal lobes, with the capacity to fix one pentasymmetron and link two neighboring pentasymmetrons. M1249L extensively interacts with p17 and p72 capsomers to form a rigid zipper structure, facilitating the formation of capsids (Wang et al., 2019).
p17 is encoded by the D117L gene and is a major structural transmembrane protein localized in the capsid and inner lipid envelope (Muñoz and Tabarés, 2022; Zheng et al., 2022). p17 is an essential and highly abundant protein required for the assembly of capsids and icosahedral morphogenesis (Xia et al., 2020). p17 is closely associated with the base domain of p72, with three copies of p17 encircling each p72 capsomer within the inner capsid shell, which firmly anchors p72 capsomers on the inner membrane, ensuring the stabilization of capsids (Andrés et al., 2020). Recently, p17 was shown to inhibit cell proliferation through ER stress and ROS-mediated cell cycle arrest, which may implicate p17 in ASF pathogenesis. p17 may also inhibit the cGAS-STING pathway through its interaction with STING and interference in the recruitment of TBK1 and IKKϵ, which would inhibit the IFN response and would promote immune evasion (Xia et al., 2020; Zheng et al., 2022).
p49 is encoded by the ASFV B438L gene, forms the outer capsid with p72, is located in close proximity to the capsid vertices, and is required for the formation of the capsid vertices (Alejo et al., 2018). P49 associates with the membrane where it mediates the docking of the penton complex to the inner membrane, recruiting capsomers to form the penton core, which initiates assembly (Wang et al., 2019). p49 is not involved in particle transportation from the virus assembly site to the plasma membrane (Epifano et al., 2006).
In summary, p72 capsomers are insufficient to assemble higher-order structures through only the three known and distinct assembly patterns: “head to back,” “head to head,” and “back to back” (Wang et al., 2019). H240R, M1249L, p17, and p49 are needed to build the networks that trigger the assembly of the whole capsid. But as the major capsid protein, p72 constitutes most of the outer capsid and is one of the key immune protective antigens of ASFV. The analysis of p72 structure will provide a new reference for the design of antigenic epitope-based vaccines in the future.
4. DiscussionASFV has a complex structure comprised numerous proteins. The function and mechanism of action for these proteins are not well described, which has made difficult the development of an efficient and effective ASF vaccine. Thus, it is necessary to investigate the structure and function of ASFV proteins, as a means by which to understand the infectious mechanisms of the virus. However, significant progress has been made in understanding the structure and function of ASFV proteins. For example, the crystal structure of the major capsid protein, p72, has been solved, thus demonstrating the structure of its homologous trimer and the interaction with H240R, M1249L, p17, and p49 proteins, providing a deeper understanding of the assembly of ASFV in host cells after infection. ASFV has a complex structure, with not only a capsid, but also a capsule, inner membrane, core shell, and genome. Proteins within these structures have specific functions, such as A179L and A224L that inhibit the apoptosis of host cells, and g5Rp that promotes the translation of virus mRNA within host cells. pS273R catalyzes the maturation of pp220 and pp62 multiprotein precursors into the core shell proteins p5, p34, p14, p37, p150, and p15, p35, and p8 that are involved in ASFV. Vaccines based on structural design can improve the antigenicity and immune protective qualities of vaccines (Nuñez Castrejon et al., 2022; Thomas and Abraham, 2022), but ASFV proteins are numerous, with more than 150–200 proteins produced during infection. At present, only a small number of ASFV protein structures are known and the function of most of these proteins is still unknown. Resolution of the structure and function of ASFV proteins will provide a clearer understanding of the interaction between virus and host, and the mechanism of ASFV replication and transmission. Further, this understanding will allow for convenient screening for protective antigens, analysis of neutralization epitopes, and the design of more targeted and effective ASF vaccines.
Author contributionsSY and WL conceived and designed the study. SY wrote the first draft of the manuscript. SY, WL, CM, and GZ revised the manuscript. SY and CM made the diagram. JS and HC performed the funding acquisition. All authors contributed to the article and approved the submitted version.
FundingThis work was financially supported by the National Key Research and Development Program of China (2021YFD11801402).
AcknowledgmentsWe thank the members of our laboratory for their constructive comments.
Conflict of interestThe authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s noteAll claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
ReferencesAlonso, C., Miskin, J., Hernáez, B., Fernandez-Zapatero, P., Soto, L., Cantó, C., et al. (2001). African swine fever virus protein p54 interacts with the microtubular motor complex through direct binding to light-chain dynein. J. Virol. 75, 9819–9827. doi: 10.1128/JVI.75.20.9819-9827.2001
PubMed Abstract | CrossRef Full Text | Google Scholar
Andrés, G.́., Alejo, A.́., Salas, J.́., and Salas, M.́. L. (2002a). African swine fever virus polyproteins pp220 and pp62 assemble into the core shell. J. Virol. 76, 12473–12482. doi: 10.1128/JVI.76.24.12473-12482.2002
PubMed Abstract | CrossRef Full Text | Google Scholar
Andrés, G., Charro, D., Matamoros, T., Dillard, R. S., and Abrescia, N. G. A. (2020). The cryo-EM structure of African swine fever virus unravels a unique architecture comprising two icosahedral protein capsids and two lipoprotein membranes. J. Biol. Chem. 295, 1–12. doi: 10.1074/jbc.AC119.011196
PubMed Abstract | CrossRef Full Text | Google Scholar
Andrés, G.́., García-Escudero, R.́., Salas, M.́. L., and Rodríguez, J. M. (2002b). Repression of African swine fever virus polyprotein pp220-encoding gene leads to the assembly of icosahedral core-less particles. J. Virol. 76, 2654–2666. doi: 10.1128/JVI.76.6.2654-2666.2002
PubMed Abstract | CrossRef Full Text | Google Scholar
Báez-Santos, Y. M., St John, S. E., and Mesecar, A. D. (2015). The SARS-coronavirus papain-like protease: structure, function and inhibition by designed antiviral compounds. Antiviral Res. 115, 21–38. doi: 10.1016/j.antiviral.2014.12.015
PubMed Abstract | CrossRef Full Text | Google Scholar
Banjara, S., Caria, S., Dixon, L. K., Hinds, M. G., and Kvansakul, M. (2017). Structural insight into African swine fever virus A179L-mediated inhibition of apoptosis. J. Virol. 91:e02228. doi: 10.1128/JVI.02228-16
PubMed Abstract | CrossRef Full Text | Google Scholar
Banjara, S., Shimmon, G. L., Dixon, L. K., Netherton, C. L., Hinds, M. G., and Kvansakul, M. (2019). Crystal structure of African swine fever virus A179L with the autophagy regulator Beclin. Viruses 11:789. doi: 10.3390/v11090789
PubMed Abstract | CrossRef Full Text | Google Scholar
Barber, C., Netherton, C., Goatley, L., Moon, A., Goodbourn, S., and Dixon, L. (2017). Identification of residues within the African swine fever virus DP71L protein required for dephosphorylation of translation initiation factor eIF2α and inhibiting activation of pro-apoptotic CHOP. Virology 504, 107–113. doi: 10.1016/j.virol.2017.02.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Barrado-Gil, L., del Puerto, A., Muñoz-Moreno, R., Galindo, I., Cuesta-Geijo, M. Á., Urquiza, J., et al. (2020). African swine fever virus ubiquitin-conjugating enzyme interacts with host translation machinery to regulate the host protein synthesis. Front. Microbiol. 11:622907. doi: 10.3389/fmicb.2020.622907
PubMed Abstract | CrossRef Full Text | Google Scholar
Borca, M. V., Irusta, P., Carrillo, C., Afonso, C. L., Burrage, T., and Rock, D. L. (1994). African swine fever virus structural protein p72 contains a conformational neutralizing epitope. Virology 201, 413–418. doi: 10.1006/viro.1994.1311
PubMed Abstract | CrossRef Full Text | Google Scholar
Brun, A., Rivas, C., Esteban, M., Escribano, J. M., and Alonso, C. (1996). African swine fever virus gene A179L, a viral homologue of bcl-2, protects cells from programmed cell death. Virology 225, 227–230. doi: 10.1006/viro.1996.0592
PubMed Abstract | CrossRef Full Text | Google Scholar
Cackett, G., Matelska, D., Sýkora, M., Portugal, R., Malecki, M., Bähler, J., et al. (2020). The African swine fever virus Transcriptome. J. Virol. 94:e00119–20. doi: 10.1128/JVI.00119-20
留言 (0)