





記住我
The protozoan Leishmania infantum is the etiological agent of visceral leishmaniasis (VL), one of the most important parasitic diseases worldwide. Brazil ranks the top five countries with the highest number of human cases and the highest fatality rate (PAHO, 2020). Despite many control measures for VL in Brazil, the incidence of human cases is still rising, especially in urban areas. Indeed, the country shows an average of 3.404 new cases/100,000 people per year, and 23 out of 27 states notified human VL cases (MS-BR, 2019; MS-BR, 2021), indicating a widespread occurrence of this disease.
The adaptive plasticity of Lutzomyia longipalpis complex (the invertebrate vector of L. infantum) to urban environments (Salomón et al., 2015), and the presence of other host species than human and domestic dogs (the main vertebrate reservoir) in urban VL endemic areas contribute to the complexity of the enzootic scenarios. There is strong evidence that synanthropic species as rodents, bats and marsupials play a role in the maintenance of L. infantum in urban and peri-urban endemic areas in Brazil (Oliveira et al., 2005; Carreira et al., 2012; Lima et al., 2013; de Castro Ferreira et al., 2015; de Rezende et al., 2017; Castro et al., 2020; Vieira et al., 2022).
Wild carnivore species have been reported parasitized by L. infantum in Brazil since the half of the 20th century, when the Deane’s couple described the infection in the crab-eating fox (Cerdocyon thous) (Deane and Deane, 1955), although the host was inaccurately reported as hoary fox (Lycalopex vetulus) (Courtenay et al., 1996). Indeed, crab-eating fox can be considered a putative L. infantum reservoir host (Roque and Jansen, 2014), as attested by parasitological assays in different studies (Lainson et al., 1990; Silva et al., 2000; Courtenay et al., 2002; Quinnell and Courtenay, 2009). Nevetheless, the free-living crab-eating fox infected by L. infantum seems to be less competent to infect vectors than dogs (Courtenay et al., 2002). Apart from crab-eating fox, the role of other wild Carnivora in the epidemiology of VL is still unknown. It has been recorded the presence of L. infantum in intact skin of a captive bush dog (Speothos venaticus) (Figueiredo et al., 2008), although the potential of its transmissibility for the vector was reported to be low (Mol et al., 2015). The transmissibility shown for captive maned wolves (Chrysocyon brachyurus) was also very low (Mol et al., 2015). As these species do not live near human habitations, they are considered to have a secondary importance in epidemiology of VL in the urban areas (Dantas-Torres and Brandão-Filho, 2006).
The South American coati (Procyonidae: Nasua nasua; hereafter “coati”) is a carnivore species well adapted to urban forest fragments in some regions of Brazil (Estevam et al., 2020; Barreto et al., 2021). To date, the only association between L. infantum and this taxon was reported by Paiz et al. (2015), who showed serological evidence of infection. Additionally, some reports have detected Leishmania spp. in this carnivore species through serological and molecular methods (Voltarelli et al., 2009; Porfirio et al., 2018; Reis et al., 2020), and L. shawi was isolated from this mammal (Lainson et al., 1989).
Campo Grande (CG), the capital of Mato Grosso do Sul (MS) state, Midwest Brazil, is considered an endemic area for human and canine VL (Botelho and Natal, 2009; Furlan, 2010; Brazuna et al., 2012; de Sousa et al., 2013; SES/MS, 2022). In this region, L. infantum has been detected in domestic cats (Antunes et al., 2016; Metzdorf et al., 2017) and wild mammal species as opossum and bats (Humberg et al., 2012; de Rezende et al., 2017; Castro et al., 2020). Moreover, L. longipalpis is present in this region (Cunha et al., 2014), indicating a high-risk area for L. infantum transmission (Falcão de Oliveira et al., 2020; SES/MS, 2022). Since, Barreto et al. (2021) reported that coati is a conspicuous species in the urban forest fragments at CG, we aimed to investigate the L. infantum infection in this mammal population. Our hypothesis is that this wild mammal species is affected by the local epidemiological scenario of L. infantum transmission.
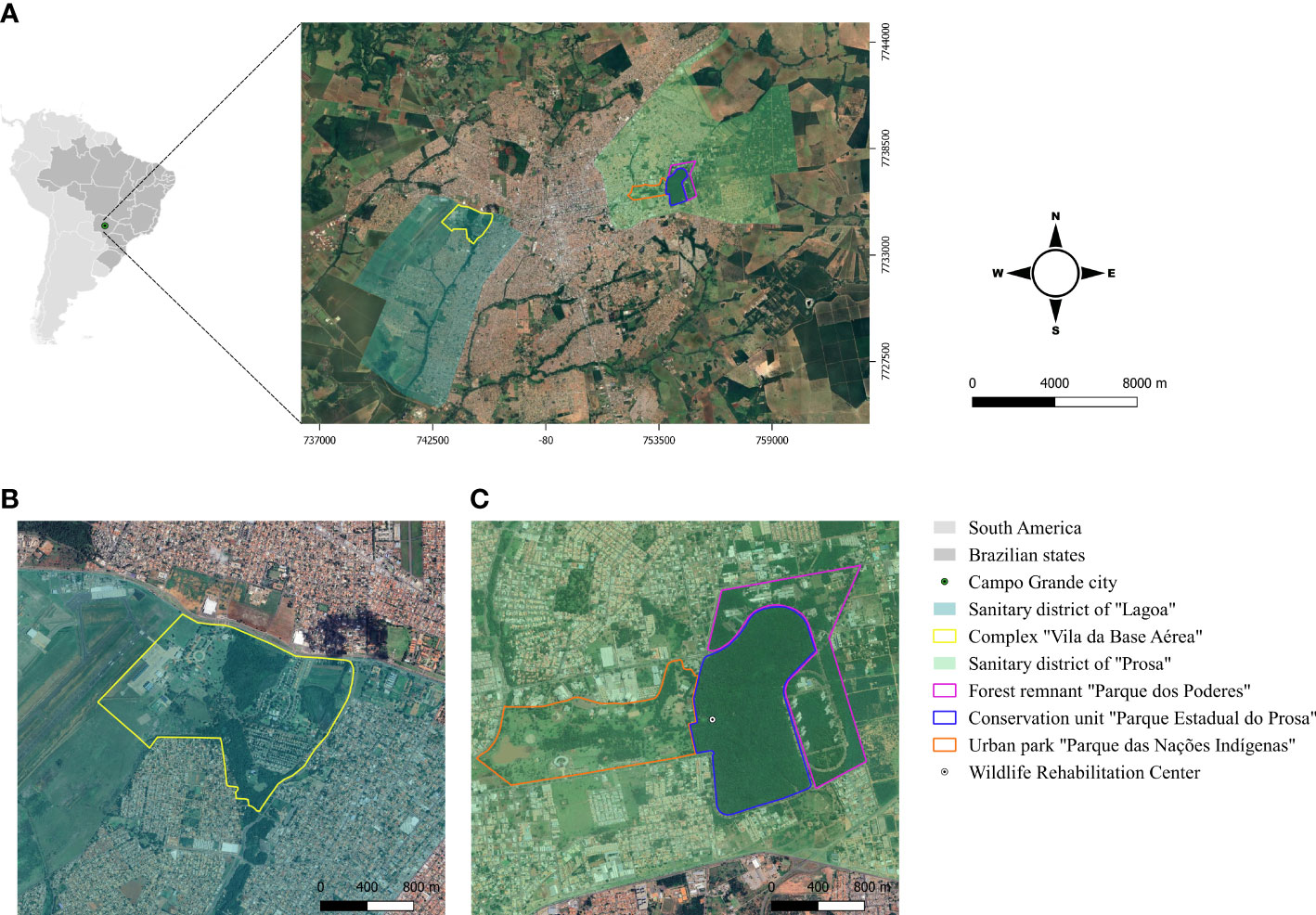
2 Material and methods2.1 Study areas and field proceduresThe study was conducted in two forest fragments “Parque Estadual do Prosa” (PEP) (20°26’59’’S, 54°33’55’’O) and “Vila da Base Aérea” (VBA) (20°28’17’’S, 54°39’14’’O), located in the urban area of CG/MS (Figure 1A). The population densities of coatis in these areas were estimated as 11.2 individuals/km2 in PEP and 19.5 individuals/km2 in VBA (Barreto et al., 2021). The fieldwork occurred every two months from March 2018 to May 2019 encompassing two weeks in each area.
Figure 1 (A) Geographical location of the study areas in Campo Grande, Mato Grosso do Sul, Midwest Brazil; (B) Conservation unit “Parque Estadual do Prosa” and its adjacent areas “Parque dos Poderes” and “Parque das Nações Indígenas”; (C) Complex “Vila da Base Aérea”.
The PEP is a conservation unit of 135 hectares composed of cerradão (tall savanna woodland) and riparian forest, and is located in the sanitary district of “Prosa” (Figure 1B). Human access is restricted in the PEP, but there is daily flow of people in the adjacent areas, the urban park “Parque das Nações Indígenas” and the public administration area “Parque dos Poderes” located in a forest remnant of Cerrado. Coatis have free access in all the three areas either by crossing the paved streets or passing through the branches of the trees near the borders (Figures 2A, B). In addition, the PEP presents a diverse mammalian fauna and shelters a Wildlife Rehabilitation Center (CRAS) (https://www.imasul.ms.gov.br/centro-de-reabilitacao-de-animais-silvestres-cras/) that receives and keeps several wild animals from all over the MS state.
Figure 2 South American coatis (Nasua nasua) in the conservation unit “Parque Estadual do Prosa” (PEP) and in the residential area “Vila da Base Aérea” (VBA) at Campo Grande, Mato Grosso do Sul, Midwest Brazil. (A) South American coatis crossing paved streets from “Parque dos Poderes” to PEP; (B) South American coatis using the arboreal strata to move from PEP to Parque das Nações Indígenas; (C) South American coatis searching for food in outdoor trash cans in VBA; (D) South American coatis foraging near the not fenced houses in VBA.
The VBA, located in the sanitary district of “Lagoa”, is a complex of 197 hectares divided into a military operational area and a residential area surrounded by a dense vegetation that includes riparian forest and veredas (a plant formation with hydromorphic soil located in the vicinity of river springs) (Figure 1C). The residential portion is inhabited by at least 730 humans who raise domestic animals such as dogs, cats and chickens. The houses are not fenced and have many trash cans in front, which serve as food source for the coatis (Figures 2C, D). During fieldwork, few individuals of other wild mammal species were seen in the area, as the nine-banded armadillo (Dasypus novemcinctus), Azara’s agouti (Dasyprocta azarae), Brazilian guinea pig (Cavia aperea) and howler-monkey (Alouatta caraya), and other species such as the coati and Capybara (Hydrochoerus hydrochaeris) are abundant.
The coatis were captured and randomly recaptured in box traps (90 × 45 × 50; Equipos Fauna®, Brazil) baited with 15 to 25 grams of bacon. All the captured coatis were sedated with an association of tiletamine hydrochloride and zolazepam hydrochloride (Telazol 100g; Zoetis®, USA), tagged with subcutaneous transponders (Animal Tag®, Brazil) and numbered colored ear-tags (Qualyplast® Brazil). Blood samples were collected through femoral venipuncture using 0.25 x 0.8 mm needles (BD Vacutainer®) and deposited in clot activator tubes to obtain the serum. Bone marrow (BM) samples were collected from manubrium sterni of animals > 2.5 kg using 0.40 x 1.2 mm hypodermic needles and 10 mL syringes, and then placed in tubes containing Ethylenediamine Tetraacetic Acid (EDTA). These procedures were performed after appropriate asepsis of the collection site using bactericide soap, iodized ethanol and ethanol 70%. Ear skin fragments (0.4 to 30 mg) were obtained with sterilized material and stored in 99% ethanol (Sigma-Aldrich®, United States). After total recovery from anesthesia, the animals were released at the capture site.
The field procedures were conducted in accordance with the Instituto Chico Mendes de Conservação da Biodiversidade (license number 56912-2), the Instituto de Meio Ambiente de Mato Grosso do Sul (license number 05/2017, process No.61/405959/2016), and Air force cooperation agreement (No.01/GAP-CG/2018). The present study was approved by the Ethics Committee for Animal Use of Universidade Católica Dom Bosco, Campo Grande, MS (license number 001/2017).
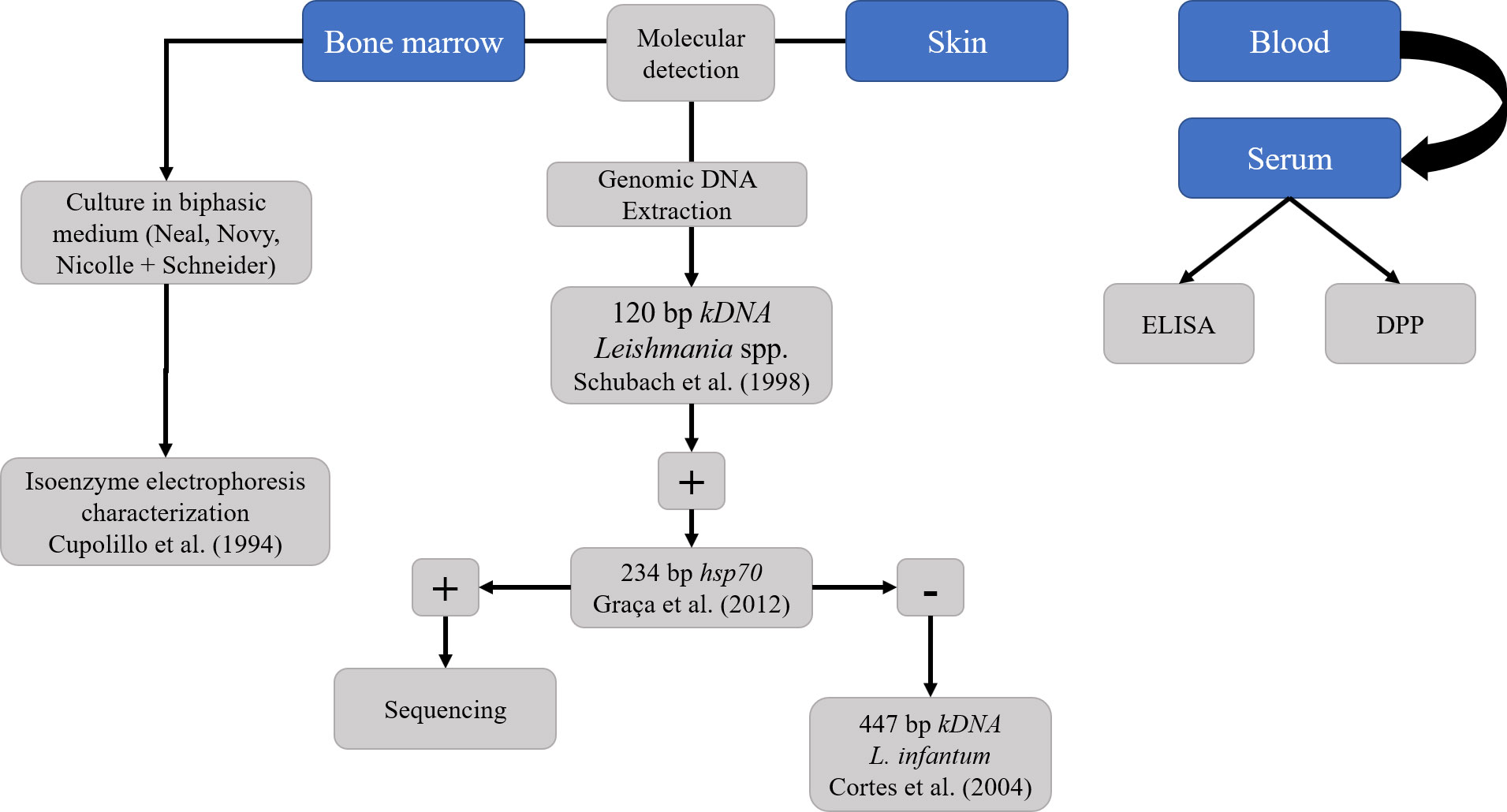
2.2 Serological detection of Leishmania infantumA serological survey for the detection of anti-L. infantum IgG was performed through the Enzyme-Linked Immunosorbent Assay (ELISA) (modified from EIE®, BioManguinhos, Rio de Janeiro, Brazil) (Alves et al., 2016) (Figure 3). The sera of the coatis were tested with anti-Raccoon IgG HRP conjugated (Bethyl Laboratories®, Inc., Montgomery, Texas, United States), diluted at 1:70,000, and each microtiter polystyrene plate contained positive and negative control samples in duplicates. The cut-off point was established by the mean Optical Density (OD) of the negative control ± three standard deviations and the gray range adopted was 20% above the cut-off value. Serum samples were also submitted to a Dual Path Platform chromatographic immunoassay (DPP® CVL BioManguinhos, Rio de Janeiro, Brazil) for rapid detection of K26/K39 specific antibodies against L. infantum (da Costa et al., 2003; Rodrigues et al., 2022). To check the reliability of rapid test, the serum samples were tested for Protein A affinity, showing high affinity, as similar as the dog serum samples (data not shown). To improve the accuracy of the serological detection of L. infantum, the serum samples of coatis were also screened for Trypanosoma cruzi infection using ELISA according to Alves et al. (2016), and the Chagas/Bio-Manguinhos Lateral Flow Immunochromatographic Rapid Test (Chagas-LFRT), as described by Rodrigues et al. (2022). All the tests were performed according to the manufacturer’s recommendations, and seropositivity was considered only when the samples presented positivity in both anti-L. infantum IgG ELISA and DPP® CVL.
Figure 3 Diagnostic strategy for detection of Leishmania infantum in South American coatis (Nasua nasua) from forest fragments of Campo Grande, Mato Grosso do Sul, Midwest Brazil.
2.3 Parasitological and molecular detection of Leishmania infantumThe axenic cultures were performed by placing 200 µL of BM aspirates in duplicates in biphasic media, Novy-MacNeal-Nicolle (NNN) with Schneider’s insect medium overlay supplemented with 10% fetal bovine serum. The tubes were incubated at 26°C – 28°C and weekly examined for 30 days. After the exponential growth phase of the flagellates, the positive culture was cryopreserved at – 196°C under liquid nitrogen. The promastigote forms of the isolated culture were deposited in the Collection of Leishmania (CLIOC) (clioc.fiocruz.br) of Instituto Oswaldo Cruz and characterized by them using isoenzyme electrophoresis of G6PDH and 6PGDH (Cupolillo et al., 1994).
Genomic DNA was extracted from 200 µL of the BM samples through the phenol-chloroform method according to Sambrook and Russell (2001), and from the skin fragments using the DNeasy® Blood and Tissue Kit (Qiagen®, Netherlands). The DNA was eluted in 50 µL elution buffer and stored at -20°C until molecular analysis. The concentration and quality were assessed through spectrophotometry (Biodrop, Analítica®, Brazil).
The BM and skin samples were firstly screened through Polymerase Chain Reaction (PCR) using a pair of primers (L1/L2) directed to the kinetoplastid DNA (kDNA) minicircle molecules of Leishmania spp. according to Schubach et al. (1998). Positive samples were then submitted to PCR targeting the heat-shock protein 70 gene (HSP70) and sequenced, as described by Graça et al. (2012) (Figure 3). Sanger sequencing was performed in the AB3500 platform (Applied Byosystems®, United States) at ACTGene Molecular Analysis biotechnology company (https://actgene.com.br/). Negative samples at HSP70 PCR were submitted to L. infantum-specific kDNA PCR (MC1/MC2) according to Cortes et al. (2004) (Figure 3). All the PCRs were conducted in a thermocycler (GeneAmp PCR System 9600, Applied Byosystems®, United States), using positive controls of L. infantum (LHV14 strain) and L. amazonensis (IFLA/BR/1967/PH8 strain) and negative controls (ultrapure water). The PCR products were visualized on 1.5% agarose (Kasvi®, Brazil), stained with Gel Red Nucleic Acid Stain (Biotium®, EUA).
2.4 Statistical analysisIn order to observe the agreement between serological assays (ELISA and DPP® CVL), the Cohen’s Kappa test was performed with a 95% confidence interval and interpreted as follows: poor agreement (k = 0), slight agreement (0.20 ≤ k ≥ 0), fair agreement (0.40 ≤ k ≥ 0.21), moderate agreement (0.6 ≤ k ≥ 0:41), substantial agreement (0.80 ≤ k ≥ 0.61), and almost perfect agreement (1.0 ≤ k ≥ 0.81). To check a possible influence of the studied areas and sex on L. infantum infection, a Chi-squared test (p < 0.05) was applied considering the individuals. We considered infection by L. infantum as positivity in any of the parasitological or molecular tests and/or positivity in both serological assays. Both Cohen’s Kappa and Chi-squared tests were performed through the R software (R Development Core Team, 2015).
3 Results3.1 Captures, recaptures and acquisition of samplesDuring the study, a total of 110 coatis were captured in the VBA and PEP. Blood was collected in all capture occasions corresponding to 21 males and 30 females from PEP, and 25 males and 34 females from VBA. The BM samples were collected from 85 individuals corresponding to 16 males and 26 females from PEP, and 19 males and 24 females from VBA. Skin samples were collected from 65 ear-tagged animals corresponding to 10 males and 20 females from PEP, and 14 males and 21 females from VBA.
Throughout the study, we recaptured a total of 41 individuals (eight males and eight females from PEP, and 10 males and 15 females from VBA) in 80 occasions (Figure 4). Serum samples were collected, and serologic tests were performed in all recaptures, totalizing 190 samples (73 from PEP and 117 from VBA). Bone marrow samples were collected from 51 recaptures and submitted to molecular analysis (totalizing 136 samples = 63 from PEP and 73 from VBA). Skin samples were not collected during recaptures.
Figure 4 Recapture events of South American coatis (Nasua nasua) from the conservation unit “Parque Estadual do Prosa” (PEP) and the residential area “Vila da Base Aérea” (VBA) from March 2018 to April 2019.
3.2 Serological detection of Leishmania infantumA total of 65 samples from 33 individuals were seropositive (ELISA+/DPP+) to L. infantum, indicating an overall seroprevalence of 30% (33/110). In PEP, the seroprevalence rate was 11.8% (6/51), corresponding to two males and four females, while in VBA the seroprevalence was estimated in 45.8% (27/59), corresponding to 11 males and 16 females. Despite the discordance observed in 19 samples (16 ELISA+/DPP- and three ELISA-/DPP+), the Cohen’s kappa test attested a substantial agreement between ELISA and DPP (k = 0.78; CI 95% = 0.68 – 0.87; p = 0.001). Only one serum sample of a non-recaptured female coati from VBA, that was seropositive to L. infantum, presented seropositivity for T. cruzi, indicating a mixed infection.
3.3 Parasitological and molecular detection of Leishmania infantumDuring our study, we isolated L. infantum from one BM sample derived from a seropositive male captured in VBA (ID 11528) (Figure 5). This isolate was deposited in the CLIOC (access number: IOC-L 3780), but the molecular diagnosis was not performed in this individual due to insufficient amount of BM sample.
Figure 5 Parasitological and molecular detection of Leishmania infantum from the bone marrow samples of South American coatis (Nasua nasua) from urban forest fragments in Campo Grande, Mato Grosso do Sul, Midwest Brazil.
The screening kDNA PCR in BM samples showed an infection rate of 41.2% (35/85) for Leishmania sp. Posteriorly, L. infantum infection was confirmed in 16 individuals (nine by HSP70 sequencing and seven by MC1/MC2 PCR), seven from PEP (four males and three females) and nine from VBA (three males and six females). The results of parasitological and molecular assays performed in the coatis are presented in Figure 5. From the 19 individuals negative to L. infantum in molecular tests, four were classified as seropositive and 15 were seronegative. Only one skin sample was positive in kDNA PCR and L. infantum infection was confirmed by MC1/MC2 PCR. The same individual (ID 10826, a male from VBA) was seropositive and also positive in BM molecular assay (with L. infantum infection confirmed by HSP70 sequencing).
The BLASTn analysis of the nine HSP70 DNA sequences revealed identity of 100% with L. donovani HSP70 gene obtained from samples of humans from Ethiopia and China (accession numbers: FN669773.1 and JX970996.1 respectively). The obtained sequences were deposited in the GenBank database under the following accession numbers: ON805836 to ON805844 (Figure 5).
3.4 Data analysisTogether, the parasitological, serology and molecular tests showed a L. infantum infection rate of 36.4% (40/110). These 40 individuals correspond to 24 positive by serological tests (including an individual positive in parasitological test), seven positive by molecular tests, and nine positive by both tests. We observed through Chi-squared test that infection was significantly higher in VBA (49.1%, 29/59) than in PEP (21.6%, 11/51) (Chi2 = 90; df = 1; p = 0.003). Our analysis did not show influence of sex in L. infantum infection (Chi2 = 1.580; df = 1; p = 0.208).
When considering only individuals assessed by both serology and molecular tests (N = 85), we observed distinct L. infantum infection patterns (Table 1).
Table 1 South American coatis (Nasua nasua) infected by Leishmania infantum by both serological and molecular diagnosis at urban forest fragments in Campo Grande, Mato Grosso do Sul, Brazil.
3.5 Longitudinal surveyAmong the 40 coatis positive for L. infantum, 19 individuals were recaptured during the study at least on one occasion. Eleven individuals remained positive by at least one diagnostic test throughout their recaptures (Figure 6). Of these, seven were only seropositive (IDs 10433, 10435, 10439, 10442, 10508, 10999 and 11522) and four presented positivity also in molecular tests (IDs 10443, 10444, 10507 and 10509). Five individuals (IDs 10320, 10322, 10323, 10440 and 10831) presented positivity (four by MC1/MC2 PCR and one by serological test) either in the capture or in the first recapture, and negativity in the subsequent recapture (Figure 6). We observed three seroconversion events, with individuals (IDs, 10441, 10511 and 10513) presenting positivity only in the subsequent recaptures (Figure 6). The remaining twenty-one positive animals were not recaptured: 13 were seropositive, five were positive in both serology and molecular tests and three were positive only in molecular assays.
Figure 6 Longitudinal results of 19 South American coatis (Nasua nasua) infected by Leishmania infantum in urban forest fragments in Campo Grande, Mato Grosso do Sul, Midwest Brazil, from March 2018 to April 2019. ID: Identification number; PEP: conservation unit Parque Estadual do Prosa; VBA: residential rea Vila da Base Aérea. Positivity in molecular was obtained either by HSP70 sequencing or MC1/MC2 PCR. Seropositivity was considered when both ELISA and DPP® CVL tests were positive.
4 DiscussionIn this study we report serological, parasitological and molecular detection of L. infantum in coatis from forested fragments of an endemic region for VL, with an overall infection rate of 36.4% (40/110). Our results demonstrate that this wild carnivore species is somehow affected by the L. infantum epidemiological scenario at the studied area. Indeed, (i) the endemicity of VL in humans and dogs at CG (SES/MS, 2022), (ii) abundance of the main vector L. longipalpis s.l. (de Souza Fernandes et al., 2022), and (iii) the high density of coatis in forest fragments of CG (Barreto et al., 2021) may favor the L. infantum transmission to coatis. Future studies may indicate the genetic flow of L. infantum populations among the multiple hosts involved in this epidemiological scenario, which was not the aim of the present study. The evaluation of recaptured individuals was a particular aspect of the present study because it provided evidence of long-term exposure of L. infantum in the studied populations, including seroconversion events.
Our findings indicated that L. infantum infect coatis from both studied forest fragments in CG, but a higher rate of infection in VBA when compared to PEP was observed (p = 0.003), indicating a spatial influence. This finding may be related to the geographical localization of the studied areas, as well as differences in land use. For instance, VBA is a residential area with many dogs, the main domestic reservoir host of L. infantum, and chickens, which although refractory to Leishmania infection, are amplifiers and attractors of sandfly populations (Casanova et al., 2013). Even though maximal dispersion of L. longipalpis is reported to be 243 meters in urban areas (de Oliveira et al., 2013), Coatis at VBA often forage near houses looking for food in outdoor trash cans, which may favor their exposition to with L. longipalpis in this environment. As described by Barreto et al. (2021), VBA has a denser population of coatis than PEP, and in this area the animals are prevented from dispersing out due to the physical barrier and the lack of functional connectivity in the surroundings. In addition, VBA is in the sanitary district of “Lagoa”, a region that had the higher number of notified cases and cumulative incidence of human VL from 2002 to 2009 (Brazuna et al., 2012). This sanitary district also had the second highest number of canine VL from 2007 to 2009 in CG, and a large sandfly abundance (Brazuna et al., 2012; de Souza Fernandes et al., 2022).
Oppositely, the lower infection rate observed in PEP (21.6%) may be related to several factors including the absence or low abundance of competent reservoirs, a higher richness of mammal species than VBA, and the CRAS, that is located in this conserved area. Although the coatis have access to the adjacent areas Parque das Nações Indígenas and Parque dos Poderes, there are no resident dogs in these areas that could sustain high rates of L. infantum infection, as in VBA. Also, there are 17 mammal species listed in the PEP Management Plan, and this number can be higher if we consider the captive animals of the CRAS, while in VBA few species were sighted. Assuming that the PEP presents a higher diversity of wild mammal species in comparison to VBA, we can expect that the PEP has a higher number of hosts not competent to be source of infection for the vector, which could reduce the risk of L. infantum infection in coatis through dilution effect (Keesing and Ostfeld, 2021). In addition, the CRAS, located inside the PEP, may influence the distribution of L. longipalpis due to the constant presence of captive animals and large amount of organic material from feces and concentrate food that could attract the vector and avoid its dispersion.
The detection of L. infantum in an ear skin fragment of an individual that was also seropositive and presented L. infantum DNA in BM sample (ID 10826), indicates that L. infantum is capable to colonize intact skin of coatis. In view of the importance for L. infantum transmission, domestic dogs are the main reservoir species mainly because they can present high parasitic loads in the skin, favoring the infection of the vectors (Dantas-Torres, 2007). Wild carnivores, as maned wolf, bush dog and crab-eating fox have been reported presenting L. infantum DNA in skin (Figueiredo et al., 2008; Souza et al., 2010; Jusi et al., 2011), although the first two demonstrated low competence to transmit the parasite to the vectors. Since the maintenance of a multi-host parasite as L. infantum in a given environment can involve different mammal species with diverse epidemiological competences (Roque and Jansen, 2014), efforts toward evaluation of blood meal source of sand flies in the forest fragments of CG, as well as isolation and quantification of parasite load from skin of different mammal species must be considered. Our results put the coati together with maned wolf and bush dog as carnivore species that may harbor parasite in intact skin. Nevertheless, their role in L. infantum epidemiology remains to be uncovered.
Our data showed no significant differences related to sex in the infection rate of L. infantum infection. The sex-related response to L. infantum infection includes a complex interaction among host, vector, and environment (Lockard et al., 2019), and therefore the sex differences in the prevalence of VL may not have a pattern. While higher VL prevalence in males have been recorded in humans and dogs naturally and experimentally infected (Guerra-Silveira and Abad-Franch, 2013; Lockard et al., 2019), studies with long-term data have shown no differences in the seroprevalence of L. infantum infection between male and female stray dogs (Miró et al., 2007; Müller et al., 2022). Risueño et al. (2018) found only a marginally greater L. infantum prevalence in female compared to male free-living rabbits. Given the nature of our data, we could not discriminate whether the absence of sex difference in the infection rate of L. infantum was due to biological differences or sex-related exposure. However, we can presume that social behavior of coatis in the urban matrix can contribute to a similar rate of infection between male and female. Coatis form social groups consisting of adult females and their offspring, with adult males joining the group only in the reproductive period (Gompper and Decker, 1998). However, Barreto et al. (2021) observed that males are cohesive to the group regardless of the reproductive period.
Our study showed that many individuals presented distinct infection patterns concerning their results in the serological and molecular assays. This dynamic, as well events of new infections observed in the longitudinal studies are expected in populations in endemic areas because individuals are constantly being exposed to infected vectors (Herrera et al., 2004; Oliva et al., 2006). As observed, seven seronegative coatis displayed positivity in the molecular assays suggesting recent infections, a common phenomenon found in populations inhabiting endemic areas (Herrera et al., 2004; Oliva et al., 2006). In dogs, the humoral response may take around three to five months after L. infantum infection to produce sufficient immunoglobulins to be detected by serological tests (Paranhos-Silva et al., 2003). It is important to mention that in endemic areas for leishmaniasis, about one fifth of L. infantum infected dogs presents low or undetectable anti-Leishmania antibodies (Ashford et al., 1995). On the contrary, seropositive animals with negative molecular results are expected because the serological assays detect antibodies that are maintained in serum for longer periods, while the molecular assays are positive only when parasites are present in the specific amount of analyzed sample. We highlight the importance of combine different diagnostic tests in order to improve the detectability of infected individuals.
The molecular diagnostic strategy used here allowed us to detect a greater number of positive samples. In fact, as the sensitivity of the HSP70 gene target is low when compared to the kDNA PCR assay (Graça et al., 2012), and the use of MC1/MC2 (L. infantum kDNA), as it is more sensitive and highly specific for L. donovani s.l., improved the detectability of molecular diagnostics. Concerning the recaptures, molecular negativity observed in the first recaptures of the individuals 10320 and 10322, and in the second recapture of the individual 10323 (Figure 6) could be attributed to fluctuations in detectable DNA from the bone marrow. Transient PCR bone marrow positivity has been reported in dogs naturally and experimentally infected with L. infantum (Paranhos-Silva et al., 2003; Oliva et al., 2006). As discussed by these authors, this may also be attributed to limited sensitivity of specific molecular tests, in our case, MC1/MC2 and HSP70, especially in the first and second recaptures of the individuals 10320 and 10323 respectively, which were positive in the screening kDNA for Leishmania spp. A rare case of seroreversion was observed in one individual (10831) that was seropositive in the capture and seronegative in the three subsequent recaptures (Figure 6). Seroreversion was already reported in both canine and human VL (Ostyn et al., 2011; Maia et al., 2020), probably by decrease of antibody concentration over time, as observed in the ELISA OD from this individual (Capture: 0,168; First recapture: 0,149; Second recapture: 0,104; Third recapture: 0,075).
In contrast, we found evidence of active transmission of L. infantum in the area, evidenced by new infections occurring in coatis during the study period. Indeed, five individuals were seronegative in the capture or in a given recapture, and positive by serology or molecular diagnosis in subsequent recapture. Two individuals (IDs 10443 and 10509) remained positive in molecular test in consecutive captures, indicating that individuals may be capable of sustain BM parasitism for at least five months. At this point, the remaining question is whether these animals are capable of sustain high parasite loads in the skin and act as source of infection to sand flies.
Our results point to the occurrence of other Leishmania species besides L. infantum in the sampled coatis, since (i) 54.3% (19/35) of the infections detected by kDNA PCR were not confirmed by L. infantum-specific targets, and (ii) we found 16 samples ELISA positive and DPP negative that were also negative in the serological tests performed to detect other common trypanosomatids that infects wild mammals: T. cruzi. Geographical overlap among Leishmania species is reported (Talmi-Frank et al., 2010), and probably occurs in Brazil, which shelters the largest richness of Leishmania species among Latin American countries (Herrera et al., 2020). In Campo Grande municipality, Nantes et al. (2021) obtained DNA sequences with high similarity to L. amazonensis and L. guyanensis from blood samples of opossums, and Castro et al. (2020) reported L. braziliensis infection in the bats Lasiurus cinereus and Cynomops planirostris. In addition, the presence of other species of Leishmania than L. infantum circulating in coatis of Campo Grande is possible due to the occurrence of many phlebotomine species as Nyssomyia whitmani, Ny. antunesi and Bichromomyia flaviscutellata, that are involved in transmission of several species of Leishmania commonly reported in outbreaks of cutaneous leishmaniasis such as L. braziliensis, L. shawi, L. guyanensis, L. lindenbergi and L. amazonensis (Costa et al., 2007; Silva et al., 2008; Oliveira et al., 2012; Vásquez Trujillo et al., 2013; Carvalho et al., 2018; de Souza Fernandes et al., 2022).
5 ConclusionCoatis living in urban forested fragments at CG are affected by the epidemiological scenario of VL transmission that evolves dogs, humans and L. longipalpis vectors, as demonstrated by their overall infection rate of 36.4%. The importance of this finding is highlighted by: (i) the high population densities of this mammal host in different urban forest fragments, (ii) their proximity to domiciliated areas and contact with human and dogs, (iii) the ability to maintain viable parasites in bone marrow, as demonstrated by L. infantum isolation and (iv) the possibility to maintain parasites in no lesioned ear skin, as demonstrated by L. infantum DNA detection. Given these findings, we highlight the importance to investigate the pattern of L. infantum infection in coatis inhabiting urban forest fragments to define their role as L. infantum reservoirs in endemic areas for VL.
Data availability statementThe datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article.
Ethics statementThe animal study was reviewed and approved by Ethics Committee for Animal Use of Universidade Católica Dom Bosco, Campo Grande, MS (license number 001/2017).
Author contributionsGCdeM, WTGB, ALRR and HMH wrote the manuscript. GCdeM, WTGB, ACR and WOdeA collected biological samples. SCdasCX, FMA and ARdaS contributed in diagnostic tests execution. FMS performed statistical analysis. CEdeO, GEdeOP, SCdasCX, FMA, GBdeA, AMJ, ALRR and HMH critically reviewed the manuscript. All authors contributed to the article and approved the submitted version.
FundingThis research was supported by the Foundation for Support to the Development of Education, Science and Technology of the State of Mato Grosso do Sul (FUNDECT) (Process 59/300.187/2016), Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ) (Process E-26/202.994/2019 and E-26/010.002276/2019), the National Council for Scientific and Technological Development (CNPq) to HMH (Process 308768/2017-5), AMJ (PQ1-A) and ALRR (CNPq Process 425293/2018-1), and scholarship to FMS (process 88887.369261/2019-00), GCdeM (process 88887.149245/2017-00) and WTGB (process 150911/2021-1). The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
AcknowledgmentsThe authors are grateful to the Base Aérea de Campo Grande (BACG) for the cooperation with the field works, and to the Dr. João Santana da Silva from the University of São Paulo (USP) and Dra. Carla Cardozo Pinto Arruda from the Federal University of Mato Grosso do Sul (UFMS), for providing the L. infantum and L. amazonensis positive controls respectively. The authors are especially thankful to the Insana Huna Research Group (www.insanahuna.com) for the fieldwork support.
Conflict of interestThe authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s noteAll claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
ReferencesAlves, F. M., de Lima, J. S., Rocha, F. L., Herrera, H. M., Mourão, G., de, M., et al. (2016). Complexity and multi-factoriality of Trypanosoma cruzi sylvatic cycle in coatis, Nasua nasua (Procyonidae), and triatomine bugs in the Brazilian pantanal. Parasit Vectors. 9 (1), 378. doi: 10.1186/s13071-016-1649-4
PubMed Abstract | CrossRef Full Text | Google Scholar
Antunes, T. R., Peixoto, R. A. V., Oliveira, B. B., Sorgatto, S., do Nascimento Ramos, C. A., de Souza, A. I. (2016). Detection of Leishmania infantum in peripheral blood smear and lymph node of a domestic feline. Acta Sci. Vet. 44 (Suppl 1), 162. doi: 10.22456/1679-9216.83208
CrossRef Full Text | Google Scholar
Ashford, D. A., Bozza, M., Freire, M., Miranda, J. C., Sherlock, I., Eulalio, C., et al. (1995). Comparison of the polymerase chain reaction and serology for the detection of canine visceral leishmaniasis. Am. J. Trop. Med. Hyg. 53, 251–255. doi: 10.4269/ajtmh.1995.53.251
PubMed Abstract | CrossRef Full Text | Google Scholar
Barreto, W. T. G., Herrera, H. M., de Macedo, G. C., Rucco, A. C., de Assis, W. O., Oliveira-Santos, L. G., et al. (2021). Density and survivorship of the south American coati (Nasua nasua) in urban areas in central–Western Brazil. Hystrix It. J. Mamm. 32 (1), 82–88. doi: 10.4404/hystrix-00386-2020
CrossRef Full Text | Google Scholar
Botelho, A. C., Natal, D. (2009). First epidemiological description of visceral leishmaniasis in campo grande, state of mato grosso do sul. Rev. Soc Bras. Med. Trop. 42 (5), 503–508. doi: 10.1590/s0037-86822009000500006
PubMed Abstract | CrossRef Full Text | Google Scholar
Brazuna, J. C., Silva, E. A., Brazuna, J. M., Domingos, I. H., Chaves, N., Honer, M. R., et al. (2012). Profile and geographic distribution of reported cases of visceral leishmaniasis in campo grande, state of mato grosso do sul, Brazil, from 2002 to 2009. Rev. Soc Bras. Med. Trop. 45 (5), 601–606. doi: 10.1590/s0037-86822012000500012
PubMed Abstract | CrossRef Full Text | Google Scholar
Carreira, J. C., da Silva, A. V., de Pita Pereira, D., Brazil, R. P. (2012). Natural infection of Didelphis aurita (Mammalia: Marsupialia) with Leishmania infantum in Brazil. Parasitol. Vectors. 5, 111. doi: 10.1186/1756-3305-5-111
CrossRef Full Text | Google Scholar
Carvalho, B. M., dos Santos, T. V., da R. Barata, I., Lima, J. A. N., Silveira, F. T., Vale, M. M., et al. (2018). ). entomological surveys of Lutzomyia flaviscutellata and other vectors of cutaneous leishmaniasis in municipalities with records of Leishmania amazonensis within the bragança region of pará state, brazil. J. Vector Ecol. 43 (1), 168–178. doi: 10.1111/jvec.12296
CrossRef Full Text | Google Scholar
Casanova, C., Andrighetti, M. T., Sampaio, S. M., Marcoris, M. L., Colla-Jacques, F. E., Prado, A. P. (2013). Larval breeding sites of Lutzomyia longipalpis (Diptera: Psychodidae) in visceral leishmaniasis endemic urban areas in southeastern Brazil. PloS Negl. Trop. Dis. 7 (9), e2443. doi: 10.1371/journal.pntd.0002443
PubMed Abstract | CrossRef Full Text | Google Scholar
Castro, L. S., Dorval, M. E. C., Matheus, L. M. D., Bednaski, A. V., Facco, G. G., Silveira, M., et al. (2020). Leishmania presence in bats in areas endemic for leishmaniasis in central-west Brazil. Int. J. Parasitol: Parasites Wildl. 11, 261–267. doi: 10.1016/j.ijppaw.2020.02.008
PubMed Abstract | CrossRef Full Text | Google Scholar
Cortes, S., Rolão, N., Ramada, J., Campino, L. (2004). PCR as a rapid and sensitive tool in the diagnosis of human and canine leishmaniasis using leishmania donovani s.l.-specific kinetoplastid primers. Trans. R. Soc Trop. Med. Hyg. 98 (1), 12–17. doi: 10.1016/s0035-9203(03)00002-6
PubMed Abstract | CrossRef Full Text | Google Scholar
Costa, S. M., Cechinel, M., Bandeira, V., Zannuncio, J. C., Lainson, R., Rangel, E. F. (2007). Lutzomyia (Nyssomyia) whitmani s.l. (Antunes & coutinho 1939) (Diptera: Psychodidae: Phlebotominae): geographical distribution and the epidemiology of American cutaneous leishmaniasis in Brazil – mini review. Mem. Inst. Oswaldo Cruz. 102, 149–153. doi: 10.1590/s0074-02762007005000016
PubMed Abstract | CrossRef Full Text | Google Scholar
Courtenay, O., Quinnell, R. J., Garcez, L. M., Dye, C. (2002). Low infectiousness of a wildlife host of Leishmania infantum: the crab-eating fox is not important for transmission. Parasitol 125 (Pt 5), 407–414. doi: 10.1017/s0031182002002238
CrossRef Full Text | Google Scholar
Courtenay, O., Santana, E. W., Johnson, P. J., Vasconcelos, I. A., Vasconcelos, A. W. (1996). Visceral leishmaniasis in the hoary zorro Dusicyon vetulus: a case of mistaken identity. Trans. R. Soc Trop. Med. Hyg. 90 (5), 498–502. doi: 10.1016/s0035-9203(96)90293-x
PubMed Abstract | CrossRef Full Text | Google Scholar
Cunha, R. C., Andreotti, R., Cominetti, M. C., Silva, E. A. (2014). Detection of Leishmania infantum in Lutzomyia longipalpis captured in campo grande, MS. Rev. Bras. Parasitol. Vet. 23 (2), 269–273. doi: 10.1590/S1984-29612014049
PubMed Abstract | CrossRef Full Text | Google Scholar
Cupolillo, E., Grimaldi, G., Jr, Momen, H. (1994). A general classification of new world Leishmania using numerical zymotaxonomy. Am. J. Trop. Med. Hyg. 50 (3), 296–311. doi: 10.4269/ajtmh.1994.50.296
PubMed Abstract | CrossRef Full Text | Google Scholar
da Costa, R. T., França, J. C., Mayrink, W., Nascimento, E., Genaro, O., Campos-Neto, A. (2003). Standardization of a rapid immunochromatographic test with the recombinant antigens K39 and K26 for the diagnosis of canine visceral leishmaniasis. Trans. R. Soc Trop. Med. Hyg. 97 (6), 678–682. doi: 10.1016/s0035-9203(03)80102-5
PubMed Abstract | CrossRef Full Text | Google Scholar
Dantas-Torres, F. (2007). The role of dogs as reservoirs of Leishmania parasites, with emphasis on Leishmania (Leishmania) infantum and Leishmania (Viannia) braziliensis. Vet. Parasitol. 149, 139–146. doi: 10.1016/j.vetpar.2007.07.007
PubMed Abstract | CrossRef Full Text | Google Scholar
Dantas-Torres, F., Brandão-Filho, S. P. (2006). Visceral leishmaniasis in Brazil: revisiting paradigms of epidemiology and control. Rev. Inst. Med. Trop. Sao Paulo. 48 (3), 151–156. doi: 10.1590/s0036-46652006000300007
PubMed Abstract | CrossRef Full Text | Google Scholar
Deane, L. M., Deane, M. P. (1955). Observações preliminares sobre a importância comparativa do homem, do cão e da raposa (Lycalopex vetulus) como reservatórios da Leishmania donovani em áreas endêmicas de calazar, no ceará. Hospital 48, 79–98.
de Castro Ferreira, E., Cruz, I., Cañavate, C., de Melo, L. A., Pereira, A. A., Madeira, F. A., et al. (2015). Mixed infection of Leishmania infantum and Leishmania braziliensis in rodents from endemic urban area of the new world. BMC Vet. Res. 11, 71. doi: 10.1186/s12917-015-0392-y
留言 (0)