1. IntroductionDuring cultivation, tomato plants could be infected by several pathogens, such as Verticillium dahliae (Vd), Fusarium oxysporum, Phytophthora infestans, Botrytis cinerea, and so forth. Verticillium wilt is a highly destructive soil-borne fungal disease caused by V. dahliae [
1]. The disease has changed from secondary disease to main disease in China in recent years. Heavily infected tomato plants have small fruits or no fruits, which significantly reduces tomato quality and yield, especially in a protected field. Though the Ve resistance gene against Verticillium wilt is widespread in the current tomato varieties, it confers resistance to race 1 strains of V. dahliae and not to race 2 strains [
1]. At present, there are no commercially available cultivars that are resistant to Verticillium wilt. Use of chemical pesticides is the main approach to control Verticillium wilt [
2], but chemical control harms human health and the environment, and the control effects decrease gradually due to the drug resistance of pathogens. Therefore, biocontrol of Verticillium wilt by antagonistic microorganisms has been a promising and sustainable measure because of less environment pollution and a lower health danger compared with the chemical control.Members of the genus Bacillus are ubiquitous nonpathogenic bacteria in nature. Bacillus amyloliquefaciens can be used as a biocontrol agent for a number of pathogens and as a plant growth–promoting bacterium, because it does not harm humans and animals and is eco-friendly [
3]. It produces a series of metabolites inhibiting fungal and bacterial activities, with the advantages of fast growth, strong resistance, and simple nutrition requirements. B. amyloliquefaciens is reported as a most promising bacterium in many kinds of plants to biocontrol lots of bacterial and fungal plant pathogens, including Colletotrichum orbiculare [
4], F. oxysporum [
5], Sclerotinia sclerotiorum [
6], Xanthomonas citri [
7], Pectobacterium carotovorum [
8], B. cinerea [
9], and Rhizopus oryzae [
10]. However, few studies reported a biocontrol effect and prevention mechanism of B. amyloliquefaciens on Verticillium wilt in tomato plants.
In this study, strain Oj-2.16 was isolated from rhizosphere soil of the medicinal plant Ophiopogon japonicus and identified by morphological characteristics and 16S rDNA sequencing. Its biocontrol effect on tomato Verticillium wilt and growth-promoting effect on tomato seedlings were assayed, and the activities of the tomato defense-related enzyme induced by strain Oj-2.16 were explored.
2. Materials and Methods 2.1. Materials
Fungal pathogens were provided by the key laboratory of plant–microbe interactions of Henan province, China, including V. dahliae, Botryosphaeria dothidea, F. graminearum, F. verticillioides, and F. oxysporum. Carbendazim was purchased from Sichuan Guoguang Agrochemical Co., Ltd.
2.2. Isolation and Purification of Rhizospheric Soil Bacteria of O. japonicusFive samples of rhizosphere soil were obtained from an O. japonicas field in Shangqiu, Henan Province, China. Bacteria were isolated from O. japonicas rhizosphere soil attached to roots according to a previous report [
11]. The single colonies of bacteria were picked and purified by streak culture on LB agar, and the strains were stored in a tube containing 30% (v/v) glycerol. 2.3. Determination of Bacterial Antagonistic Activity
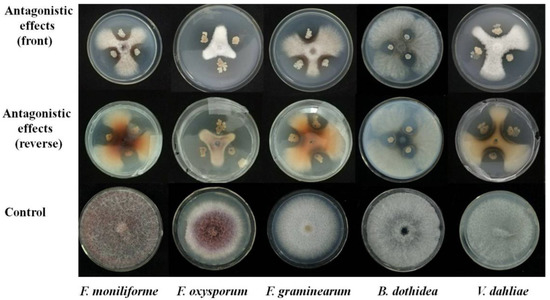
The panel confrontation way was used to screen the antagonistic activity of rhizosphere soil bacteria to different kinds of fungal plant pathogens. The 0.5 cm diameter fungus cake was set at the PDA plate center and cultivated for 24 h at 28 °C in the incubator. Bacterial strains were cultivated overnight at 37 °C in LB medium and placed to three symmetrical sites around the fungus cake on the PDA plate and with a 2.5 cm distance to the plate edge. Bacterial strains were not inoculated to the control plate. All treatments were followed by a culture in the incubator at 28 °C. The diameters of the inhibited pathogen and the control pathogen were measured after 4 days incubation. The relative inhibition rates were calculated using the formula: (diameter of the pathogen on the control plate—diameter of the pathogen on the treated plate/diameter of the pathogen on the control plate) × 100%.
2.4. Scanning Electron Microscopy Observations of the Inhibited Hyphae of Pathogenic FungiThe mycelial morphology and structural change of the pathogen in antagonistic treatment and control treatment were observed by scanning electron microscopy (Hitachi SU8010, Hitachi Higher Technologies Corp., Tokyo, Japan) as described previously [
12]. 2.5. Morphological, Physiological, and Biochemical Characteristics of Strain Oj-2.16The color and colony appearance were detected on the macromorphological level, whilst the bacterial structure was observed under a microscope. The physiological and biochemical indices were detected in terms of Bergey’s Manual of Determinative Bacteriology, including Gram staining, methyl red reaction, Voges–Proskauer reaction, gelatin liquefaction, nitrate reduction, D-glucose utilization, fructose utilization, sucrose utilization, mannitol utilization, and starch hydrolysis [
13]. IAA production was tested with the Salkowski method [
14]. Siderophore production was detected on the medium with Chrome Azurol S blue [
15]. 2.6. 16S rDNA Sequence Identification and Phylogenetic AnalysesThe genomic DNA of the bacterial strain Oj-2.16 was obtained by the rapid DNA isolation kit of the bacterial genomic (Sangon Biotech Co., Ltd., Shanghai, China) on the basis of the manufacturer’s instructions. The 16S rDNA sequences of strain Oj-2.16 were amplified, adopting the forward primer fD1 (5′-AGAGTTTGATCCTGGCTCAG-3′) and reverse primer rP1 (5′-TACCTTGTTACGACTT-3′) by PCR. The reaction was performed using 25 μL 2×Taq mixture, 5 μL DNA template, 2.5 μL forward and reverse primers, and 15 μL ddH2O. The PCR reaction conditions were 94 °C for 3 min; followed by 35 cycles of 94 °C for 30 s, annealing at 55 °C for 30 s, and extending at 72 °C for 1 min; and finally extending at 72 °C for 10 min. The PCR fragments were visualized using 1% DNA agarose gel electrophoresis. The amplified fragments were sequenced by Sangon Biotech Co., Ltd. (Shanghai, China). The obtained sequence was then submitted to GenBank. The homology analysis of 16S rDNA sequences was performed by the BLAST tool [
16]. Finally, the phylogenetic tree was built to analyze the taxonomic status of the strain by the MEGA using the neighbor joining method [
17]. 2.7. PCR Amplification of Lipopeptide and Dipeptide Biosynthetic GenesDNA was amplified by primers for lipopeptide genes, including surfactin, fengycin, iturin, and dipeptide gene bacilysin (
Table 1). The PCR mix was the same as in
Section 2.6, and the PCR reaction conditions were 94 °C for 5 min; followed by 32 cycles of 94 °C for 15 s, annealing at 55 °C for 15 s, and extending at 72 °C for 30 s/kb; and finally, extending at 72 °C for 10 min [
18]. The PCR fragments were visualized by 1% DNA agarose gel electrophoresis. 2.8. Biocontrol and Growth-Promoting Effects of the Strain Oj-2.16 on Tomato in Pot Experiments
Seeds of the tomato cultivar Moneymaker were picked and sterilized as follows: dipping in 3% NaClO for 5 min and washing three times using aseptic water. After treatment, the seeds were germinated in culture dishes with wet aseptic filter papers at 26 °C with 16 h light/8 h dark cycle per day. The germinating seedlings were planted in a pot (11 cm high, 9.5 cm in diameter) with autoclaved soil (2:1 nutritional soil: vermiculite). The pots were put in growth chamber at 26 °C with 16 h light/8 h dark. After four true leaves were formed, each pot of tomato seedlings was managed according to the following: (1) treatment by 20 mL of aseptic water; (2) treatment by 20 mL suspensions of strain Oj-2.16 (1.0 × 108 CFU mL−1); (3) first treatment by 20 mL of aseptic water and two days later, treatment by 20 mL spore suspensions (1.0 × 108 spores mL−1) of V. dahliae; (4) first treatment by 20 mL suspensions of strain Oj-2.16 and two days later, treatment by 20 mL spore suspensions of V. dahliae; (5) first treatment by 20 mL carbendazim (1 g L−1) and two days later, treatment by 20 mL spore suspensions of V. dahliae. One treatment was performed on 20 plants, each plant in one pot. Five treatments with 100 plants were placed randomly. The experiment was conducted in three independent repetitions. Statistical analyses were conducted using the Duncan’s test with SPSS V21.0.
After 20 days of V. dahliae spore inoculation, disease incidence, disease index, and biocontrol efficacy were detected as described by Chen et al. [
19]. Disease incidence (%) = [(diseased plants number)/total number of plants treated)] × 100; disease index (%) = × 100; biocontrol efficacy (%) = [(the control disease index—the treatment disease index)/the control disease index] × 100. The disease severity of Verticillium wilt in tomato plants was categorized into five grades (grade 0–4) as the criteria of disease grades presented by Wu et al. [
20], the percentage of infected tomato leaves was categorized as 0, ≤25, 26–50, 51–75, ≥76%.
The plant growth status was then assessed. The tomato plants were rooted out, and they were rinsed using running water and dried naturally. The shoot height, root length, stem width, and fresh weight of each plant were tested. The tomato plants were oven-dried for 3 days at 80 °C to test the dry weight.
2.9. Antioxidant Enzyme Activity Analysis
After 6 days of V. dahliae inoculation, antioxidant enzyme activity levels of each treatment were measured. First, 1 g of tomato leaves were picked, washed, dried, put into a precooled mortar, mixed using 8.0 mL of 0.05 mol L−1 (pH 7.8) phosphate buffer, grinded into homogenate, and centrifuged at 10,000 rpm for 15 min and 4 °C by centrifuge (Eppendorf 5804R, Eppendorf SE, Hamburg, Germany). The supernatant was obtained, which was the crude enzyme extracted from tomato leaves.
The activity of superoxide dismutase (SOD) was tested by the nitro blue tetrazolium (NBT) photoreduction method [
21], and the amount of enzyme which was required to inhibit 50% of the photoreduction of NBT was defined as one unit of SOD. The activity of peroxidase (POD) was tested by the guaiacol colorimetry method [
21], and the amount of enzyme which was required to increase the A470 value by 0.01 per minute was identified as one unit of POD. The activity of catalase (CAT) was measured by the hydrogen peroxide method [
21], and the amount of enzyme which was required to reduce the A240 value by 0.1 per minute was defined as one unit of CAT. Phenylalanine ammonia lyase (PAL) was measured by the phenylalanine method [
21], and the amount of enzyme which was required to increase the A290 value by 0.01 per minute was defined as one unit of PAL. The malondialdehyde (MDA) concentration was measured according to the thiobarbituric acid method [
22]. 4. DiscussionBiological control has been recognized as a promising and sustainable measure because of less pollution to the environment and a lower health danger compared with the chemical control. B. amyloliquefaciens with a higher antagonistic activity against phytopathogens can be successfully used as a potential biocontrol agent to control diverse crop diseases, including potato scab caused by Streptomyces griseoplanus [
24]; wheat root rot caused by Bipolaris sorokiniana [
25]; ginseng gray mold caused by B. cinerea [
26]; alfalfa anthracnose caused by C. truncatum [
27]; tomato bacterial wilt caused by Ralstonia solanacearum [
28]; tomato bacterial canker caused by Clavibacter michiganensis [
29]; wheat Fusarium head blight caused by F. graminearum [
30]; and sugar beet leaf spot disease caused by Pseudomonas syringae pv. aptata [
31]. It was also reported that B. amyloliquefaciens strains can promote the growth of plants by producing IAA hormones, siderophores, volatile organic compounds (VOCs) and improving the usability of nutrients [
32,
33,
34].The present study revealed that B. amyloliquefaciens Oj-2.16 suppressed the growth of the fungal pathogen V. dahliae and controlled Verticillium wilt in tomato. The in vitro inhibition rate of hyphal growth was 67.50% and the biocontrol efficacy of B. amyloliquefaciens Oj-2.16 was 89.26%, which was slightly higher than the efficacy of the chemical fungicide carbendazim in pot experiments. B. amyloliquefaciens is one of the most prevalently studied biocontrol agents. However, only limited experiments have been performed to biocontrol Verticillium wilt by Bacillus. Similar results were obtained by the utilization of B. atrophaeus to impart resistance to Verticillium wilt of cotton [
35] and B. velezensis to control Verticillium wilt of olive trees [
36].In addition, inoculation with B. amyloliquefaciens Oj-2.16 significantly increased shoot height, root length, stem width, fresh weight, and dry weight of tomato seedlings compared with the control, attributed to the features of strain Oj-2.16 to produce IAA, siderophores, assimilate various carbon sources, including glucose, fructose, and mannitol. These results showed that the antagonistic strain Oj-2.16 could also serve as a plant growth stimulator or biofertilizer, which could be superior to the chemical fungicide carbendazim that does not have the ability of promoting growth. Ji et al. [
31] reported that B. amyloliquefaciens Ba13 significantly increased tomato plant height and fresh weight by 10.98% and 20.15%, respectively, which were lower than those increased by B. amyloliquefaciens Oj-2.16, namely 44.44% and 57.65%, respectively.B. amyloliquefaciens is well known for its efficacy in the biocontrol of plant diseases, which involves mechanisms like secretion of antimicrobial substances, competition for nutrients and ecological niche, induction of host systemic resistance (ISR), growth promotion, and enhancement of colonization ability [
37,
38]. Induction of the enzyme activities related to plant defense is a form of systemic resistance, which plays key roles in disease resistance response of plants [
39,
40]. Our results showed that B. amyloliquefaciens Oj-2.16 triggered tomato plant resistance to Verticillium wilt by increasing the activities of defense-related antioxidant enzymes. POD, SOD, CAT, and PAL enzyme activities significantly increased in the plants treated with strain Oj-2.16 upon inoculation of V. dahliae compared with the pathogen-inoculated control. The enhanced activities of antioxidant enzymes can effectively scavenge harmful reactive oxygen species to maintain the stability of plant defense systems, so the treated tomato returned to normal growth (
Figure 5). MDA content is the index of the level of membrane-lipid peroxidation. Compared with such value under the mere Vd pathogen-inoculated control, MDA content under V. dahliae inoculation and B. amyloliquefaciens Oj-2.16 treatment was significantly lower, indicating a lower damage degree of the membrane system.Scanning electron microscopy observation revealed that the hyphae of V. dahliae were destroyed to be dehydrated, distorted, dissociated and forming hollows, after fungi were treated with B. amyloliquefaciens Oj-2.16 spore suspension in panel confrontation experiments. B. amyloliquefaciens might secrete some antimicrobial substances, such as antimicrobial proteins and peptides, which can damage the mycelial structure, including deformation, expanding hyphae, and leaking protoplasm [
26,
41]. Genes involved in antifungal lipopeptide and dipeptide biosynthesis were detected in B. amyloliquefaciens strain Oj-2.16, suggesting that Oj-2.16 could produce fengycin, surfactin, iturin, and bacilysin. To explore its biocontrol mechanism and broaden its application, the expression, isolation, and function identification of peptide products produced by strain Oj-2.16 in the process of controlling tomato Verticillium wilt need to be studied further.
In summary, B. amyloliquefaciens Oj-2.16 was firstly isolated from rhizosphere soil of O. japonicas with a broad antagonistic activity against V. dahliae, B. dothidea, F. oxysporum, F. graminearum, and F. moniliforme. B. amyloliquefaciens Oj-2.16 can effectively biocontrol tomato Verticillium wilt disease and promote tomato growth. Strain Oj-2.16 can produce IAA, siderophores, assimilate various carbon sources including glucose, fructose, and mannitol, all of the features contributed to promoting growth. B. amyloliquefaciens Oj-2.16 triggered tomato plant resistance to Verticillium wilt by increasing the levels of defense-related antioxidant enzymes POD, SOD, CAT, and PAL. Induction of the activities of plant defense enzymes is a form of systemic resistance, which is the main antifungal mechanism to suppress tomato Verticillium wilt disease.


留言 (0)