
記住我
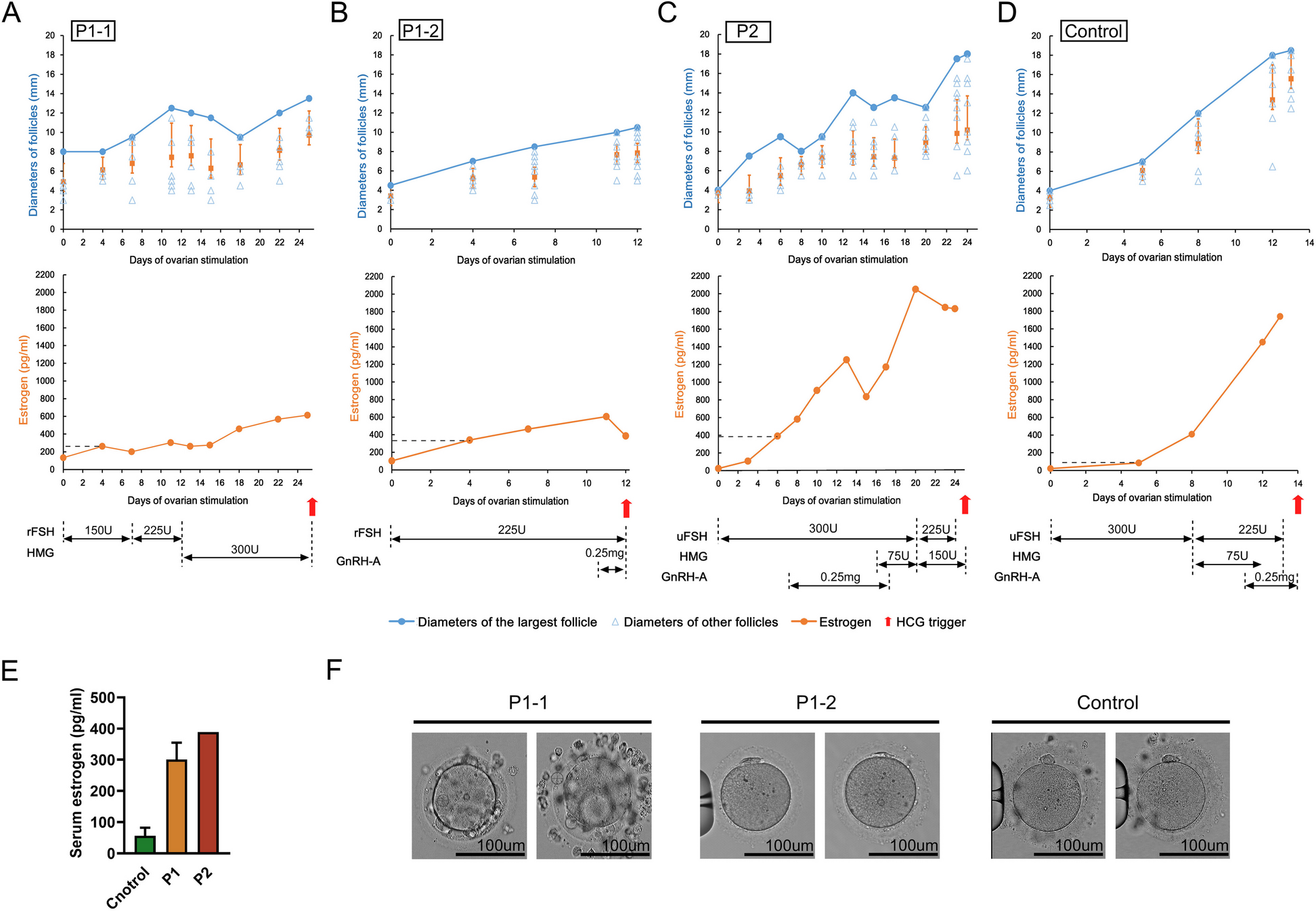



It has been well documented that increased ROS and diminished antioxidant capacity induce cellular senescence, and catalase enzymes have been used to alleviate senescence through its antioxidant defense mechanism [6, 8, 20]. We investigated whether catalase is used to alleviate ROS and diminish aging in catalase-deficient mice. For this, WT and catalase-KO mouse embryonic fibroblasts cells (MEFs) were isolated and cultured. Catalase deficiency was confirmed in KO MEFs by immunoblot analysis with anti-catalase (Additional file 1: Fig. S1a). KO MEFs displayed flattened and enlarged senescence phenotypic morphology at early passage (P2) and showed increased senescence phenotypic morphology with increasing passage (P4) (Additional file 1: Fig. S1b). To determine whether the flattened and enlarged MEFs were senescent, β-galactosidase staining was performed. As expected, KO MEFs showed positive staining for senescence-associated β-galactosidase, which significantly increased in P5 but not in WT cells (Fig. 1A and Additional file 1: Fig. S1c). To illustrate the senescence-induced phenotype, immunoblot analysis was performed for WT and KO MEFs from P2 and P5. Notably, the expression levels of senescence-related proteins p21 and p16 increased in KO P5 MEFs (Fig. 1B–D). Furthermore, to confirm the aging phenotype, catalase-KO mice with WT littermates at the age of 9 weeks (9 W) and 53 weeks (53 W) were subjected to the experiment. Partial hair in the dorsal back skin of 53 W KO mice turn gray whereas it looks normal in WT mice (Fig. 1E, indicated by red arrow). β-galactosidase staining was performed in the liver of mice, which showed positive staining in KO mice livers at 53 W (Fig. 1F). Immunoblot analysis of liver homogenates from mice showed the induction of senescence-related proteins in KO mice at 53 W (Fig. 1G–I). Together, these data suggest that catalase deficiency induces an aging phenotype faster than WT mice.
Fig. 1

Catalase-deficient mice induce aging phenotype faster than WT mice. A Morphology and senescence-associated β-galactosidase staining of WT and catalase KO MEFs at passage 2 (P2) and passage 5 (P5) levels. Intensities of senescent cells at each passage, P2 and P5, in WT and KO MEFs. Positive intensities of β-galactosidase staining were measured compared to control using the ImageJ software. Scale bar represents 100 μm. B Proteins were extracted from MEFs at P2 and P5 level. Immunoblot analysis was performed using whole-cell lysates with indicated senescence-associated antibody. C, D quantified protein level of P21 and P16 normalized with actin. E Morphology of dorsal back hair of WT and KO mice at 53 W. F Senescence-associated β-galactosidase staining was performed in the liver section of 9 weeks (9 W) and 53 weeks (53 W) WT and catalase-KO mice. Intensities of senescent cells. Positive intensities of β-galactosidase staining were measured compared to control using the ImageJ software. Scale bar represents 100 μm. G Immunoblot analysis was performed from the liver sample from WT and catalase-KO mice at the age of 9 W and 53 W with indicated antibody. H, I quantified protein level of P21 and P16 normalized with actin. Bar graph represents mean ± SD (n = 3 experiments). *P < 0.05; **P < 0.0001; ns: non-significant
Catalase-deficient mice induce ROS through mitochondria and peroxisomeAs reported earlier, ROS generation is the main cause of aging due to decreased cellular antioxidant capacity [6, 8, 20]. We hypothesized that ROS generation might be increased in catalase-KO mice. As expected, the fluorescence intensity measured by 2′7′-dichlorofluorescein diacetate (DCFH-DA) staining showed an increase in catalase-KO MEFs at P5 (Fig. 2A). The fluorescent signal of DCFH-DA staining, representing ROS, was quantified and showed that ROS generation was significantly higher in catalase-KO cells at P5 (Fig. 2A). Likewise, we examined the expression of another ROS marker, 4 hydroxynonenal (4-HNE), in MEFs by immunofluorescence staining (IF). Consistent with DCFH-DA staining, the fluorescence intensity of 4-HNE in KO MEFs increased at P5 (Fig. 2B). In addition, to confirm ROS generation in vivo, intracellular ROS levels were measured in liver lysates of mice. Total ROS levels increased significantly in KO mice at 53 W (Fig. 2C). The endogenous source of ROS contains different cellular locations, with mitochondria and peroxisomes being the major sites [2, 21, 22]. The principal source of ROS produced by mitochondria is the superoxide anion, a byproduct of the electron transport chain responsible for oxidative damage by aerobic energy metabolism [23, 24]. To detect mitochondrial ROS, MitoSOX red, a mitochondrial superoxide indicator, was used in WT and KO MEFs at the P2 and P5 levels (Fig. 2D). Catalase-KO cells at P5 showed increased levels of the red fluorescence signal. In addition, in vivo, the level of ACOX1 (acyl-CoA oxidase 1), a major producer of ROS in the peroxisome and the first and rate-limiting enzyme in fatty acid β-oxidation, increased significantly in KO mice at 53 W (Fig. 2E). To confirm the induction of ROS in catalase-KO mice, MEFs were treated with the antioxidant N-acetyl-L-cysteine (NAC). Treatment with NAC, however, inhibited the level of ROS generation in KO MEFs at P5 (Fig. 2A, B, and D). Taken together, these data suggest that catalase-KO mice induce ROS production through both mitochondria and peroxisomes. Generation of ROS is susceptible to lipid oxidation and protein carbonylation that lead to DNA degradation, indicating nucleic acid oxidative damage [25]. DNA released from nucleus to cytosol after oxidative damage contributes to senescence [26]. Based on this, Using double-stranded DNA (dsDNA) antibodies to detect DNA by immunofluorescence (IF), we found that almost 30% of KO MEFs at P5 exhibited extra nuclear DNA in contrast to 15 and 5% in WT P5,P2 and KO P5 cells (Fig. 2F).Taken together, it shows that extranuclear DNA in KO MEFs regulate aging through ROS.
Fig. 2

Aging is induced in catalase-deficient mice through ROS generation. A Representative fluorescence images of DCFH-DA staining of WT and catalase-KO MEFs at P2 and P5 levels. Quantification of cells showing green fluorescence (corresponding to DCFH-DA) and fluorescence intensity of MEFs. *P < 0.05 WT P5 versus KO P5; KO P5 versus KO P5 + NAC. B Representative fluorescence images of MEFs fixed and immunostained with anti-4HNE (green). Scale bar represents 20 μm. C Total ROS was measured in liver lysates of WT and catalase-KO mice at 9 W and 53 W. Values represent mean ± SD (n = 3, 4). *P < 0.05 WT 53 W versus KO 53 W. D Representative (red) fluorescence image of MitoSOX staining in MEFs, as in A. E ACOX1 levels were measured using the ACOX1 ELISA kit from the liver lysates of mice as in C. F IF staining of anti-dsDNA (red), DAPI (blue) in MEFs indicated as in A. Insets(*,#, $) enlarged cells; scale bar, 20 μm. Intensities of dsDNA positive cells at each passage, P2 and P5, in WT and KO MEFs.G MEFs at P2 and P5 were treated with 5 mM NAC overnight, and senescence-associated β-galactosidase staining was analyzed. The percentage of senescent cells was analyzed in WT and KO MEFs treated with NAC. Positive intensities of β-galactosidase staining were measured using the ImageJ software. Bar graph represents mean ± SD (n = 3 experiments). Scale bar represents 100 μm. *P < 0.05, WT P2 versus KO P2; KO P2 versus KO P2 + NAC; WT P5 versus KO P5. KO P5 versus KO P5 + NAC. H Proteins were extracted from MEFs as in G. Immunoblot analysis was performed using whole-cell lysates with indicated senescence-associated antibodies. I, J quantified protein level of P21 and P16 normalized with actin. Bar graph represents mean ± SD (n = 3 experiments). *P < 0.05; **P < 0.0001
We hypothesized that ROS generation in catalase-KO mice may induce cellular senescence, as previously described [6, 8, 20]. Hence, β-galactosidase staining was again performed in WT and catalase-KO MEFs and co-treated with NAC (Fig. 2G). As expected, positive staining of senescence-associated β-galactosidase in KO MEFs was significantly diminished by treatment with NAC. Moreover, NAC treatment also decreased senescence-related protein in KO P5 MEFs (Fig. 2H–J). Together, these data suggest that catalase deficiency induces an aging phenotype through ROS generation.
Catalase-deficient mature adult mice induce leaky lysosomeLysosomes are the main catabolic organelle that play an essential role in cellular process, including response to nutrient availability, stress resistance, plasma membrane repair development, and cellular differentiation [27]. In line with catabolic organelles, lysosomal activity is strongly influenced by aging by altering the physical and chemical properties of these organelles and rendering them more sensitive to stress [12]. Considering this notion, immunoblot analysis of mice liver homogenates was performed to check the lysosomal marker protein LAMP1 (Fig. 3A). Lysosomal protein levels was significantly increased in both the mice liver at the age of 53 W (Fig. 3A, B). Aging has also been reported to increase lysosomal volume [28]. Hence, to check the volume, the morphology of lysosomes was analyzed using immunofluorescence (IF). WT and KO MEFs were immunostained with the lysosomal marker LAMP1 (Fig. 3C). Catalase-deficient MEFs at the P5 level showed an enlarged cellular size, which significantly increased the red fluorescence signal toward the cytoplasm of the cell. To confirm the lysosomal area, the fluorescent signal of LAMP1 was quantified and showed that lysosomal area was significantly higher in catalase-KO MEFs at P5 (Fig. 3D). Again we questioned, did increased in the lysosomal area affect the acidity of lysosme? Hence to confirm this, MEFs were immunostained with lysotracker for labeling and tracking of the acidotropic probe for lysosomes (Fig. 3E). The fluorescence intensity of acidic vesicle specifically accumulating in lysosomes decreased, in KO MEFs at the P5 level (Fig. 3E). Lysosomal activity is highly influenced by hydrolytic enzymes residing in the lumen of the lysosomal membrane, which is highly acidic [19]. As the acidotropic probe for lysosomes decreased in KO MEFs, we assumed that the resident hydrolytic enzymes in the lysosomal lumen were leaked. Hence, leaky lysosomal content may make the hydrolytic enzyme alkaline, causing lysosomes to fuse in the cytosol, which may increase lysosomal size. To check the lysosomal content, we analyzed the protein levels of cathepsin D (cathD) and cathepsin B (cathB), two major lysosomal hydrolases that can serve as molecular reporters for lysosomal functions, in WT and KO liver homogenates (Fig. 3F). Both lysosomal hydrolases were accumulated in the supernatant fraction in KO mice at 53 W, whereas both cath D and B were normal in the pellet fraction in the other groups (Fig. 3F–H). Again LAMP1 protein was significantly increased in the pellet of KO mice liver at the age of 53 W (Fig. 3F, I). Furthermore, cath D activity was measured in the liver lysates of mice (Fig. 3J). cath D activity significantly decreased in KO mice at 53 W. Together this data confirmed that catalase KO mice at 53 W induce leaky lysosome causing accumulation of lysosomal hydrolases in the cytoplasm that make hydrolytic enzyme alkaline and increase lysosomal volume towards the cytoplasm.
Fig. 3

Catalase-deficient mature adult mice induce leaky lysosomes. A Protein was extracted from liver lysates from WT and catalase-KO mice at 9 W and 53 W.Immunoblot analysis was performed using whole-cell lysates with LAMP1 antibody. B Quantified protein level of LAMP1 normalized with actin. C Representative fluorescence images of MEFs fixed and immunostained with anti-LAMP1 (red) and DAPI (blue). Scale bar represents 20 μm. D Intensities of lysosomal area showing red fluorescence (corresponding to control) and fluorescence intensity of MEFs. E Representative fluorescence images of lysotracker for WT and catalase-KO MEFs at P2 and P5 levels. Scale bar represents 20 μm. F Liver lysates from WT and catalase-KO mice at 9 W and 53 W were fractioned as described in the Materials and Methods. Equivalent volumes of each fraction were subjected to immunoblotting using the indicated antibodies. G–I Quantified protein level of cathD, cathB and LAMP1 on pellet and supernatant fractionations, normalized with actin. J Relative cath D levels were measured from liver lysates of mice, as indicated in A. Bar graph represents mean ± SD (n = 3 experiments). *P < 0.05; **P < 0.0001; ns: non-significant
Leaky lysosome persuades alteration of autophagy through ROS in catalase-deficient mature adult miceDamaged lysosomes are selectively sequestered by autophagy [19]. Hence, we hypothesized that damaged lysosomes in KO mice may be recruited by autophagy machinery, which are then engulfed by autophagosomes. To confirm the autophagic process, MEFs were co-immunostained with LC3, an autophagy marker, with lysotracker (Fig. 4A). In contrast to our assumption, the LC3 positive puncta green fluorescence decreased and were not fused with lysosomes in KO MEFs due to decreased lysosomal acidic probe, which showed less fusion of autophagosomes with lysosomes (Fig. 4A–D). Further, immunoblot analysis was performed in WT and KO MEFs at the P2 and P5 levels. Notably, the expression levels of autophagy substrate marker P62 and autophagosome by LC3II were significantly increased in KO MEFs at P5 (Fig. 4E). To confirm autophagic dysregulation in vivo, liver tissues from mice were immunoblotted with an autophagy marker (Fig. 4F). Consistent with MEFs, liver tissues from KO mice at 53 W showed dysregulation of basal autophagy in comparison to WT and KO 9 W livers (Fig. 4F). It’s known that ROS mediate leaky lysosomes [29]. Hence, we hypothesize that ROS might be the main player to dysregulate autophagy through leaky lysosome. Hence, to confirm this, WT and catalase-KO MEFs were treated with or without NAC at both passages (P2 and P5). Treatment with NAC slightly recovered the acidic probe of lysosomes, which fused with autophagosomes in KO MEFs at the P5 level (Fig. 4A–E). Hence, these data suggest that leaky lysosomes alter autophagy through ROS in catalase-deficient mature adult mice.
Fig. 4

Catalase-deficient mature adult mice induce alteration of autophagy through ROS. A MEFs from WT and catalase-KO mice at P2 and P5 levels were treated with 5 mM NAC, fixed, and immunostained with anti-LC3 (green),Lysotracker (red), yellow showed LC3 puncta co-localized to lysosme (arrow head). Scale bar represents 10 μm. Quantification of B LC3, C lysotracker and D co-localized cell analyzed from each experimental group as in A. The bar graph represents the mean ± SD (n = 3 experiments). *P < 0.05; **P < 0.0001. E Proteins were extracted from MEFs as in A. Immunoblot analysis was performed using whole-cell lysates with the indicated antibodies. F Protein was extracted from liver lysates from WT and catalase-KO mice at 9 W and 53 W. Immunoblot analysis was performed using whole-cell lysates with the indicated antibodies
Leaky lysosomes induce lipofuscin accumulation through ROS in catalase-deficient mature adult miceAlthough leaky lysosomes are induced in catalase-deficient MEFs, the damaged lysosomes were not degraded through the autophagic process. Instead, basal autophagy was dysregulated in KO MEFs. To find the mechanistic evidence, we examined the morphology of the liver by H&E staining in mice. The liver morphology of 53 W KO mice showed microvesicular steatosis (i.e., accumulation of small fat droplets) in the cytosol of hepatocytes (arrows), with brown pigment (arrowhead), whereas the livers of WT and KO 9 W mice showed normal lobular architecture with hepatocytes arranged in hepatic cords (Fig. 5A). To confirm the accumulation of fat droplets in hepatocytes, triacylglycerol (TG) was measured in the liver lysates of mice. Consistent with H&E staining, liver lysates of 53 W KO mice showed a significant increase in liver TG compared to other group (Fig. 5B). Furthermore, oil red O staining (ORO) also showed the induction of lipid droplets in the hepatocytes of 53 W liver sections (Fig. 5C). During aging, the volume and structure of hepatocyte organelles change [30]. Although we observed a significant increase in body weight of 53 W old KO mice, but there were no significant changes in liver weight (Additional file 1: Fig. S2a, b). We hypothesized that the accumulation of small lipid droplets in the cytoplasm of hepatocytes in KO 53 W old mice may be an undigested lipid, lipofuscin, which showed brown pigmentation on hepatocytes (Fig. 5A, white arrow head). Lipofuscin is a highly oxidized insoluble protein that fails to degrade damaged and denatured proteins [31]. Moreover, it is a chemically and morphologically polymorphous waste material that accumulates at the primary site of the lysosome and disturbs lysosomal degradation and causes lysosome leakage [32, 33]. To examine the accumulation of lipofuscin or leaky lysosomes, MEFs were co-immunostained with LGALS1 (galectin-1), a leaky lysosome marker with lysotracker (Fig. 5D). Catalase-KO MEFs at P5 level showed a significant increase in LGALS1 puncta that were loaded on the lysosomes (Fig. 5D, arrow); although acidic vesicles in lysosomes by lysotracker were less, almost all LGALS1 puncta were localized to lysosomes. In contrast, LGALS1 puncta were less or not observed at all in the WT at P2, P5, and KO P2 levels. It is known that enhanced ROS results in the leakage and accumulation of lipofuscin in lysosomes [29, 33]. Hence, to confirm this, WT and catalase-KO MEFs were treated with or without NAC at both passages (P2 and P5). Treatment with NAC slightly rescued the acidic vesicles of lysosomes and decreased the localization of LGALS1 puncta to the lysosomes in KO MEFs at P5 level (Fig. 5D). Together, these data suggest that leaky lysosomes induce lipofuscin accumulation through ROS in catalase-KO mature adult mice.
Fig. 5
Catalase-deficient mature adult mice induce lipofuscin accumulation through ROS. A Representative hematoxylin and eosin (H&E) staining of livers from WT and catalase-KO mice at the age of 9 W and 53 W, respectively. The livers of 53 W KO mice showed microvesicular steatosis (i.e., accumulation of small fat droplets) in the cytosol of hepatocytes (arrows) with a golden-brown pigment (arrowhead) known as lipofuscin. Scale bar represents 50 μm. B Liver samples from mice as mentioned in A were homogenized, and TG levels were analyzed. Bar graph represents mean ± SD (n = 3 experiments). *P < 0.05, WT 53 W versus KO 53 W. C Cryosectioned liver tissues from mice were stained with ORO. Scale bar represents 50 μm. D Representative fluorescence images of MEFs at P2 and P5 treated with 5 mM NAC, fixed, and immunostained with Lysotracker (red), LGALS1 (green), and DAPI (blue). Scale bar represents 5 μm
Leaky lysosome affects lysosomal pH that activates mTORC1 and leads to cellular senescenceNext, we constructed leaky lysosomes using the well-known lysosomal membrane permeabilization (LMP) marker L-leucyl-L-leucine methyl ester (LLOME) in hepatoma cells and questioned whether leaky lysosomes induce ROS and cellular senescence. For this, we treated HepG2 cell with LMP inducer LLOME for 24 h and examined the morphology of lysosomes by immunostaining with lysotracker. As expected, the specific accumulation of acidic vesicles in lysosomes decreased in LLOME-treated cells (Fig. 6A). The disruption of acidic hydrolases in lysosomes or leaky lysosomes is induced through extensive ROS [29]. To confirm this, HepG2 cells were stained with DCFH-DA (Fig. 6B). Fluorescence intensity by DCFH-DA staining increased in LLOME-treated cells. Hence, to confirm the leaky lysosomes induced by ROS accumulation, HepG2 cells were co-treated with antioxidant NAC in LLOME treated cells. As expected, NAC significantly recovered the acidic vesicles of lysosomes and inhibited DCFH-DA fluorescence intensity in HepG2 cells (Fig. 6A–B). LLOME treatment increased the cytosolic release of lysosomal hydrolases [19]. Hence, to check the lysosomal content, the protein levels of cath D and B were immunoblotted in HepG2 cells treated with LLOME (Fig. 6C). As expected, both lysosomal hydrolases (cath D and B) accumulated in the supernatant fraction in LLOME-treated cells, whereas they were normal in the pellet fraction of untreated cells (Fig. 6C–E). Also the protein level of lysosome significantly increased in LLOME treated cell (Fig. 6C, F). Further immunoblot analysis of HepG2 cells showed increased protein expression of the ROS marker 4-HNE and peroxisomal oxidase ACOX1 in LLOME-treated cells (Additional file 1: Fig. S3a). However, we did not observe any changes in mitochondrial enzymes, including COX1, COX4, voltage-dependent anion channel (VDAC), and antioxidant proteins, including SOD1 and SOD2.
Fig. 6

Catalase-deficient mature adult mice induce mTORC1 activation. A HepG2 cells were treated with 1 mM LLOMe for 24 h and co-treated with 5 mM NAC overnight. Representative fluorescence images of HepG2 cells immunostained with Lysotracker (red) and DAPI (blue). Scale bar represents 5 μm. B Representative fluorescence images of DCFH-DA staining of HepG2 cells treated as in a. Scale bar represents 20 μm. C Lysates of HepG2 cells treated as in A were fractioned as explained in the Materials and Methods section. Equivalent volumes of each fraction were subjected to immunoblotting using the indicated antibodies. D–F Quantified protein level of cathD, cathB and LAMP1 normalized with actin. G Representative fluorescence image of HepG2 cells treated as in A, immunostained with anti-TFEB (red) and DAPI (blue). Scale bar represents 20 μm. Quantified bar represents percentage of nuclear localization of TFEB on treated cell as indicated. H Protein was extracted from HepG2 cells as in a. Immunoblot analysis was performed using whole-cell lysates with indicated antibodies. Bar graph represents mean ± SD (n = 3 experiments). *P < 0.05; **P < 0.0001
The link between decreased lysosomal function and aging has been well studied [12, 34]. Hence, we investigated whether LMP drug LLOME induces cellular senescence in cells. β-galactosidase staining was performed in HepG2 cells. As expected, cells treated with LLOME showed positive staining for senescence-associated β-galactosidase (Additional file 1: Fig. S3b). However, treatment with NAC inhibited the positive staining of senescence-associated β-galactosidase. Further immunoblot analysis was performed in HepG2 cells showing increased expression of aging-related proteins, including p16 and p21, in LLOME-treated cells (Additional file 1: Fig. S3c). During lysosomal damage, transcription factor EB (TFEB), a major regulator of autophagy and lysosomal biogenesis, has been shown to rapidly translocate to the nucleus and activate the transcription of its target gene for the activation of lysosomes [35]. Hence, immunostaining was performed for translocation of TFEB to HepG2 cells. As expected, LLOME-treated cells showed translocation of TFEB to the nucleus in LLOME-treated cells (Fig. 6d). Hence, lysosomal rupture induces the biogenesis of lysosomes through autophagy (Additional file 1: Fig. S3d), as previously described [19, 35]. Meanwhile, immunoblot analysis also showed that treatment with LLOME increased the expression of phosphorylated S6 (pS6), a marker of downregulation of mTORC1 (Fig. 6H). Further immunostaining with anti-mTORC1was performed in LLOME-treated cells, which showed increased expression of mTORC1 protein (Additional file 1: Fig. S3e). Together, these data show that leaky lysosomes affect lysosomes and induce cellular senescence probably through mTORC1 activation.
Rapamycin attenuated cellular senescence induced by catalase-deficient cellsmTOR is a key component of cellular metabolism that promotes cell growth and proliferation via nutrient sensing. In addition to cellular growth and proliferation, mTOR has also been associated as a lifespan regulator in mice [36,37,38,39]. The lifespan-enhancing effects of rapamycin have been linked to mTORC1 inhibition [40]. Hence, we treated cells with rapamycin, an mTORC1 inhibitor, to reverse cellular senescence induced by catalase-KO mice. For this, WT and KO MEFs were treated with rapamycin, and β-galactosidase staining was performed to check the senescence phenotype. KO MEFs displayed flattened and enlarged senescence phenotypic morphology at early passage (P2) and showed increased senescence phenotypic morphology with increasing passage (P5) in addition to positive staining for senescence-associated β-galactosidase, but not in WT MEFs (Fig. 7A). However, treatment with rapamycin inhibited positive staining of senescence-associated β-galactosidase, but KO MEFs still displayed flattened and enlarged senescence phenotypic morphology. Further immunoblot analysis of MEFs showed the induction of senescence-related proteins,P21 and P16 in KO cells that was suppressed by co-treatment with rapamycin (Fig. 7B–D). mTORC1 is also known to suppress autophagy, and activation of autophagy by suppression of mTORC1 can slow age by clearing the accumulating old and dysfunctional organelles [40, 41]. Hence, to check the clearance of old and dysfunctional organelles by autophagy, rapamycin was used to treat MEFs, and immunoblot analysis was performed. Remarkably, the increased levels of autophagy substrate marker P62 and autophagosome marker LC3II were significantly decreased by rapamycin in KO MEFs at P5 level (Fig. 7E). Furthermore, the mTORC1 marker protein pS6 was decreased by rapamycin, suggesting that autophagy was initiated (Fig. 7E). In addition, accumulation of lysosomal content, such as cath D, in the supernatant fraction in KO MEFs at level P5 was slightly suppressed by rapamycin (Fig. 7F, G). Similarly, cath D activity was also recovered in KO MEF following treatment with rapamycin (Fig.
留言 (0)