
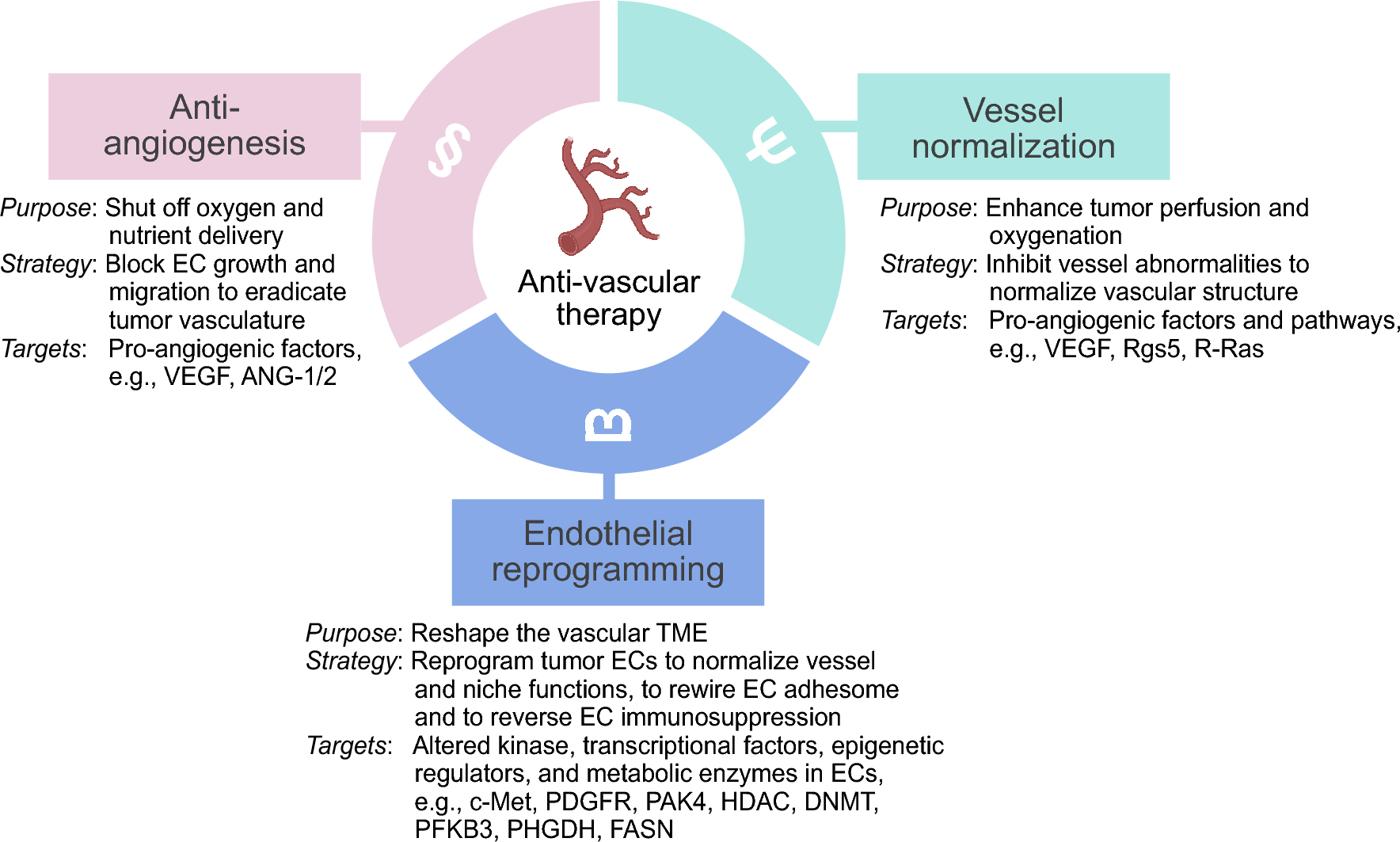
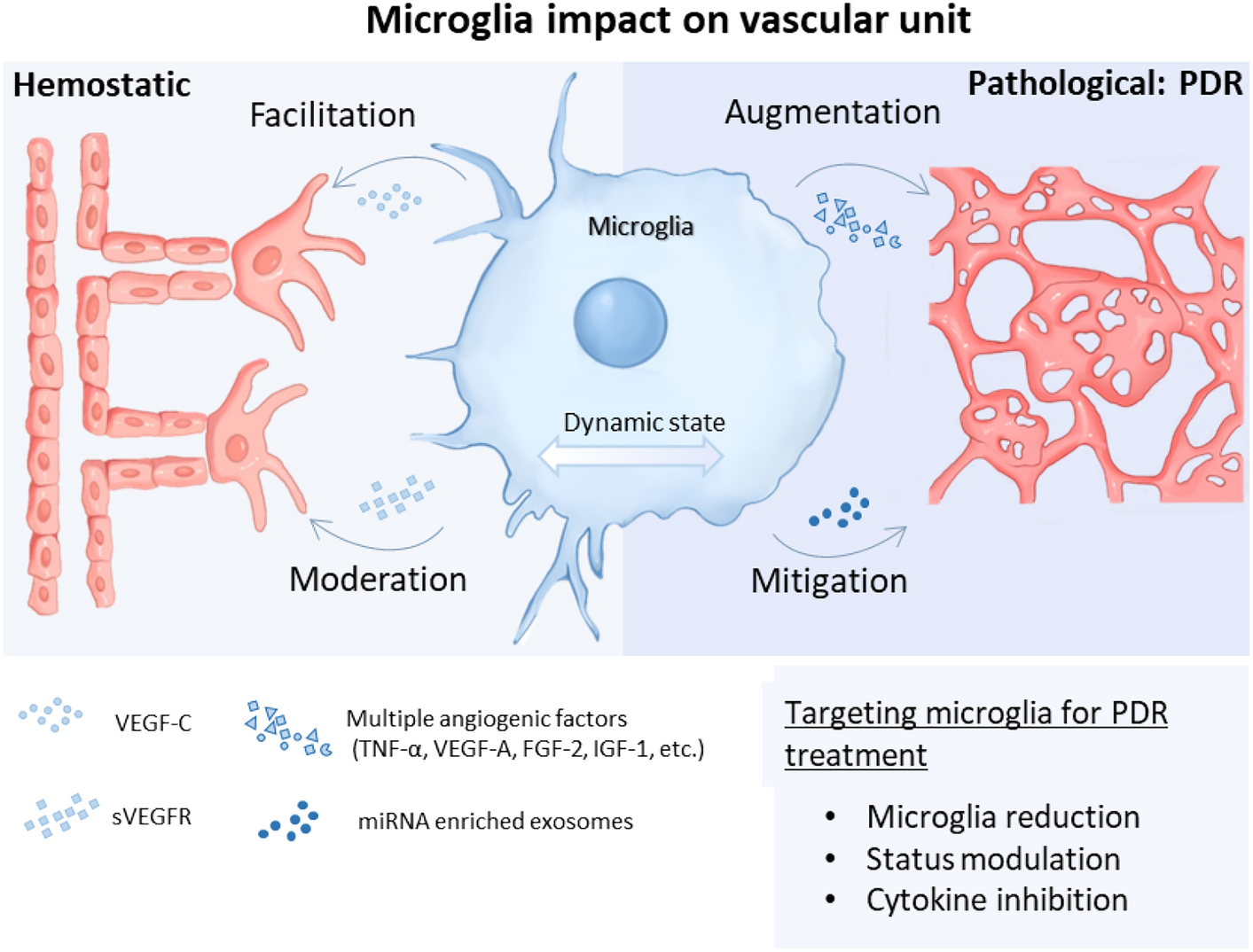
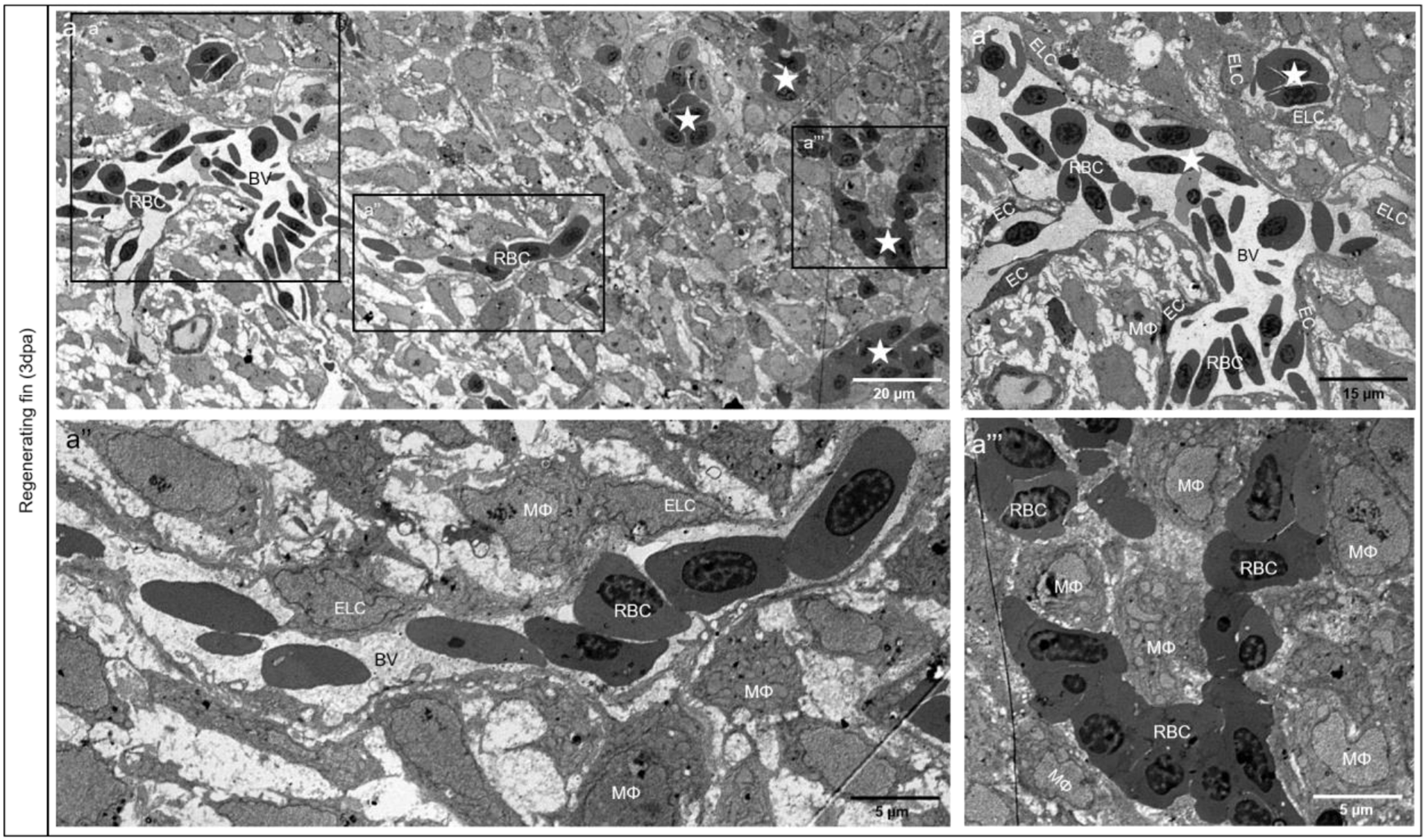
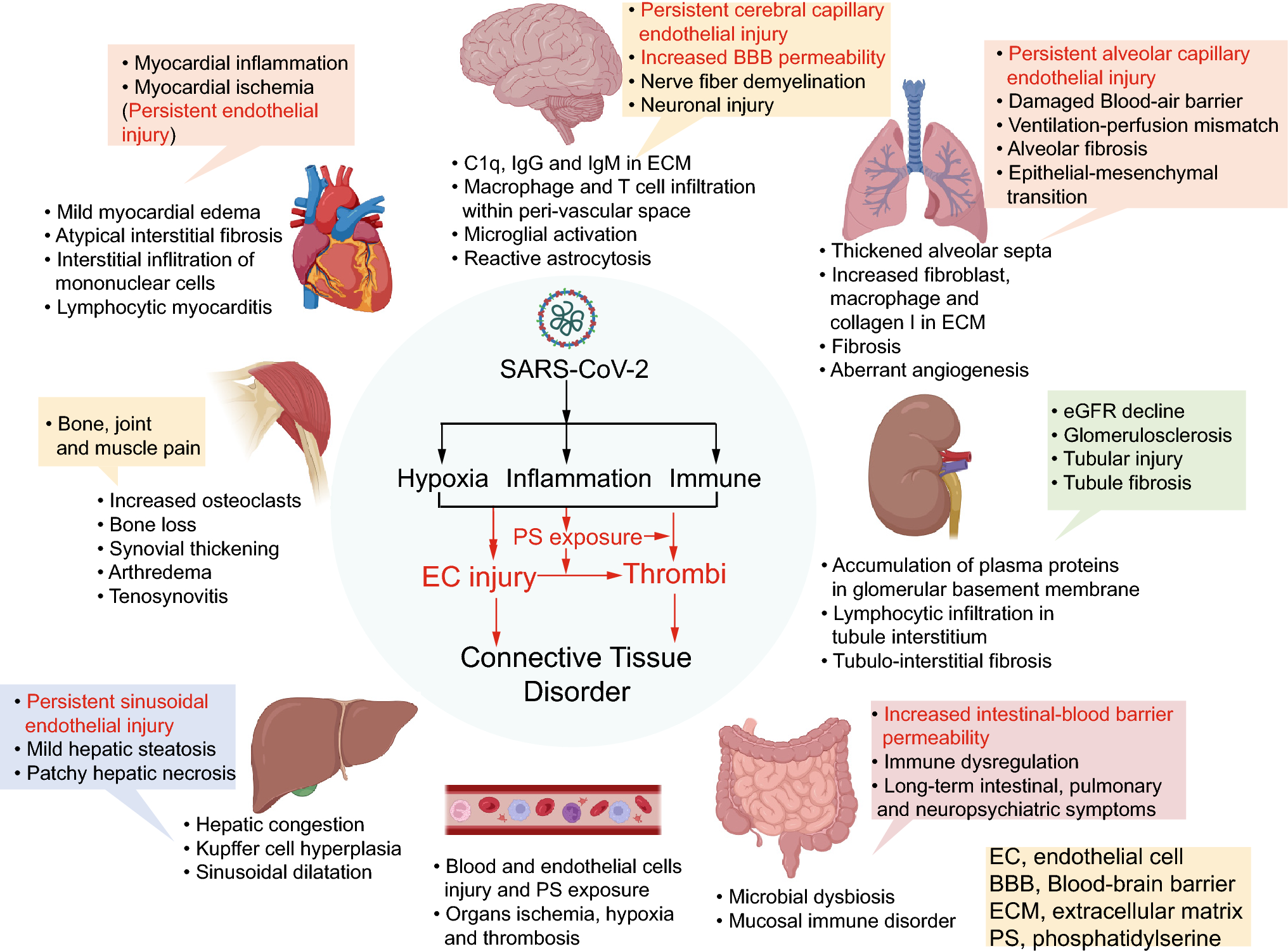
記住我


Conventional light microscopy of COVID-19 heart samples showed minor inflammatory infiltrates consisting mainly of macrophages in only 1 of 24 cases. The Influenza group showed no significant inflammatory infiltration in all 16 analyzed cases using light microscopy. In contrast, non-influenza/non-SARS-CoV-2 myocarditis cases revealed lymphocytic infiltrates, with minor infiltrates in 4 of the 8 cases, severe lymphocytic infiltrates in 3 of the 8 cases and 1 of the 8 cases with only minimal sparse inflammatory infiltrate with adjacent single-cell necrosis, all but one fulfilling the Dallas criteria for myocarditis (Fig. 1A), while no necrosis was observed in COVID-19, influenza or control tissue. Regarding interstitial fibrosis, no statistically significant (Chi-square test p = 0.45) difference was found between the four groups, with COVID-19 (mean score 0.85) displaying the highest and the Influenza group (mean score 0.6) displaying the lowest fibrotic remodeling, respectively (Fig. 1A, B). Cardiomyocyte hypertrophy was most pronounced in the COVID-19 group (mean score 1.2), followed by control cases (mean score 1.1), and with influenza (mean score 0.4) and common viral myocarditis cases (mean score 0.8) showing the least amount of hypertrophy (combined Chi-square test p < 0.0001). However, COVID-19 patients were statistically significantly older (73.7 ± 10.8 years) compared to influenza (52.3 ± 15.3 years, p < 0.001), common myocarditis (54.8 ± 21.3; p < 0.05), and non-inflamed control patients (54.9 ± 18.0, p < 0.05). Immunohistochemistry for SARS-CoV-2 spike and nucleocapsid protein as well as RNA-FISH for SARS-CoV-2 RNA showed no specific signal in the analyzed COVID-19 tissues. However, RT-PCR for SARS-CoV-2 RNA yielded positive results in 17 of the 24 analyzed samples (supplementary Table 1).
Fig. 1
A Histological comparison of COVID-19, influenza, and lymphocytic non-influenza myocarditis to non-infected controls. The myocardium of a COVID-19 patient shows no inflammatory infiltrate or cardiomyocyte necrosis with minor interstitial fibrosis (red arrowheads) and minor hypertrophy (COVID-19 patient ID 3). Myocardium of an influenza patient (Influenza patient ID 5) with minor interstitial fibrosis, moderate hypertrophy, and no inflammatory infiltrate or cardiomyocyte necrosis, whereas the myocardium in lymphocytic non-influenza myocarditis (Myocarditis patient ID 5) revealed dense infiltration of the inflammatory cells with single-cell necrosis (black arrowhead). Myocardium of a control patient (Control patient ID 8) displayed no inflammatory infiltrate, necrosis, fibrosis or hypertrophy, H&E staining, Magnification 100x, scale bar 100 µm. B Scanning electron micrographs of control heart tissue (left), COVID-19 (center), and lymphocytic myocarditis (right). In COVID-19 heart tissue (center), heart muscle fibers (myo) with slight hypertrophy are surrounded by a meshwork of collagenous fibers (black arrowheads). The orthogonal cardiac muscle orientation seems to be altered compared to the parallel organizations of myocardial strands (myo) in non-infected control heart tissue (left). The myocardial morphology in lymphocytic non-influenza myocarditis (right) is severely disturbed with pronounced edema, sporadic necrosis, and extensive lymphocytic infiltrates (yellow arrowheads), scale bars: 30 µm
A more detailed analysis of the inflammatory cell composition showed a marked increase of macrophages (CD68 +) in all infectious groups compared to controls, with influenza cases displaying the most prominent infiltrate (130.2 ± 1.0) with a twofold higher number of macrophages per mm2 compared to COVID-19 (65.2 ± 0.8) and common myocarditis (72.5 ± 1.6) (Fig. 2, supplementary table 3). Lymphocytic (CD4, CD8, CD20) infiltrates were scarce in both COVID-19 and influenza cases, whereas common myocarditis cases displayed—besides the prominent macrophage infiltrate—a marked mixed lymphocytic inflammatory infiltrate (Fig. 2, supplementary table 3).
Fig. 2
A–D MPX staining of cardiac tissue depicting CD68 + macrophages in orange, CD4 + T helper cells in green, CD8 + cytotoxic T cells in yellow, and CD20 + B-cells in magenta. All infected hearts (COVID-19, influenza, and lymphocytic non-influenza myocarditis) displayed a prominent infiltrate of CD68 + macrophages. While COVID-19 A (COVID-19 patient ID 24) and influenza, B (Influenza patient ID 9) hearts showed nearly absent lymphocytic infiltrate, lymphocytic non-influenza myocarditis, C (Myocarditis patient ID 5) was characterized by a mixed, T-cell dominated infiltrate. Non-infected control hearts, D (Control patient ID 1) showed markedly less inflammatory cells with a mixed population of macrophages and predominant t-cells and only scarce B-cells. Magnification 400x. Scale bars = 100 µm. E Histogram of the inflammatory cell infiltrates (CD20, CD4, CD68, CD8). Cell counts are normalized to cells per mm2 myocardial tissue. *p < 0.05, **p < 0.01, ***p < 0.001
CD11b + /TIE2 + monocytes/macrophages in COVID-19Macrophages were the predominant inflammatory cell type in both COVID-19 and influenza cases (Fig. 3A). Regarding macrophage subpopulations, a shift towards an M2 phenotype as indicated by the significant increase in CD16 + CD163 + macrophages in COVID-19 (141.4 ± 129.3 cells per mm2) and lymphocytic non-influenza myocarditis (1112.8 ± 1683.4 cells per mm2) when compared to influenza (32.6 ± 34.6 cells per mm2). Other marker combinations (CD16- CD163 + and CD16 + CD163 + S100A9 + , respectively) did not yield significant differences (Supplemental Table 3). We found a diffuse infiltration of CD11b + -macrophages in the perivascular connective tissue in 18 of 24 COVID-19 heart samples (Fig. 3A, B, supplementary Table 3) to a varying degree. As expected, CD11b expression was found primarily on monocytes and perivascular macrophages (Fig. 3A). As we could find a higher expression of TIE2 (TEK) in COVID-19 compared to influenza, non-influenza viral myocarditis, and control samples by nanostring analysis, we performed additional immunohistochemical stains demonstrating TIE2 expression primarily on macrophages (supplementary Fig. 1). TIE2-expressing macrophages (TieMs) were present at a significantly higher level in COVID-19 samples (0.7 ± 0.2 cells per mm2; Fig. 3C) compared to influenza samples (0.22 ± 0.108 cells per mm2; Fig. 3D) and control samples (0.27 ± 0.07 cells per mm2, Fig. 3D). Furthermore, immunohistochemical presence of TieMs was elevated more than fivefold in hearts from COVID-19 patients hospitalized later than 10 days after the detection of the infection (1.9 ± 0.6 cells per mm2, Fig. 3D) compared to COVID-19 patients hospitalized earlier (0.4 ± 0.1 cells per mm2, Fig. 3D). This is supported by an increased expression of Angiopoietin 2 in COVID-19 hearts on a gene expression level (Fig. 4) as well as on protein level, detected by IHC (supplementary Fig. 1), compared to controls.
Fig. 3
Macrophage expression of CD11b and TIE2. A Multiplex immunohistochemistry depicts a diffuse infiltration of CD11b+ macrophages (red) in the perivascular connective tissue in COVID-19 hearts (COVID-19 patient ID 17), (cardiomyocytes green, nuclei blue), scale bar 100 µm. B Bar diagram showing relative infiltration of CD11b+ inflammatory cells in non-infected control (Ctrl), influenza (Inf), COVID-19 (CoV), and lymphocytic non-influenza myocarditis (Myo) heart specimens morphometrically assessed by multiplex immunohistochemistry (MPX). Cell counts are normalized to cells per mm2 tissue. C Immunohistochemical staining against TIE2 demonstrates the perivascular localization of TIE2+ cells (red arrowheads) in the myocardium of a COVID-19 patient (COVID-19 patient ID 17), scale bar: 10 µm. D Bar diagram showing the infiltration of Tie-2+ inflammatory cells in non-infected control (Ctrl), influenza (Inf), COVID-19 (CoV), and lymphocytic non-influenza myocarditis (Myo) heart specimens. Cell counts are normalized to cells per mm2 myocardial tissue. Due to the small sample size of lymphocytic non-influenza myocarditis and a high variance among the samples, no statistical tests for significance were carried out. COVID-19 specimens were subdivided into two cohorts of cases with a hospitalization time < 10d and > 10d. *p < 0.05, **p < 0.01, ***p < 0.001
Fig. 4
Differential regulation of mRNA expression in the cardiac tissue of COVID-19, influenza, and lymphocytic non-influenza myocarditis cases assessed by multiplex NanoString nCounter analysis system. The boxplots show the expression levels of representing genes of inflammation, hypoxia, angiogenesis, monocyte recruitment, and cell adhesion. Boxplots showing normalized log2 counts of mRNA expression and standard error of the mean, whiskers indicate outliers. Significance was evaluated with Benjamini–Hochberg correction. *FDR < 0.05, **FDR < 0.01, ***FDR < 0.001
Vascular remodeling by intussusceptive angiogenesis contributes to cardiac adverse repair in COVID-19 microangiopathyUsing conventional light microscopy and immunohistochemical staining for fibrin, only scarce small vessel thrombi and no large vessel thrombi could be demonstrated in COVID-19 hearts (Fig. 1A), while two heart samples in the influenza group showed large vessel thrombi without thrombi in small vessels (Fig. 5A–C). Interestingly, contrary to the light microscopy findings, SEM imaging of corrosion casts of COVID-19 hearts showed a marked increase in the presence of ultrastructurally detectable thrombi (uTH), as indicated by abrupt breakoff of capillaries (Fig. 5D and Fig. 6A). These capillaries had a diameter of 1–3 µm, too small to be reliably detectable in conventional light microscopy, especially in autopsy material. Moreover, COVID-19 hearts showed an altered vascular architecture with a loss of vascular hierarchy, tortuous arrangement (Fig. 6A), and irregular sinusoidal vessel networks with frequent vessel diameter changes compared to the paralleled alignment of cardiac microvascularity in the healthy controls (compare Fig. 6B, D). Furthermore, the presence of transluminal intussusceptive pillars indicated by small holes in vascular corrosion casts (Fig. 5E and Fig. 6D), partly as doublets and triplets, at numerous vessel branches (Fig. 6D), could be found in COVID-19 hearts, but not in non-influenza myocarditis cases. Quantification of corrosion casts proved that non-influenza viral myocarditis cases had significantly fewer intussusceptive pillars (2.8 ± 3.7 SE) and uTh (0.1 ± 1.0 SE) compared to COVID-19 hearts (9.4 ± 1.0 SE and 2.3 ± 0.3 SE, respectively, Fig. 5F). Of note, we found a positive correlation between the presence of microthrombi and the occurrence of intussusceptive pillars in COVID-19 hearts (Fig. 5G). Corrosion casts of control hearts did not show signs of uTH or intussusceptive pillars (Fig. 6B). Archived material of the 2009 influenza pandemic suitable for corrosion casting was not available.
Fig. 5
Visualization of ultrastructurally detectable thrombi (uTh) in COVID-19 hearts. A H&E staining of a thrombus in a smaller blood vessel in a field of interstitial fibrosis. B Immunohistochemical staining against activated fibrin displayed the formation of thrombus in a larger blood vessel. C Occasionally, small intracapillary megakaryocytes were observed in COVID-19 autopsy tissue, magnification 400× (COVID-19 patient ID 9). D, E Scanning electron micrograph of microvascular corrosion casting depicting numerous irregularly dilated and blind-ending vessels with vanishing microvascular hierarchy and micro-extravasation (black arrowheads) indicative for microangiopathy in COVID-19 heart tissue. Cardiac involvement of COVID-19 demonstrates caliber changes with dilated segments and focal vasoconstrictions (red arrowheads). The expansion of vascular plexus by intussusception (yellow arrowheads) is distinctly occurring in the dilated vessel segments, preferably on sites of vessel branching. Scale bars 100 µm. F Quantification of visible microthrombi (indicated by premature obliteration of the capillary network (approximate diameter 1–3 µm)) and intussusceptive neoangiogenesis (indicated by the formation of intussusceptive pillars) in COVID-19 and lymphocytic non-influenza myocarditis compared to healthy control tissue. G Correlation between the presence of uTh formation and the number of intussusceptive pillar formation in COVID-19 and lymphocytic non-influenza myocarditis compared to non-infected control tissue
Fig. 6
Assessment of the cardiac microvasculature by microvascular corrosion casting and X-ray phase-contrast tomography. Scanning electron micrographs of microvascular corrosion casts revealed in COVID-19 hearts A a distorted microvascular architecture with dilated segments and blind-ending stocks (black arrowheads), whereas the vascular architecture of control heart tissue, B displayed a regular hierarchical organization (approximate vessel diameter 1–3 µm). C Microvascular corrosion casting exposed an irregularly shaped and tortuous microvascular architecture with numerous tiny holes, scale bar 20 µm. D Scanning electron micrograph of COVID-19 cardiac vascular plexus highlights the confluent expansion of transluminal intussusceptive pillars (black arrowheads) at a branching point, often seen as doublets and triplets, scale bar 5 µm. E Volume rendering of a tomographic reconstruction obtained by synchrotron-radiation-based X-ray phase-contrast tomography highlighting the mild interstitial fibrosis (orange) in COVID-19 heart tissue. In the magnification of the presented 2D-slice (marked by a red rectangle) a nucleus of an endothelial cell and an intraluminal pillar (black arrowheads) are visible. F Volume segmentation of microvasculature (depicted in orange) in phase-contrast synchrotron-radiation-based-X-ray tomographs demonstrate the altered microvascular architecture (Arrowheads) in COVID-19 hearts compared to the parallel alignment of coronary plexus in control hearts. The reconstructed dataset shown in F has been recorded at 167 mm voxel size. A cube of 350 µm side length is shown
Phase-contrast synchrotron radiation tomographic microscopy (SRXTM) identified the spatial coincidence of morphological aspects of disturbed, feathered course of heart fibers, mild interstitial fibrosis, and the occurrence of intussusceptive pillars (Fig. 6E). Moreover, volume segmentation of SRXTM tomographs demonstrated vascular alterations in COVID-19 hearts at the level of the afferent, large-caliber vessels (Fig. 6F), compared to regular vascular architecture in the analyzed control tissue.
Differential regulation of myocardial mRNA expression and associated biological functions in COVID-19, influenza, and myocarditisWe performed a digital multiplexed gene expression analysis using NanoString nCounter® technology. In COVID-19 hearts, we identified significant upregulation of pro-inflammatory genes IL1B, IL-6, IL8, and the toll-like receptor TLR2 (Fig. 4). Moreover, significant upregulation of hypoxia and angiogenesis-related genes as documented by VEGFC, FLT1, and NOS3 was found (Fig.4, FDR < 0.05). Gene expression analysis showed a significant upregulation of genes associated with monocyte recruitment such as CXCR4, MMP9, CCR2, CXCR2, and MYD88 (Fig. 4, FDR < 0.001). Moreover, cell adhesion markers such as ICAM1, PECAM1, and SELE were markedly, and significantly, upregulated in COVID-19, influenza, and non-influenza viral myocarditis (Fig. 4, FDR < 0.001). TIE2 was not ubiquitously upregulated in COVID-19, influenza, and non-influenza viral myocarditis compared to controls, but 10 of the 24 COVID-19 hearts displayed a marked upregulation of up to tenfold (supplementary material).
In addition to canonical pathways, differentially expressed genes were categorized to related functional pathways via gene set enrichment analysis against the Gene Ontology (GO) aspect biological functions. A complete list of GO annotations and a complete list of the analyzed genes is given in the Supplementary material. Gene expression data analysis of 797 analyzed genes showed clear differences between COVID-19 and influenza cases, but not between COVID-19 cases and non-influenza lymphocytic myocarditis (Fig. 7A). While influenza cases were primarily characterized by pro-inflammatory signaling and classical anti-viral response gene expression (e.g., BCL6, DLL4, GDF15, PTX3, HMGB2, and IL-8). COVID-19-associated gene expression showed significant upregulation of angiogenesis, cell migration, and epithelial-mesenchymal-transition (EMT) pathways in the ingenuity pathway analysis (Fig. 7B).
Fig. 7
Functional gene expression analysis of COVID-19, influenza, and lymphocytic non-influenza myocarditis heart tissues. A Functional Pathway analysis via gene set enrichment analysis against the Gene Ontology aspect biological functions highlights the differential functional gene expression in the heart tissue of COVID-19 and influenza patients. The activation of biological functions in cardiac injury patterns compared to healthy controls was predicted for each sample. Color indicates up- (red) and down (blue)-regulation; circle size depicts FDR. Only significantly up-or down-regulated pathways are shown. B Spider-Plot depicting the enrichment of biological functions from Gene Ontology based on gene expression data of COVID-19 and influenza heart samples as compared with expression in non-infected control specimen. The y-axis shows the normalized enrichment scores (NES) on a scale from 0 to 2. C Venn diagram of statistically differentially expressed genes of COVID-19 and influenza heart samples as compared with expression in controls in both disease groups (Student’s t test, controlled for the familywise error rate with a Benjamini–Hochberg false discovery rate threshold of 0.05). Up-regulation and down-regulation of genes are indicated by colored arrowheads suffixed to the gene symbols (green denotes upregulation, red denotes down-regulation). Numbers given are the total of differentially regulated genes, displayed are the top 10 up- or down-regulated genes. Note that there were no differentially expressed genes for lymphocytic non-influenza myocarditis
Comparative gene analysis demonstrated twice as much differentially expressed genes in COVID-19 compared to influenza (152 genes in COVID-19 vs 72 genes in Influenza) (Fig. 7C).
留言 (0)