
記住我
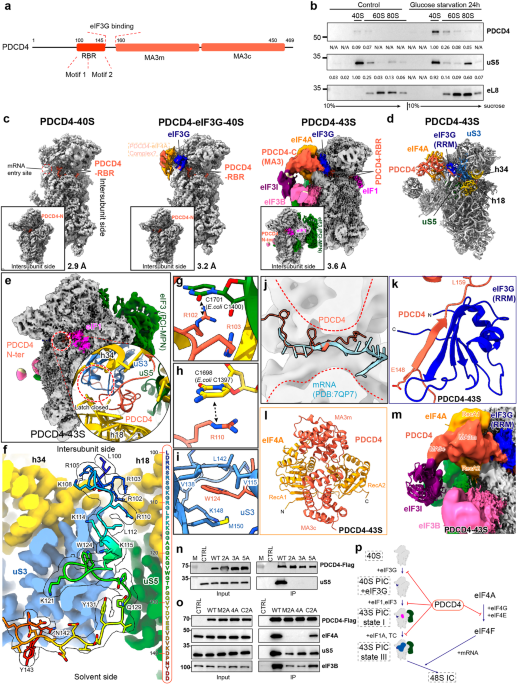

Itch, especially persistent itch (or chronic pruritus) in patients with allergic skin diseases and/or neuropathic problems, significantly affects sleep, mood and individual health. Extensive efforts have been made to develop novel therapeutic strategies to fight itch. Unfortunately, there is no FDA-approved treatment for chronic pruritus despite great efforts in antipruritic research in the past few decades.1
Bombesin (Bn) is an amphibian tetradecapeptide found in the skin of European Bombina. Two mammalian Bn-like peptides have been characterized, neuromedin B (NMB) and gastrin-releasing peptide (GRP), which are the endogenous ligands for neuromedin B receptor (NMBR or BB1 receptor) and gastrin-releasing peptide receptor (GRPR or BB2 receptor), respectively.2 In addition, there is an orphan bombesin receptor, BRS3 or BB3R, which shares sequence and structural similarity with NMBR and GRPR but is not activated by NMB or GRP (Supplementary information, Fig. S1).2 Both NMB and GRP are involved in a variety of physiological and pathological processes, including itch induced by histaminergic or nonhistaminergic pruritogens1,3,4,5,6,7 (Fig. 1a). As itch-associated neuropeptide receptors in the spinal cord, both NMBR and GRPR are at the core of itch transmission and play a pivotal role in itch biology;3 accordingly, they are attractive targets for antipruritic intervention.
Fig. 1: Cryo-EM structures of the NMB30–NMBR–Gq and GRP(14–27)–GRPR–Gq complexes and analyses of the binding and selectivity for two itch-associated neuropeptides.
a A schematic illustration of the chemical itch pathways, showing the pivotal roles of the two itch-associated neuropeptides (NMB and GRP) and receptors (NMBR and GRPR) in itch transmission. b–d Cryo-EM density map (b), model (c) of the NMB30–NMBR–Gq complex, and the dumbbell-shaped NMB30 in NMBR (d). NMB30 is shown in coral; the receptor NMBR is displayed in slate blue. e–g Cryo-EM density map (e), model (f) of the GRP(14–27)–GRPR–Gq complex, and the dumbbell-shaped GRP(14–27) in GRPR (g). GRP(14–27) is shown in orchid; the receptor GRPR is displayed in green. The heterotrimeric Gq proteins in both structures are colored by subunit: Gαq, yellow; Gβ, cornflower blue; Gγ, deep salmon; scFv16, gray. h–k Cross-section views of the NMB30-binding pocket in NMBR (h), and the GRP(14–27)-binding pocket in GRPR (j). Detailed interactions of NMB30 with NMBR (i), and GRP(14–27) with GRPR (k). Key receptor ECLs and TM helixes are annotated, while hydrogen bonds are depicted as red dashed lines. l Detailed interactions between L3N and F9N with residues in NMBR and H3G and L9G with residues in GRPR. m A comparison of detailed interactions of L3N and H3G and corresponding residues in NMBR and GRPR. n A comparison of detailed interactions of F9N and L9G and corresponding residues in NMBR and GRPR. For l–n, the related residues in the ligands and receptors are shown in stick representation. o Structural superposition of two active bombesin receptors, active CCKAR (PDB 7EZM) and inactive CCKAR (PDB 7F8U). The movement directions of TM6 and TM7 in bombesin receptors relative to inactive CCKAR are indicated by red arrows. CCKAR, cholecystokinin A receptor. Inactive CCKAR, active CCKAR, NMBR, and GRPR are colored gray, yellow, slate blue and green, respectively.
To explore the mechanism of peptide recognition by itch receptors, we assembled agonist–bombesin receptor–Gq complexes for cryo-electron microscopy (cryo-EM) studies (Supplementary information, Fig. S2) using approaches of BRIL fusion, NanoBiT tethering, Gq engineering, and antibody scFv16, which had previously been used successfully in solving several GPCR/G-protein complexes.8,9 The cryo-EM structures of NMBR and GRPR, both complexed with the engineered Gq and selective peptide agonists NMB30 and GRP(14–27), were solved at global resolutions of 3.15 Å and 3.30 Å, respectively (Fig. 1b–g; Supplementary information, Figs. S3–S5 and Table S1).
In both structures, unambiguous electron densities were observed for the C-terminal 9 residues of both bombesin agonists, which are close to the minimal 10-residue peptide required for full potency.10 Compared to the shallow, solvent-exposed pockets in the other class of itch receptors, MRGPRX2/4,11,12 both bombesin receptors exhibit classically deep pockets that are commonly found in neuropeptide receptors (Supplementary information, Fig. S6). Each bombesin agonist has a dumbbell-shaped overall conformation and adopts a binding pose perpendicular to the membrane plane (Fig. 1d, g), with its C-terminus inserted deep into the transmembrane domain (TMD) bundle and its N-terminus (referring throughout the paper to the resolved N-terminal amino acids) pointing to the extracellular surface, partially explaining the conserved activation mechanism between these two bombesin receptors. The ligand-recognition region of both receptors can be divided into two major parts: (1) the extracellular loops around the N-terminal dumbbell end and (2) the bottom of the TMD pocket burying the C-terminal dumbbell end (Fig. 1h–k). All positions of bombesin peptides are numbered from the amino terminus of NMB or NMC for clarity (Supplementary information, Table S2).
In the NMBR structure, the N-terminal dumbbell end comprises NLWAT (2–6)N (the superscript N refers to NMB30) residues, forming extensive hydrophobic interactions with residues in the binding pocket (Fig. 1i). In addition, R1032.64 forms a close hydrogen bond with T6N and plays a vital role in NMBR activation, which is consistent with our mutagenesis analysis showing that R1032.64A decreased the potency of NMB30 to activate NMBR (Supplementary information, Fig. S7a and Table S3). At the bottom of the binding pocket is the C-terminal dumbbell end formed by the HFM motif (Fig. 1i). H8N is well sandwiched by P1203.29 and P20045.52, which is further stabilized by the conserved disulfide bond between C1163.25 and C19845.50. H8N also forms hydrogen bonds with E1784.60, S1824.64, C19845.50 and I19945.51 (Fig. 1i). Alanine mutations of P1203.29, E1784.60, and P20045.52 had a great influence on receptor activation, especially the P20045.52A and P20045.52S mutations, suggesting the key role of ECL2 in peptide recognition (Supplementary information, Fig. S7b, c and Table S3). F9N forms a stabilizing π–π packing with H2836.52, while F9N and M10N fit into a hydrophobic crevice formed by the residues C962.57, V1273.36, W2796.48, H2836.52, and F3147.43 (Fig. 1i). The hydrophobic network here is essential for NMBR activation, since replacing V1273.36 with alanine entirely abolished the activity of NMB30 (Supplementary information, Fig. S7c and Table S3). The C-terminal amide forms a hydrogen bond with D1002.61, and the C-terminal carbonyl group forms an additional hydrogen bond with R3107.39 (Fig. 1i). Alanine replacements of D1002.61 and R3107.39 displayed a 10- to 50-fold reduction in NMB30 potency (Supplementary information, Fig. S7a, e and Table S3). It is noteworthy that the remarkable conformational shift of the C-terminal amidated end plays a pivotal role in NMB30 potency, since our mutation of Q1233.32 to alanine was associated with only a ~30-fold reduction in NMB30 potency, while the mutation of Q1233.32 to arginine with a bulky side chain to clash with the C-terminal amidated end completely abolished the ability of NMB30 to stimulate NMBR (Supplementary information, Fig. S7c and Table S3). Together, these detailed structural analyses provide important information to better understand the mechanism of NMB30 recognition by NMBR.
In the GRPR structure, the N-terminus adopts a more extended conformation. The side chains of R1002.64 and R2876.58 form three hydrogen bonds with the main-chain carbonyl groups of W4G, A5G, and V6G and drag the N-terminus of GRP(14–27) towards ECL1 and ECL2, where H3G forms a hydrogen bond with the backbone carbonyl of A1032.67 and an additional hydrogen bond with the side chain of S195ECL2 (Fig. 1k). In addition, W4G forms aromatic packing with H3007.31 and F3017.32 (Fig. 1k), which further stabilizes the N-terminal bending conformation. At the bottom of the binding pocket is the C-terminal dumbbell end formed by the HLM motif of GRP(14–27) (Fig. 1j, k). H8G is also sandwiched by P1173.29 and P19845.52, which are further stabilized by the conserved disulfide bond between C1133.25 and C19645.50 and the hydrogen bond between H8G and E1754.60 from TM4 (Fig. 1k). Additionally, R3087.39 forms two hydrogen bonds with H8G and L9G, while N2806.51 forms a hydrogen bond with the backbone carbonyl of L9G, and M10G forms two hydrogen bonds with D972.61 and Q1203.32 (Fig. 1k). As in the NMBR structure, the side chains of L9G and M10G fit into a hydrophobic crevice formed by the residues L892.53, C932.57, V1243.36, W2776.48, H2816.52, and F3127.43 (Fig. 1k). Alanine replacements of V1243.36, H2816.52, and F3127.43 displayed a predominant influence, showing a 3- to 6-fold reduction in GRP(14–27) potency to induce receptor activation (Supplementary information, Fig. S7f–j and Table S3). Together, these results reveal the mechanism of GRP(14–27) recognition and highlight the various peptide-binding modes between the two bombesin receptors.
The above structural insights into the binding pockets encompassing the two peptides reveal that minor amino acid differences between the two receptors reshape their binding modes and allow different peptide preferences, whereas most residues in the binding pockets of NMBR and GRPR are highly conserved (Supplementary information, Figs. S1, S8). NMB30 differs from GRP(14–27) at three residues within the C-terminal decapeptide, and the mutations of L3N and F9N in NMB30 to the equivalent H3G and L9G in GRP(14–27) are known to increase potency and specificity.10 Considering the report that NMBR binds to NMB30 with 640-fold higher affinity than GRP(14–27), whereas GRPR binds to GRP(14–27) with 650-fold higher affinity than NMB30,2 we performed structural analysis to explore the itch-associated peptide selectivity of NMBR and GRPR. The main differences are located in the ECL2 regions with a deflection of 2.1 Å, as measured between the Cα atoms of the conserved F195ECL2 in NMBR and F193ECL2 in GRPR (Fig. 1l). The swing of the side chain of F193ECL2 in GRPR away from the peptide GRP(14–27) creates sufficient space to fit the bulky side chain of H3G (Fig. 1m), which is reported to be the most important residue for receptor subtype selectivity.10 The characteristic conformation of F193ECL2 in GRPR is stabilized by the side chain of F184ECL2, which is shifted towards A19745.51. The corresponding NMBR residues are I187ECL2 and I19945.51, whose side chains are too rigid to allow similar ECL2 movement as seen in the GRPR structure. NMBR ECL2 is further stabilized by a unique hydrogen bond between T196ECL2 and E10823.49 (the cognate residue R10523.49 in GRPR pointing to the solvent). The locked conformation of F195ECL2 in NMBR greatly affects the affinity and activity of bombesin peptides with a His residue at position 3, which is consistent with a detailed study of loss- and gain-of-affinity chimeric GRPR/NMBR.13 In contrast, the structural feature of ECL2 in GRPR allows its compatibility with L3 or Q3 in its peptide ligands, which is consistent with systematic studies of 24 peptide ligands at GRPR.14 In addition, the selectivity of NMB30 for NMBR is also attributable partly to specific interactions of F9N of NMB30 with the conserved residues W6.48, H6.52, and Y6.55, which form a relatively hydrophobic binding pocket in both receptors (Fig. 1n). This NMBR pocket also contains I2165.42, which is directly packed against F9N of NMB30 (Fig. 1n). The corresponding GRPR residue is S2145.42 (Fig. 1n), which cannot form similar packing interactions with F9N of NMB30, providing a further explanation of the selectivity of NMBR for NMB30. This is consistent with previous studies with NMBR/GRPR chimeric receptors.15 In addition, T6N in NMB30 forms an intramolecular hydrogen bond interaction with L3N, which is absent in the structure of GRP(14–27) and has not been previously reported (Fig. 1l). The [V6] NMB showed a ~11-fold decrease in binding affinity to NMBR,10 suggesting that the conformation of NMB30 assisted by this unique intramolecular hydrogen bond interaction also plays a role in receptor subtype selectivity. Together, these data reveal the determinants of bombesin receptor selectivity between NMB30 and GRP(14–27).
A structural comparison of the NMBR and GRPR complexes to their closely related cholecystokinin A receptor (CCKAR) in the active (PDB 7EZM) and inactive states (PDB 7F8U) sheds light on the basis of bombesin receptor activation. Both NMBR and GRPR adopt fully active conformations similar to the active CCKAR (Fig. 1o). Bombesin receptors could be activated by bombesin peptides with C-termini in both amidated and nonamidated forms with different potencies. As described above, the C-terminal amide of the amidated NMB30 forms an additional hydrogen bond, which partially explains the phenomenon that the amidated bombesin peptides presented ~100-fold higher potencies than the nonamidated forms for their corresponding receptors (Supplementary information, Fig. S7k, l and Table S3). Superimposing the NMBR and GRPR complexes reveals that the binding modes between the two bombesin peptides are very similar, suggesting that they may activate bombesin receptors through a common mechanism (Supplementary information, Fig. S9a), although NMB30 and GRP(14–27) present different binding selectivity. The side chains of M10N and M10G insert into a conserved hydrophobic crevice at the bottom of the peptide-binding pocket and trigger the downwards rotameric movement of the toggle switch residue W6.48 in the conserved CWXP motif, facilitating the swing of F6.44 (Supplementary information, Fig. S9b). Following the repacking of an intrahelical contact between residues W6.48 and F6.44, the switching contacts of residues V3.40 towards W6.48 and L5.51 towards F6.44 contract the TM3–TM5–TM6 interface (Supplementary information, Fig. S9c), initiating the rotation of the cytoplasmic end of TM6. Overall, our study reveals the activation pathways of bombesin receptors and enriches the unique receptor-, ligand- and effector-specific activation pathways in the GPCR kingdom.
In summary, we present two cryo-EM structures of active Gq-coupled NMBR and GRPR bound to NMB30 and GRP(14–27), respectively. Our structures depict the detailed interactions between the endogenous ligands and bombesin receptors. With the in-depth knowledge of ligand binding and selectivity of bombesin receptors, new opportunities may arise to design potent and efficacious modulators of bombesin receptors for the treatment of itch and other diseases.
留言 (0)