
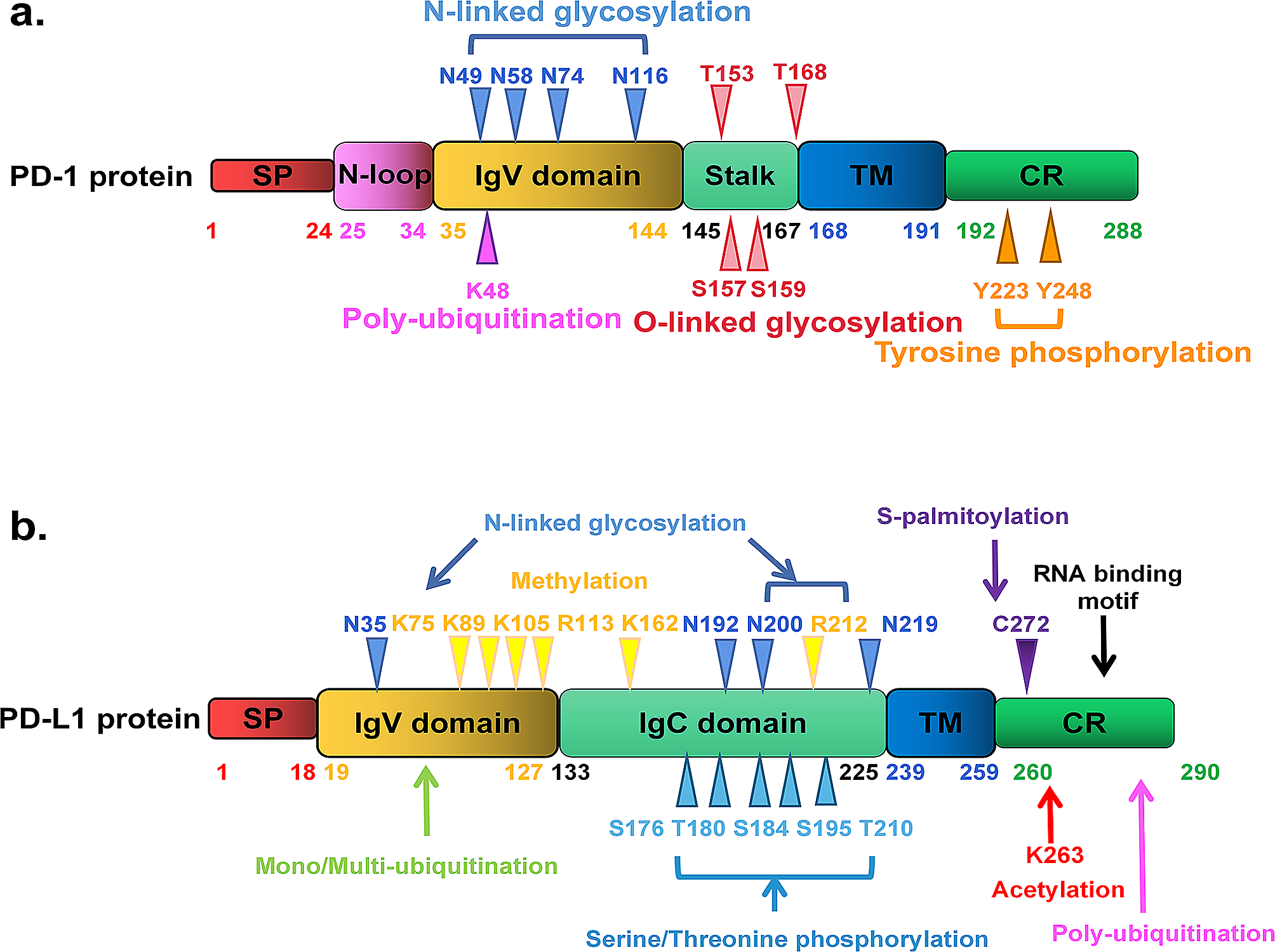
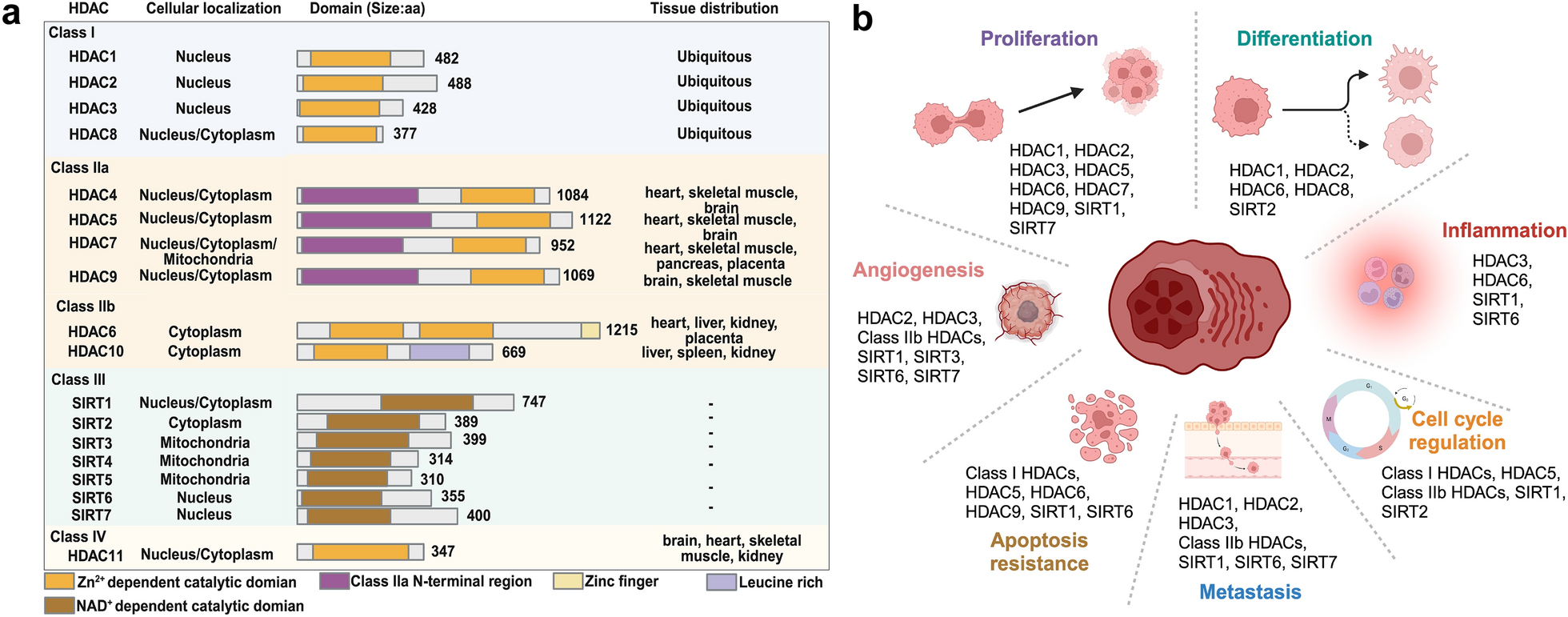
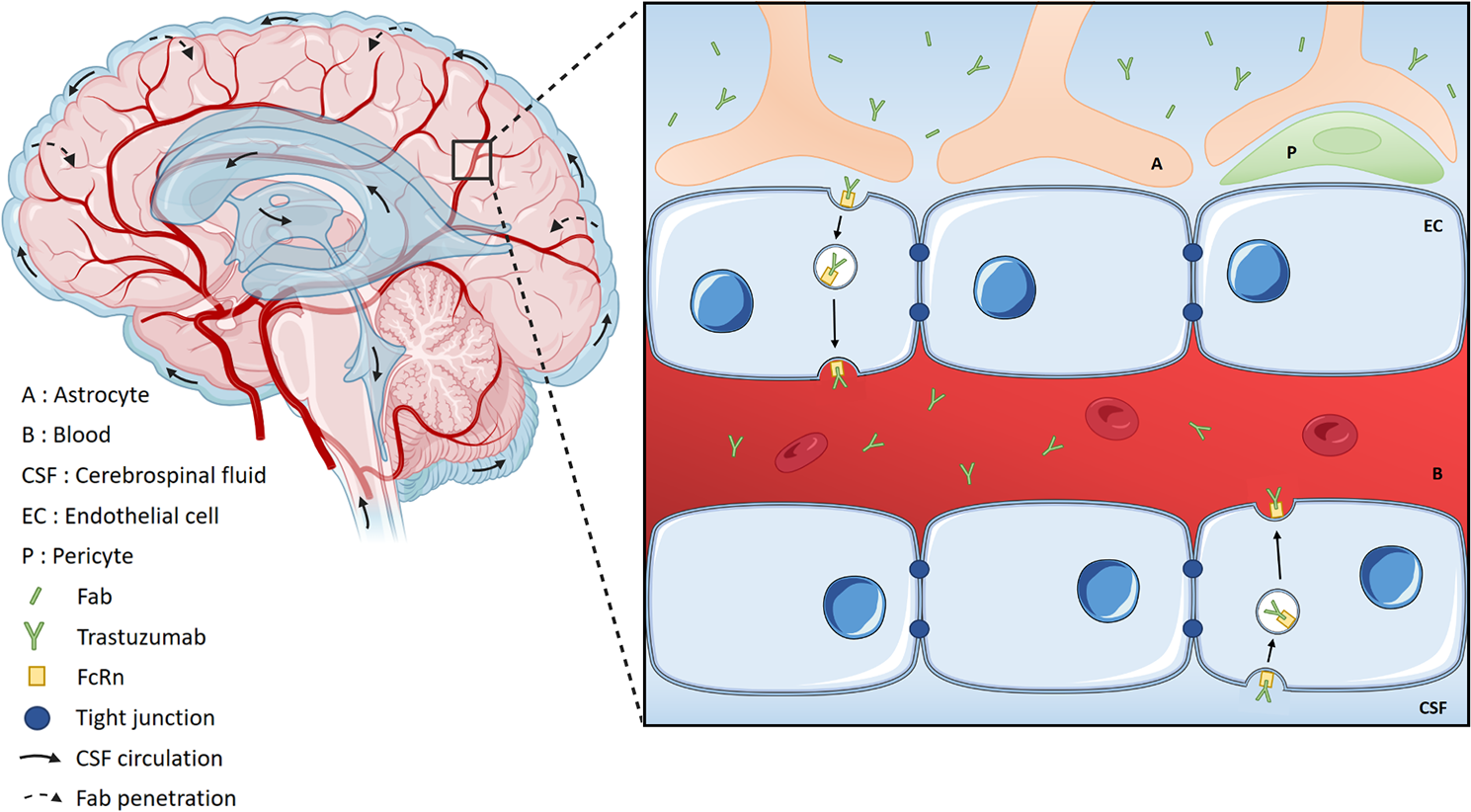
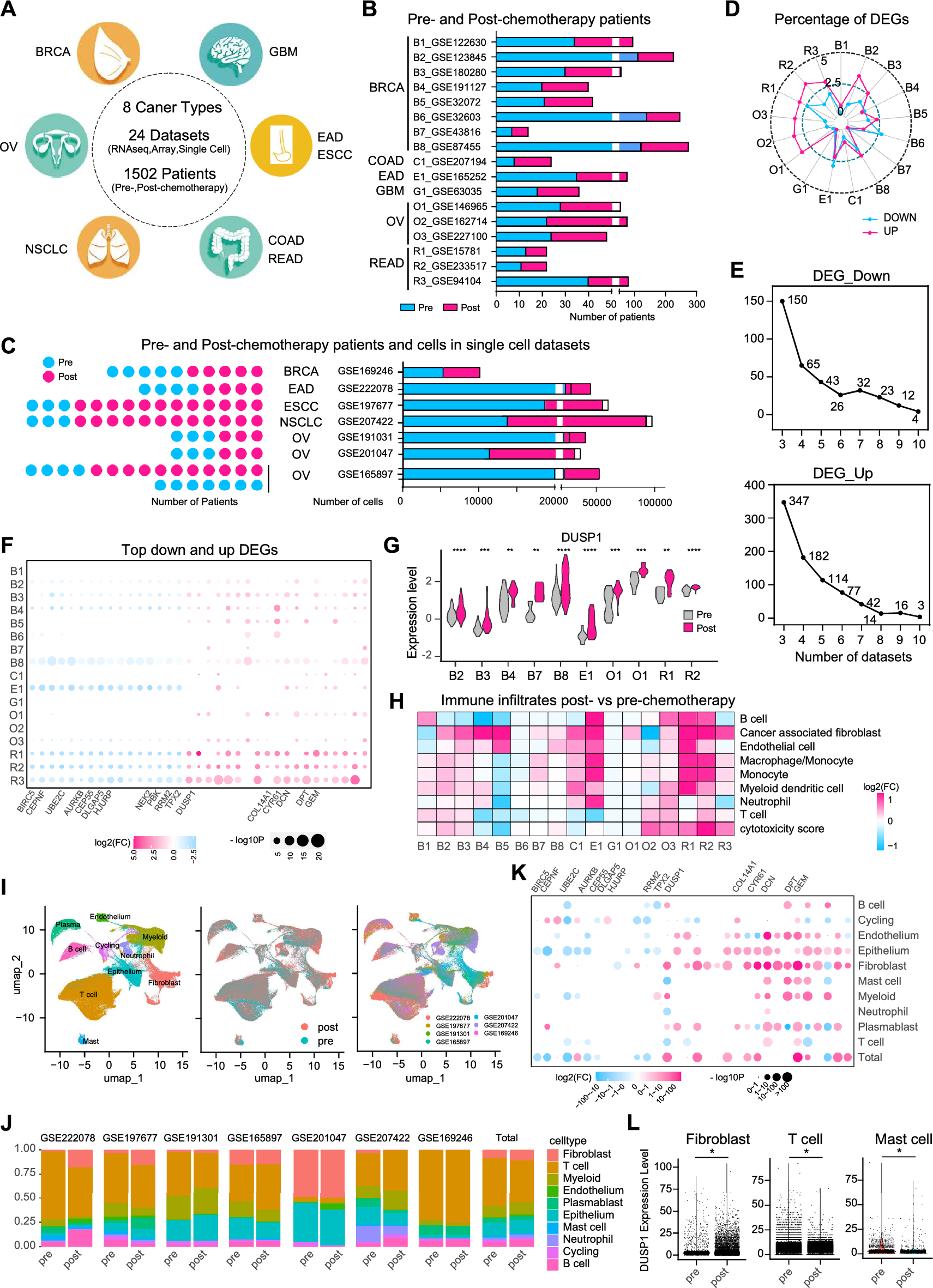
記住我
To study the involvement of IL-39 in cGVHD pathogenesis, we established a scleroderma-like cGVHD model and detected the expression of IL-39 in the lung, liver, small intestine, and spleen of recipients on days 30, 40, and 50 post transplantation. Compared with the control group (only donor bone marrow cells were transplanted), the relative expression of IL-23p19 and EBi3 was significantly increased in the intestine of cGVHD mice at 40 days post transplantation (P < 0.05), suggesting that the change in IL-39 expression may be related to cGVHD development (Additional file 1: Figure S1). To investigate the function of IL-39 in cGVHD induction, using a C57BL/6 to BALB/c cGVHD model, we injected plasmid containing Flag-tagged IL-39 gene or control plasmid into recipients 3 days before allo-HSCT using a hydrodynamic gene transfer method [13, 22]. Overexpression of IL-39 in the liver tissue was verified in mice that received the IL-39 plasmid (Fig. 1A). Serum IL-39 was also detectable up to approximately three weeks post-plasmid injection in these mice (Fig. 1B). Flag expression in the liver was confirmed by western blotting seven days after plasmid injection (Fig. 1C). To quantify the histopathology of cGVHD target organs, the skin and intestine of recipient mice were collected 56 days post transplantation. More severe tissue damage in the skin and small intestine was observed in recipients treated with IL-39 plasmid (Fig. 1D). Furthermore, we found increased collagen deposition in the skin of these mice, which was consistent with fibrosis as an important feature of cutaneous cGVHD (Fig. 1D). Although no survival difference was observed between IL-39 treated and control mice (Additional file 1: Figure S2A), treatment with IL-39 plasmid resulted in increased body weight loss and GVHD clinical scores in mice following allo-HSCT (Fig. 1F).
Fig. 1
IL-39 aggravates the development of murine cGVHD. BALB/c mice were hydrodynamically injected with minicircles or IL-39 plasmids (n = 6 per group). Three days after HGT, irradiated BALB/c recipients were infused with 1 × 107 bone marrow cells and 1 × 106 splenocytes from C57BL/6 mice, to establish a scleroderma-like cGVHD model. IL-39 expression in the liver 7 days after HGT treatment was detected by immunohistochemistry (A) and western blotting (C). IL-39 levels in the serum were detected weekly using enzyme-linked immunosorbent assay (B). Representative histopathological images and pathology scores of the skin, intestine, and Masson’s trichrome staining are shown (D). Body weights and GVHD scores in scleroderma-like mice are shown (F). Three days after HGT, irradiated recipients were infused with 5 × 106 bone marrow cells and 4 × 107 CD25− splenocytes from DBA/2 mice to establish a lupus-like cGVHD mouse model. Representative histopathological images and pathology scores of the skin, liver, intestine, kidney, and lung are shown (E). Body weights and urine protein levels in lupus-like mice are shown (G). The data are representative of at least three independent experiments. Values are presented as mean ± SD. *P < 0.05; **P < 0.01; ***P < 0.001
We further investigated the role of IL-39 in cGVHD pathogenesis using an established lupus-like cGVHD model [13]. Consistently, at 56 days after transplantation, we found more severe cGVHD in recipients treated with the IL-39 plasmid than in those treated with the minicircle control plasmid. The inflammatory response in the skin, liver, small intestine, lung, and kidney was increased, as reflected by the higher pathological scores in these organs from mice that received the IL-39 plasmid (Fig. 1E). Similar to the investigation in the scleroderma-like cGVHD model, we found no difference in survival between mice treated with IL-39 and control mice (Additional file 1: Figure S2B). The body weight of recipient mice treated with the IL-39 plasmid was markedly decreased compared to those treated with the minicircle from 5 weeks after allo-HSCT (Fig. 1G). Furthermore, higher proteinuria levels were found in recipients administered IL-39 plasmid than those in the minicircle group (Fig. 1G). Our data indicated that overexpression of IL-39 aggravates the severity of cGVHD.
IL-39 promotes donor T-cell activation and reduces the proportion of CD4+CD8+ double-positive cells in the thymus during scleroderma-like cGVHDTo further explore how IL-39 deteriorates the severity of cGVHD in recipients, we analyzed lymphocyte subsets in recipients with scleroderma-like cGVHD symptoms 8-week post transplantation. We found that the proportions of total splenic CD8+T cells and activated (CD69+) CD8+T cells in the recipients of the IL-39 group were significantly increased compared to those in the minicircle group (P < 0.01, P < 0.05). However, IL-39 had no significant effect on the proportion and number of total CD4+ T cells or activated CD4+T cells (Fig. 2A). We further detected intracellular cytokines and found that IL-39 overexpression upregulated the percentage of TNF-α-positive CD4+ and CD8+T cells in mice after allo-HSCT (Fig. 2B). IL-4 expression in CD4+T cells was slightly increased by the IL-39 plasmid treatment, but the difference was not statistically significant (Fig. 2C). These results suggest that IL-39 not only increases the percentage of CD8+T cells and promotes their activation, but also increases the proportion of TNF-α-producing T cells after allo-HSCT.
Fig. 2
IL-39 promotes pro-inflammatory responses of T and B cells in the scleroderma-like cGVHD mice model. Irradiated BALB/c recipients were infused with 1 × 107 bone marrow cells and 1 × 106 splenocytes from the C57BL/6 mice. Splenocytes (n = 6 in each group) were collected and stained for FACS analysis eight weeks after transplantation. The percentages and numbers of CD4+, CD69+CD4+T, CD8+, and CD69+CD8+T cells in H2Kb+ cells from the spleens of recipients are shown (A). Lymphocytes were isolated from the spleens of recipients and treated with PMA, brefeldin A, and ionomycin for 4-6 h. The percentage and number of TNF-α-(B) and IL-4-(C) positive T cells in the spleens of recipients are shown. The percentage and number of Treg (CD4+CD25+Foxp3+) cells in the spleens of recipients are shown (D). The percentage and number of CD4+CD8+T cells and the CD4+/CD4+CD8+ and CD8+/CD4+CD8+ ratios in the thymus of recipients are shown (E). The percentage and number of Tfh (CD4+CXCR5+PD-1+) cells in the spleens of recipients are shown (F). The percentage and number of GC B (B220+GL-7+Fas+) cells in the spleens of recipients are shown (G). The data are representative of at least three independent experiments. Values are presented as mean ± SD. *P < 0.05; **P < 0.01
Treg cells maintain immune tolerance and prevent autoimmune and inflammatory diseases, including cGVHD [23,24,25]. We found that the percentage of Tregs was slightly reduced, whereas the absolute numbers were markedly decreased after IL-39 plasmid treatment (Fig. 2D). Previous studies have demonstrated that donor CD8+T cells impaired the negative selection process during T cell development in the thymus [9]. Based on our results that IL-39 can promote the activation of CD8+T cells, we speculated that IL-39 might aggravate thymic injury. In line with this hypothesis, the frequencies and absolute numbers of CD4+CD8+ thymocytes were significantly reduced in mice treated with the IL-39 plasmid compared to those in the control group (Fig. 2E). The ratio of single positive (SP) to double positive (DP) thymocytes was also markedly increased in mice administered the IL-39 plasmid (Fig. 2E). Our data indicated that injection of the IL-39 plasmid reduced Treg numbers and might aggravate thymic damage during scleroderma-like cGVHD development, suggesting its pathogenic role in the breakdown of immune tolerance (Fig. 2E).
IL-39 promotes the differentiation of GC B cells in the spleens of scleroderma-like cGVHD recipientsTo confirm whether IL-39 affects B-cell response during cGVHD, we harvested splenocytes from recipients with scleroderma-like cGVHD and examined the expression of surface markers of B lymphocytes, including B220, Fas (CD95), GL-7, and CD138 in recipient mice on day 56 post allo-HSCT. Analysis of B cell subsets indicated that the ratio, but not the absolute number of GC B cells (GL7+CD95+), was significantly increased in recipients with IL-39 plasmid administration (Fig. 2F). T follicular helper (Tfh) cells also play a vital role in the pathogenesis of cGVHD by supporting B-cell activation and promoting GC B-cell differentiation [5, 12]. However, the percentage and absolute number of Tfh (CD4+CXCR5+PD-1+) cells were comparable between the two groups (Fig. 2G). These data suggest that IL-39 promotes germinal center responses by enhancing GC B cell differentiation during cGVHD development.
IL-39 increases T- and B-cell responses during the development of lupus-like cGVHDTo exclude the possibility of model specificity, we established a lupus-like cGVHD model, and explored the mechanisms underlying IL-39 pathogenicity in cGVHD. As we could not distinguish between donor and recipient cells in this MHC-matched model, we analyzed whole T and B cells in the recipient spleens. We found that the percentages of activated CD4+T cells and CD8+T cells were increased in the spleens of recipients who received the IL-39 plasmid compared to those receiving the minicircle (Additional file 1: Fig. S3A). Consistent with the findings in the scleroderma-like cGVHD model, we found that the frequencies of TNF-ɑ-and IL-4-expressing CD4+T cells were significantly increased in the spleens of lupus-like recipients with IL-39 plasmid treatment (Additional file 1: Fig. S3B and S3C). Although not statistically significant, there was a small increase in the proportion of TNF-α-producing CD8+T cells (Additional file 1: Fig. S3B). However, IL-39 plasmid treatment in this cGVHD model did not affect the percentage or absolute number of Treg cells (Additional file 1: Fig. S3D). We found that the frequency of CD4+CD8+ thymocytes was significantly decreased, while the ratios of CD4+/DP and CD8+/DP were significantly elevated in IL-39 plasmid-treated recipients (Fig. 3A), indicating that IL-39 could exacerbate thymic damage in both lupus-like and scleroderma-like cGVHD models. In addition, we did not observe a significant difference in the percentage or absolute number of Tfh cells in the lupus-like model (Fig. 3B), whereas the frequency and number of splenic GC B cells increased in IL-39-overexpressing mice (Fig. 3C). Taken together, our data indicate that IL-39 infusion enhances T cell activation and GC B cell differentiation, while reducing the proportion of CD4+CD8+ double-positive cells in the thymus, thereby aggravating lupus-like cGVHD.
Fig. 3
IL-39 promotes the pro-inflammatory responses of T and B cells in the lupus-like cGVHD mice model. Irradiated BALB/c recipients were infused with 5 × 106 bone marrow cells and 4 × 107 CD25− splenocytes from DBA/2 mice. Splenocytes (n = 6 per group) were collected and stained for FACS analysis 8 weeks post transplantation. The percentage and number of CD4+CD8+T cells and the CD4+/CD4+CD8+ and CD8+/CD4+CD8+ ratios in the thymuses of recipient mice are shown (A). The percentage and number of Tfh (CD4+CXCR5+PD-1+) cells in the spleens of recipients are shown (B). The percentage and number of GC B (B220+GL-7+Fas+) cells in the spleens of recipients are shown (C). The data are representative of at least three independent experiments. Values are presented as mean ± SD. *P < 0.05; **P < 0.01
Blocking IL-39 ameliorates the immune dysregulation during scleroderma-like cGVHDGiven that increased IL-39 levels exacerbated pro-inflammatory immune responses during cGVHD, we investigated whether blocking IL-39 could ameliorate immune dysregulation in the scleroderma-like cGVHD model without artificial IL-39 overexpression. We first examined the effectiveness of the anti-IL-39 antibody in vitro and found that anti-IL-39 antibody suppressed the secretion of pro-inflammatory cytokines in T cells (Additional file 1: Figure S4). We next used a scleroderma-like cGVHD model to investigate the in vivo effect of anti-IL-39 on pro-inflammatory immune responses. We observed that no mice died in either group during the entire experimental period, except for one that died before antibody infusion (Additional file 1: Figure S5A). Body weight was significantly higher in IL-39 antibody-treated mice than in control mice (P < 0.01), and there was also a significant decrease in the GVHD clinical score in IL-39 antibody-treated mice compared with control mice on day 42 (Additional file 1: Figure S5B, P < 0.05). We found that blocking IL-39 significantly decreased the frequencies of CD8+T and CD69+CD8+T cells and reduced the numbers of CD4+T and CD69+CD4+T cells in recipient spleens compared to those treated with the isotype control antibody (Fig. 4A). There was a slight decrease in the absolute number of CD8+T cells in the spleens of anti-IL-39 antibody-treated mice (P = 0.058) (Fig. 4A). Furthermore, blocking IL-39 significantly attenuated the percentage of TNF-α-or IL-4-producing CD4+T cells and reduced the proportion and absolute number of TNF-α-producing CD8+T cells (Fig. 4B, C). Accordingly, the immune regulatory function of IL-39 inhibition can be investigated in target tissues. TNF-α expression in CD8+T cells and IL-4 expression in CD4+T cells were downregulated in the intestine (Additional file 1: Figure S6A). The numbers of CD8+T cells and CD69+CD4+T cells decreased in the liver (Additional file 1: Figure S6B, P < 0.05). The number and percentage of CD8+T cells decreased in the lungs (Additional file 1: Figure S6C, P < 0.05). There was no difference in the proportion or number of Tregs between the anti-IL-39 antibody and isotype control antibody treatment groups (Fig. 4D). Anti-IL-39 treatment did not affect the frequency of CD4+CD8+ thymocytes or the SP/DP ratio (Fig. 4E). Although the percentage and number of Tfh cells were not affected by the IL-39 blockade (Fig. 4F), the proportion of splenic GC B cells was significantly reduced in anti-IL-39-treated mice (Fig. 4G). Taken together, these data suggest that the administration of anti-IL-39 antibodies might alleviate immune dysregulation during cGVHD by controlling T cell expansion and activation, suppressing pro-inflammatory cytokine production of donor T cells, and attenuating GC responses.
Fig. 4
Anti-IL-39 antibody ameliorates immune dysregulation in scleroderma-like cGVHD. Irradiated BALB/c recipients were infused with 1 × 107 bone marrow cells and 1 × 106 splenocytes from the C57BL/6 mice. Fourteen days after transplantation, each mouse in the antibody group received 100 μl (100 μg) of IL-39 antibody, while each mouse in the control group received 100 μl (100 μg) of isotype control antibody via intraperitoneal injection twice a week for 6 weeks. Splenocytes (n = 4 in each group) were collected and stained for FACS analysis eight weeks after transplantation. The percentages and numbers of CD4+, CD69+CD4+T, CD8+, and CD69+CD8+T cells in H2Kb+ cells from the spleens of recipients are shown (A). Lymphocytes were isolated from the spleen and treated with PMA, brefeldin A, and ionomycin for 4-6 h. The percentage and number of TNF-α-(B) and IL-4-(C) positive T cells in the spleens of recipients are shown. The percentage and number of Treg (CD4+CD25+Foxp3+) cells in the spleens of recipients are shown (D). The percentage and number of CD4+CD8+T cells and the CD4+/CD4+CD8+ and CD8+/CD4+CD8+ ratios in the thymuses of recipients are shown (E). The percentage and number of Tfh (CD4+CXCR5+PD-1+) cells in the spleens of recipients are shown (F). The percentage and number of GC B (B220+GL-7+Fas+) cells in the spleens of recipients are shown (G). The data are representative of at least three independent experiments. Values are presented as mean ± SD. *P < 0.05; **P < 0.01
IL-39 is positively correlated with the severity of cGVHD in patientsTo explore the effect of IL-39 on clinical cGVHD, we performed a retrospective analysis of patients who developed cGVHD after allo-HSCT and those who did not. According to the NIH consensus criteria, we grouped the patients into no cGVHD, mild cGVHD, and moderate/severe (M/S) cGVHD groups. In addition, CXCL13 has been previously suggested as an essential indicator of the occurrence and development of cGVHD [26, 27]. To further describe the disease status of patients, we detected serum CXCL13 levels, and found that the average concentration of CXCL13 was 113.5 pg/ml, 301.0 pg/ml, and 368.6 pg/ml in patients with no cGVHD, mild cGVHD, and moderate/severe (M/S) cGVHD group, respectively (Additional file 1: Table S4). We then measured IL-39 levels in the corresponding patient groups. We found that IL-39 concentrations were higher in patients with cGVHD than in those without (Fig. 5A). Furthermore, IL-39 levels showed an increasing trend in patients with M/S cGVHD compared to those with mild cGVHD (Fig. 5A). When comparing patients with or without M/S cGVHD, the AUC of CXCL13 and IL-39 were 0.686 (95% CI 0.528–0.844, P = 0.032) and 0.717 (95% CI 0.569–0.865, P = 0.013), respectively. The CXCL13 cut-off value was 125.78 pg/ml from the ROC curve, and the sensitivity and specificity were 87.9% and 47.1%, respectively. The IL-39 cut-off value was 52.44 pg/ml, the sensitivity and specificity were 69.7% and 64.7%, respectively (Fig. 5B). Therefore, our data indicate a positive correlation between IL-39 levels and cGVHD development, suggesting that high IL-39 levels may be a predictor of M/S cGVHD development in patients post allo-HSCT.
Fig. 5
Serum IL-39 levels were significantly elevated in patients with cGVHD. CXCL13 and IL-39 levels in serum were detected in non-GVHD, mild, and moderate/severe cGVHD patients (A). The prognostic value of CXCL13 and IL-39 in no-cGVHD and mild and moderate/severe cGVHD patients was evaluated using ROC curves (B). CD4+T cells, CD8+T cells, B cells, and CD11b+ cells were sorted from the splenocytes of C57BL/6 mice or PBMCs from healthy donors using a Cell Isolation Kit according to the manufacturer’s protocol. For CD4+ and CD8+T cell activation, plates were coated with 2 mg/ml anti-CD3 and 0.4 mg/ml anti-CD28 Abs overnight. Next, 2 × 105 T cells were cultured with recombinant mouse or human IL-39 proteins (20 ng/ml) for 72 h. For B cell and CD11b+ cell activation, 2 × 105 B cells and CD11b+ cells were cultured with LPS (5 ng/ml) and recombinant mouse or human IL-39 proteins (20 ng/ml) for 72 h. The relative expression of IL-39 and IL-39R genes was determined by real-time PCR in CD4+T cells, CD8+T cells, B cells, and CD11b+ cells from the splenocytes of C57BL/6 mice and PBMCs from healthy donors (C). In the following experiments, plates were coated with 2 mg/ml anti-CD3 and 0.4 mg/ml anti-CD28 Abs overnight. T cells (2 × 105 T cells were cultured with various concentrations of recombinant mouse or human IL-39 proteins for 72 h. Phosphorylation of STAT1/STAT3 detected by western blotting in sorted primary T cells from mice or healthy donors is shown (D, E). The data are representative of at least three independent experiments. Values are presented as mean ± SEM. *P < 0.05; ***P < 0.001
IL-39 affects the expression of itself and its receptor on immune cells and exerts a pro-inflammatory response in T cells via STAT1/STAT3 signaling pathwayWe further investigated potential downstream pathways in immune cells that respond to IL-39 stimulation. Currently, there are no specific antibodies against the IL-39 receptor. Therefore, we examined IL-39 or IL-39 receptor-related gene expression in sorted CD19+B cells, CD4+T cells, CD8+T cells, and CD11b+cells in mice and humans. We examined IL-39 heterodimers, including IL-23p19 and EBi3, as well as IL-39 receptor heterodimers, including IL-23R and gp130. Surprisingly, we found that although LPS (5 ng/ml) activated B cells, CD11b+ cells, and anti-CD3/CD28 stimulated CD4+T cells produced moderate amounts of genes encoding heterodimers of IL-39 and its receptor, both mouse- and human-derived activated CD8+T cells expressed high levels of EBi3, IL-23p19, IL-23R, and gp130 (Fig. 5C). Interestingly, the addition of IL-39 only slightly affected the expression of genes encoding IL-39 and its receptor in CD8+T cells, while significantly promoting the expression of IL-23R and gp130 in CD4+T cells (Fig. 5C). Moreover, IL-39 administration promoted the expression of IL-23p19 in B cells as well as the expression of both EBi3 and IL-23p19 in CD11b+ cells (Fig. 5C). Taken together, these findings suggested that activated CD8+T cells might produce massive amounts of IL-39 and express high levels of IL-39 receptor, regardless of whether they were IL-39 treated or not, whereas IL-39 administration mainly increased the expression of IL-39 receptor on CD4+T cells and promoted the production of IL-39 on B cells and CD11b+cells.
We next investigated whether IL-39 exerted its pro-inflammatory effects on T cells by activating the downstream STAT pathways. We found that the phosphorylation of STAT3 was significantly increased in sorted murine primary T cells co-cultured with 50 ng/ml mouse recombinant IL-39 protein (Fig. 5D). However, we did not observe the activation of the STAT1 pathway (Additional file 1: Figure S7). We also isolated T cells from peripheral blood mononuclear cells (PBMCs) of healthy donors and co-cultured them with recombinant human IL-39 protein. Interestingly, we found that rIL-39 significantly augmented the phosphorylation of STAT1 rather than STAT3 in human T cells (Fig. 5E). These data suggest that IL-39 may mediate inflammatory responses through activation of STAT3 in murine primary T cells and STAT1 in human T cells.
留言 (0)