
記住我




While preparing to give a lecture to medical students, Frederick Banting read an article by Dr. Moses Barron reporting autopsy findings demonstrating that pancreatic ductal lithiasis resulted in the formation of new islets surrounding the ductal obstruction. Dr. Banting scribbled down his initial study idea of tying off the ducts within pancreas in lab animals and collecting the subsequent pancreatic fluids for the treatment of diabetes and these pancreatic fluids became known as insulin.1-3 The regenerative powers of the pancreas were well described in the early 1900s by DeTakats and others who performed surgery to partially ligate the pancreas among children with diabetes.4, 5 The improvement in diabetes following the procedure was only transient because it was not known at the time that type 1 diabetes is an autoimmune disease requiring not only regeneration of new islets producing six hormones, including insulin, but also immune suppression to prevent ongoing autoimmune beta cell attack.4, 5
Over the past 30 years, researchers have conducted similar studies to Banting and have consistently found that destruction of the pancreas results in the formation of new islets. Using genomic and proteomic methods, Reg gene proteins are upregulated not only in animal models of pancreatic injury but also present in human diabetes. Initially thought to be part of the destructive and inflammatory process, Reg gene proteins and shorter bioactive regions that are within the Reg protein have been shown to initiate the formation of not only beta cells but entire islets.
Reg has been shown to initiate islet formation in utero when the pancreas is initially formed and expressed again only during times of pancreatic injury including type 1 and type 2 diabetes, pancreatitis, and pancreatic stones. Terazono and colleagues first described the Reg gene in 1988, when screening the DNA library and identified a novel gene encoding a 165-amino acid protein that was expressed in regenerating islets in 90% pancreatectomized rates but not in normal pancreatic islets.6
Terazono found the increase in expression of Reg was correlated with an increase in size of regenerating islets and the decrease in urinary glucose level. The receptor of rat Reg gene, which is homologous to the human exostosin-like glycosyltransferase 3 (EXTL3) that was later described by Kim and colleagues in 2001,8 was identified by Kobayashi in 2000.7
There are four Reg proteins among more than 30 mammals including humans, with subgroups based on their protein sequences in humans and mammals. All four human Reg proteins are found in mammals have areas of homologous sequences and there is a 70% or greater homology between the Reg proteins.
The role of Reg proteins and shorter bioactive Reg peptides within one's own pancreas in transforming pancreatic ductal cells into new islets has been shown by teams around the world, as well as having specific antiapoptotic and anti-inflammatory properties in islets.9-31 Reg gene proteins in more than 30 mammals and in humans are grouped into four categories by structure. All have homologous peptide sequences by 70% or more. Reg is also referred to as PAP (pancreatitis associated protein), PSP (pancreatic stone protein), and PTP (pancreatic thread protein) formerly called lithostathine.
Reg gene proteins found in embryogenesis are pivotal in the formation of pancreatic islets and then not expressed after fetal development, except during injury and during pregnancy, when there is a need for increased islet formation.32-35 Reg proteins were initially found at the site of pancreatic injury including new onset type 1 diabetes and considered to be deleterious and part of the inflammatory process. Over the past few decades, Reg proteins have been found to be upregulated to regenerate new cells and of key importance in the generation of new islets following pancreatic injury.36
Bonner-Weir and others have found that cells can be identified within ductal tissue that are specifically triggered to form new islets containing alpha, beta, gamma, delta, epsilon, and pancreatic polypeptide cells and that pancreatic exocrine ductal cells represent an endogenous pool for new islet production.37-40 Shorter Reg peptides that are homologous within 30 mammalian species have been also shown to transform pancreatic ductal tissue into new islets.25, 29, 30
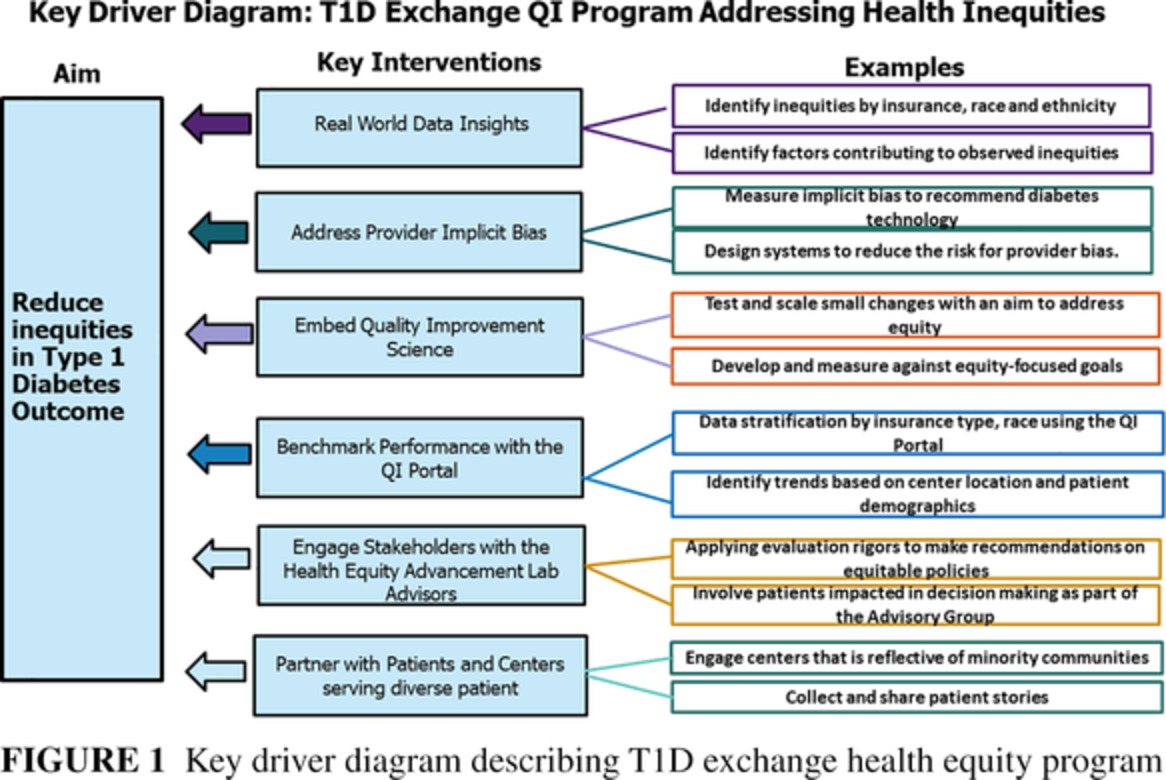
2 ISLET NEOGENESIS VERSUS BETA CELL REPLICATIONEven with more than 60 medications approved and a dozen different insulin preparations, the loss of functional islets is not addressed by any single or combination of therapeutics. Despite all the diabetes therapies and technological advances, the worldwide cost of diabetes is nearly $1 trillion ($850 billion) USD per year, and worldwide, diabetes is the number one cause of amputations, kidney disease requiring dialysis or transplant, and new blindness.
Although diabetes is a disease of loss of beta cells either from autoimmune attack or more chronic destruction, postmortem diabetes evaluations conducted both among type 1 and type 2 diabetes demonstrate that there is not only loss in the quantity of insulin-producing beta cells but also reduction in the number of islets containing alpha, beta, gamma, delta, and epsilon cells that generate islet ghrelin.41, 42
Human physiology demonstrates why insulin alone cannot restore entire islet function and why it is so difficult to achieve normoglycemia without new functioning islets. For example, 80% of all recorded sensor glucose levels among those without diabetes are below 100 mg/dL, and 95% of all glucose levels after meals are less than 121 mg/dL.43
Questions are also raised as to what is the hemoglobin A1C (A1C) for which there are no increased microvascular and macrovascular risks associated with diabetes. Three prospective studies have found identical A1C levels of 5.5%, above which there are increased risk microvascular complications in type 1 and 2 diabetes and macrovascular-related morbidity and mortality.44-46
As shown in Figure 1, beta cells are just one component of human islets, with some studies demonstrating 70% or more of human beta cells are in direct contiguity with other cell types, indicating that the importance of intracellular communication between cell types is necessary for glucose homeostasis in humans in contrast to mice.47 Islets consume 20% of the blood flow to the pancreas yet are only 2% of the mass. This unique vascularity within islets and the islet architecture that is unique to humans, help understand why human beta cells must exist within islets and why islets must exist within the pancreas.

This photomicrograph of a human islet demonstrates the beta cells staining for insulin in red, the alpha cells staining for glucagon in green with blue-stained cells indicating somatostatin. The large zones uniquely seen within the human islet are blood vessels lined by smooth muscles innervated that are innervated by the nervous system. Reproduced from Proceedings of the National Academy of Sciences Feb 2006, 103 (7) 2334–2339; DOI: 10.1073/pnas.0510790103. Copyright 2006 National Academy of Sciences. Cabrera O, Berman DM, Kenyon NS et al., The unique cytoarchitecture of human pancreatic islets has implications for islet cell function (Reference 48)
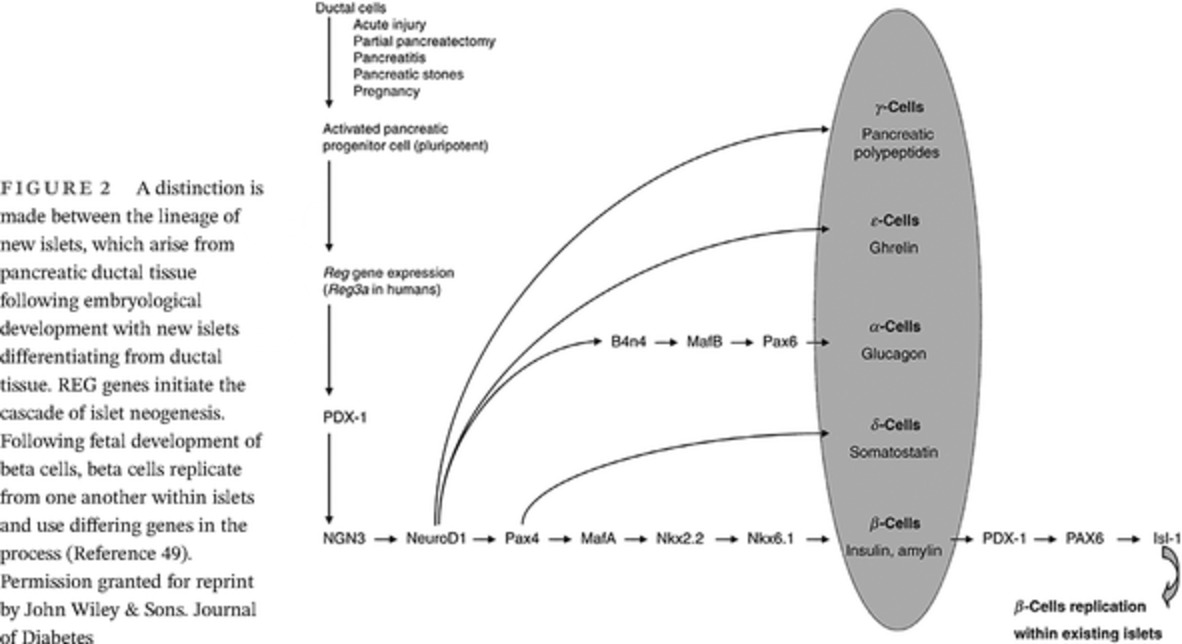
The importance of the distinctions between islets and beta cells and islet neogenesis and beta cell regeneration is that Reg proteins have been shown to begin the cascade of islet neogenesis from progenitor cells, which are found in the pancreatic exocrine cells.48 As shown in Figure 2, beta cell replication from existing beta cells involves transcription factors and occurs independently of Reg.

A distinction is made between the lineage of new islets, which arise from pancreatic ductal tissue following embryological development with new islets differentiating from ductal tissue. REG genes initiate the cascade of islet neogenesis. Following fetal development of beta cells, beta cells replicate from one another within islets and use differing genes in the process (Reference 49). Permission granted for reprint by John Wiley & Sons. Journal of Diabetes
3 THE ROLE OF REG IN REGENERATIONWhen Reg 3a was found at the site of inflammation in the pancreas during new onset type 1 diabetes it was thought to be part of beta cell destruction.36 Over the past several decades, numerous studies from teams around the world have shown that Reg proteins are expressed during acute pancreatic injury to regenerate new islets from pancreatic ductal cells found predominantly in the pancreatic exocrine cells. More recently, the anti-inflammatory actions of Reg proteins are being further elucidated.31
The expression of Reg occurs during embryogenesis when the pancreatic islets are formed for the first time and then expressed again in times of pancreatic injury, including pancreatic stones and during pregnancy.9-30, 32-35 Teams from around the world have demonstrated both in vivo and in vitro that shorter Reg peptides result in new islet formation from pancreatic ductal tissue. For example, Kapur demonstrated in vivo how administration of shorter Reg peptides (i.e., 14- and 15-amino acids peptides) induces up to a significant doubling in new islet formation from pancreatic ductal tissue and significant increases in islet markers Ins, Sox9, Ngn3, and Nkx6.1.25 Shorter Reg gene peptides have been shown to upregulate islet transcription factors: Sox9, Nkx6.1, Nkx2.2, Pax6, MafB, MafA, B4n4, NGN3, NeurodD1 and Pax4, PDX-1, NGN3, and NeurodD.25
From the Joslin Laboratory, a 15-amino shorter Reg peptide was identified in the newest islets that were budding directly from pancreatic ductal cells with high concentration of the 15-amino acid Reg peptide and diminishing as the new smaller islets became larger.49 Figure 3 demonstrates the in vitro findings of Reg peptides accelerating islet production from human ductal tissue.30

The left in vitro culture is from human ductal tissue in culture after 6 days and on the right are new islets after 6 days in culture with a 14-amino acid Reg peptide. The islets on the right stained positive for insulin, glucagon, and somatostatin. Image provided by Shaoping Deng MD, PhD, Director, Human Islet Laboratory, University of Pennsylvania (personal communication)
Data illustrates that the Reg gene proteins are responsible for triggering the cascade of forming new islets. There are distinct regeneration factors necessary for beta cell formation from existing beta cells.
Studies have shown the role of and the requirement of Reg new islet formation in aging and high-fat diet induced obesity and the role of Reg in restoring glucose homeostasis in obesity in animal models.50, 51 Studies have shown the reversal of diabetes with Reg proteins and shorter peptides in preclinical mammalian studies.52, 53
4 ROLE OF REG IN PREGNANCYPregnancy is another condition in which Reg genes are upregulated, which has been shown in preclinical studies and in expression libraries among pregnant and nonpregnant mice and rats.32-35 The ability to form new islets, even among pregnant type 1 patients with long-standing disease has been shown by the unique work of Dr. Lois Jovanovic, whose team demonstrated how consecutive pregnant patients with type 1 diabetes for an average of 21 years had normalization of endogenous insulin, as measured by C-peptide within 10 weeks of pregnancy that rose from undetectable levels before pregnancy (Figure 4).54

Data from Reference 56 demonstrate 10 consecutive patients from Dr. Lois Jovanovic's type 1 pregnancy clinic who had a duration of diabetes for more than 21 years. Before pregnancy, there were no detectable levels of endogenous insulin as measured by C-peptide, with normal fasting levels by 10 weeks of pregnancy
During normal pregnancy, there is a 200% to 250% increase in insulin secretion to maintain euglycemia in the mother, whereas there is a concomitant rise in Reg gene expression.55 There are also unique and complex immune changes during pregnancy that protect the mother from attacking her fetus as foreign, which may contribute to the lack of autoimmune attack of beta cells.
5 REG AS A DIABETES THERAPEUTICReg peptide studies have been translated into potential therapeutics as shown in human trials.56, 57 Human Reg 3 alpha and hamster-derived Reg 3 gamma have both been used in clinical trials. Reg peptides are an integral part of a pathway of protein-protein interaction. Because there are numerous inhibitory feedback loops, including hypoglycemia, which inhibits new islet formation, unregulated cell growth has not been seen in the Reg peptide trials.
Studies with short Reg peptides have been conducted in both type 1 and 2 diabetes patients through phase 2B with significant findings of reduction in A1C among type 2 patients and a significant rise of 30% in C-peptide area under the curve among type 1 patients with a mean history of the disease of 21 years (p = 0.0057).56 Reg peptides have demonstrated the potential to improve diabetes among type 2 patients, including a significant reduction in A1C that was twice that of placebo-controls (p = 0.009).57
Shorter peptide sequences homologous in 30 mammalian species, including humans, have been shown to bind to the Reg receptor resulting in the downstream formation of new islets, containing new populations of beta cells along with four other hormone-secreting cell types that contribute to glucose homeostasis.30 These shorter Reg peptides may be the first class of diabetes therapeutics to address the underlying causes of diabetes without the need for islet or stem cell transplants by using the pancreatic ductal cells that already comprise 98% of one's pancreas. The challenges remain to protect beta cells from immune destruction as seen in type 1 diabetes or in both type 1 and 2 diabetes are to maintain beta cells formed within new islets from destruction, whether that be autoimmune attack in the case of type 1 diabetes or metabolic factors contributing to beta cell loss in type 2 diabetes.
6 SUMMARYResearch teams from around the world have found that Reg gene proteins are upregulated by conducting pancreatic injury studies similar to the one conducted by Sir Frederick Banting, which led him to the discovery of insulin. Shorter bioactive regions of the protein have the potential as therapeutics to induce the formation of new islets from one's own pancreas, even among those living with diabetes for decades because Reg transforms pancreatic exocrine ductal tissue into new islets. The formation of new islets is truly the key to prevention of mortality and morbidity of diabetes. Oral peptide and weekly peptide therapeutics are now available for the treatment of diabetes, with the potential for similar delivery Reg peptides with the potential to reverse the underlying cause of diabetes.
ACKNOWLEDGEMENTThe author acknowledges Sir Frederick Banting, who even sold his Ford automobile to pay for the study animals that resulted in the discovery of insulin.
DISCLOSUREThe authors have nothing to disclose.
REFERENCES
1Barron M. The relation of the islets of Langerhans to diabetes with special reference to cases of pancreatic lithiasis. Surg Gynecol Obstet. 1920; 19: 47- 448. 2Banting FG, Best CH. Pancreatic extracts. J Lab Clin Med. 1922; 7: 464- 472. 3Banting FG, Best CH, Collip JB, Campbell WR, Fletcher AA. Pancreatic extracts in the treatment of diabetes mellitus. Preliminary report. Can Med Assoc. 1922; J 12: 141- 146. 4DeTakats G, Cuthbert FP. Surgical attempts at increasing sugar tolerance. Arch Surg. 1933; 26: 750- 764. 5DeTakats G. Ligation of the tail of the pancreas in juvenile diabetes. Endocrinology. 1930; 14: 255- 264. 6Terazono K, Yamamoto H, Takasawa S, et al. A novel gene activated in regenerating islets. J Biol Chem. 1988; 263(5): 2111- 2114. 7Kobayashi S, Akiyama T, Nata K, Abe M, Tajima M, et al. Identification of a receptor for REG (regenerating gene) protein, a pancreatic beta-cell regeneration factor. J Biol Chem. 2000; 275: 10723- 10726. 8Kim BT, Kitagawa H, Tamura J, et al. Human tumor suppressor EXT gene family members EXTL1 and EXTL3 encode alpha 1,4- N-acetylglucosaminyltransferases that likely are involved in heparan sulfate/heparin biosynthesis. Proc Natl Acad Sci U S A. 2001; 98(13): 7176- 7181. 9Watanabe T, Yonemura Y, Yonekura H, et al. Pancreatic beta cell replication and amelioration of surgical diabetes by Reg protein. Proc Natl Acad Sci USA. 1994; 91: 3589- 3592. 10Xia F, Cao H, Du J, et al. Reg3g overexpression promotes β cell regeneration and induces immune tolerance in nonobese-diabetic mouse model. J Leukoc Biol. 2016; 99(6): 1131- 1140. 11Okamoto H. The Reg gene family and Reg proteins, with special attention to the regeneration of pancreatic beta-cells. J Hepatobiliary Pancreat Surg. 1999; 6: 254- 262. 12Chen Z, Downing S, Tzanakakis ES. Four decades after the discovery of regenerating islet-derived (Reg) proteins: current understanding and challenges. Front Cell Dev Biol. 2019; 7: 235. 13Akiyama T, Takasawa S, Nata K, et al. Activation of Reg gene, a gene for a insulin-producing beta -cell regeneration: poly(ADP-ribose) polymerase binds Reg promoter and regulates the transcription by (ADP-ribosyl) autopolyation. Proc Natl Acad Sci U S A. 2001; 98(1): 48- 53. 14Saitoh S, Nishida Y, Nishida Y, et al. Distinct cell clusters touching islet cells induce islet cell replication in association with over-expression of regenerating gene (REG) protein in fulminant type 1 diabetes. PLoS One. 2014; 9(4): 1- 11. 15Newgard CPA, Hughes S, Chen L, et al. The Reg gene is preferentially expressed in the exocrine pancreas during islet regeneration. Diabetes. 1989; 38(Suppl. 1): 49A. 16Expression of reg protein in rat regenerating islets and its co-localization with insulin in the beta-cell secretory granules. Diabetologia. 1990; 33: 250- 252. 17Miyaura C, Chen L, Appel M, et al. Expression of reg/PSP, a pancreatic exocrine gene: relationship to changes in islet beta-cell mass. Mol Endocrinol. 1991; 5: 226- 234. 18Ota H, Itaya-Hironaka A, Yamauchi A, et al. Pancreatic β cell proliferation by intermittent hypoxia via up-regulation of Reg family genes and HGF gene. Life Sci. 2013; 93(18-19): 664- 672. 19Liu JL, Cui W, Li B, Lu Y. Possible roles of reg family proteins in pancreatic islet cell growth. Endocr Metab Immune Disord Drug Targets. 2008; 8(1): 1- 10. 20Lu Y, Ponton A, Okamoto H, Takasa wa S, Herrera PL, Liu JL. 55. Activation of the Reg family genes by pancreatic-specific IGF-I gene deficiency and after streptozotocin-induced diabetes in mouse pancreas. Am J Physiol Endocrinol Metab. 2006; 291(1): E50- E58. 21Zenilman ME, Perfetti R, Swinson K, Magnuson T, Shuldiner AR. Pancreatic regeneration (reg) gene expression in a rat model of islet hyperplasia. Surgery. 1996; 119(5): 576- 584. 22Zenilman ME, Magnuson TH, Swinson K, Egan J, Perfetti R, Shuldiner AR. Pancreatic thread protein is mitogenic to pancreatic-derived cells in culture. Gastroenterology. 1996; 110: 1208- 1214. 23Zenilman ME, Chen J, Magnuson TH. Effect of reg protein on rat pancreatic ductal cells. Pancreas. 1998; 17: 256- 261. 24Graf R, Schiesser M, Reding T, et al. Exocrine meets endocrine: pancreatic stone protein and regenerating protein—two sides of the same coin. J Surg Res. 2006; 133: 113- 120. 25Kapur R, Højfeldt TW, Ronn SG, et al. Short-term effects of INGAP and Reg family peptides on the appearance of small β-cells clusters in non-diabetic mice. Islets. 2012; 4(1): 40- 48. 26Rosenberg L, Vinik AI. Induction of endocrine cell differentiation: a new approach to management of diabetes. J Lab Clin Med. 1989; 114: 75- 83. 27Lipsett M, Hanley S, Castellarin M, et al. The role of islet neogenesis-associated protein (INGAP) in islet neogenesis. Cell Biochem Biophys. 2007; 48(2–3): 127- 137. 28Barbosa H, Bordin S, Stoppiglia L, et al. Islet neogenesis associated protein (INGAP) modulates gene expression in cultured neonatal rat islets. Regul Pept. 2006; 136(1–3): 78- 84. 29Li J, Wang Y, Yu X. Islet neogenesis-associated protein-related pentadecapeptide enhances the differentiation of islet-like clusters from human pancreatic duct cells. Peptides. 2009; 30: 2242- 2224. 30Levetan CS, Upham LV, Deng S, et al. Discovery of a human peptide sequence signaling islet neogenesis. Endocr Pract. 2008; 14(9): 1075- 1083. 31Nano E, Petropavlovskaia M, Rosenberg L. Islet neogenesis associated protein (INGAP) protects pancreatic β cells from IL-1β and IFNγ-induced apoptosis. Cell Death Discov. 2021; 7(1): 56. 32Xue Y, Liu C, Xu Y, et al. Study on pancreatic islet adaptation and gene expression during pregnancy in rats. Endocrine. 2010; 37: 83- 97. 33Madrid V, Borelli M, Maiztegui B, et al. Islet neogenesis-associated protein (INGAP)-positive cell mass, β-cell mass, and insulin secretion: their relationship during the fetal and neonatal periods. Pancreas. 2013; 42(3): 422- 428. 34 Mouse Genomics Informatics. Mus musculus 11 days pregnant adult female ovary and uterus cDNA, RIKEN full-length enriched library, clone:5033401N17 product: regenerating islet-derived 1, full insert sequence. http://www.ebi.ac.uk/Tools/dbfetch/emblfetch?AK133506 Accessed January 5,2021 35Assouline-Thomas B, Ellis D, Petropavlovskaia M, et al. Islet Neogenesis associated protein (INGAP) induces the differentiation of an adult human pancreatic ductal cell line into insulin-expressing cells through stepwise activation of key transcription factors for embryonic beta cell development. Differentiation. 2015; 90(4–5): 77- 90. 36Gurr W, Yavari R, Wen L, et al. A Reg family protein is overexpressed in islets from a patient with new-onset type 1 diabetes and acts as T-cell autoantigen in NOD mice. Diabetes. 2002; 51(2): 339- 346. 37 Bonner-Weir 35. S1, Toschi E, Inada A, et al. The pancreatic ductal epithelium serves as a potential pool of progenitor cells. Pediatr Diabetes. 2004; 5(Suppl 2): 16- 22. 38Inada A, Nienaber C, Katsuta H, et al. Carbonic anhydrase II-positive 36. Pancreatic cells are progenitors for both endocrine and exocrine pancreas after birth. Proc Natl Acad Sci U S A. 2008; 105(50): 19915- 19919. 39Lechner A, Habener JF. Stem/progenitor cells derived from adult tissues: potential for the treatment of diabetes mellitus. Am J Physiol Endocrinol Metab. 2003; 284: 259- 266. 40Davani B, Ariely S, Ikonomou L, Oron Y, Gershengorn MC. Human islet-derived precursor cells can cycle between epithelial clusters and mesenchymal phenotypes. J Cell Mol Med. 2009; 13(8B): 2570- 2581. 41Doniach I, Morgan AG. Islets of Langerhans in juvenile diabetes mellitus. Clin Endocrinol (Oxf). 1973; 2(3): 233- 248. 42Deng S, Vatamaniuk M, Huang X, et al. Structural, and functional abnormalities in the islets isolated from type 2 diabetic subjects. Diabetes. 2004; 53(3): 624- 632. 43Christiansen JS. What is normal glucose? – Continuous glucose monitoring data from healthy subjects. EASD, Copenhagen, 13-Sep-06. The 42nd Annual Meeting of the EASD, September 14-15, 2006, Copenhagen, Denmark 44 DCCT Research Group. The absence of a glycemic threshold for the development of long-term complications: the perspective of the diabetes control and complications trial. Diabetes. 1996; 45: 1289- 1298. 45Stratton IM, Adler AI, Neil HA, et al. Association of glycaemia with macrovascular and microvascular complications of type 2 diabetes (UKPDS 35): prospective observational study. BMJ. 2000; 321: 405- 412. 46Khaw KT, Wareham N, Luben R, et al. Glycated haemoglobin, diabetes, and mortality in men in Norfolk cohort of European prospective investigation of cancer and nutrition (EPIC-Norfolk). BMJ. 2001; 322: 15- 18. 47Cabrera O, Berman DM, Kenyon NS, Ricordi C, Berggren PO, Caicedo A. The unique cytoarchitecture of human pancreatic islets has implications for islet cell function. Proc Natl Acad Sci U S A. 2006; 103: 2334- 2339. 48Levetan C. Distinctions between islet neogenesis and β-cell replication: implications for reversal of type 1 and 2 diabetes. J Diabetes. 2010; 2(2): 76- 84. 49Guo L. Pancreatic regeneration after partial Pancreatectomy in mice mirrors that in rats with enhanced replication and Neogenesis. Diabetes. 2010; 59(suppl 1):2589-PO. 50Li Q, Li B, Miao X, et al. Reg2 expression is required for pancreatic islet compensation in response to aging and high-fat diet–induced obesity. Endocrinology. 2017; 158(6): 1634- 1644. 51Darnaud M, Cruciani-Guglielmacci C, Gonazalez P, et al. The human recombinant REG3A protein ALF-5755 restores glucose homeostasis and insulin sensitivity in high-fat fed mice and in Ob/Ob mice. Diabetes. 2020; 69(suppl 1):1064-P. 52Rosenberg L, Lipsett M, Yoon JW, et al. 86. A pentadecapeptide fragment of islet neogenesis-associated protein increases beta-cell mass and reverses diabetes in C57BL/6J mice. Ann Surg. 2004; 240(5): 875- 884. 53Siddique T, Awan FR. Effects of Reg3 Delta bioactive peptide on blood glucose levels and pancreatic gene expression in an Alloxan-induced mouse model of diabetes. Can J Diabetes. 2016; 40(3): 198- 203. 54Ilic S, Jovanovic L, Wollitzer AO. Is the paradoxical first trimester drop in insulin requirement due to an increase in C-peptide concentration in pregnant type I diabetic women? Diabetologia. 2000; 43(10): 1329- 1330. 55Sonagra AD, Biradar SM, Dattatreya K. Normal pregnancy- a state of insulin resistance. J Clin Diagn Res. 2014; 8(11): CC01- CC03. 56Ratner RE, Feeley D, Buse JB, Schwartz SL. Double-Blind, Placebo Controlled Trial of Islet Neogenesis Gene Associated Protein (INGAP) in Type 1 Diabetes (T1DM) Subjects. 65th Scientific Sessions of the American Diabetes Association. American Diabetes Association. https://professional.diabetes.org/abstract/double-blind-placebo-controlled-trial-islet-neogenesis-gene-associated-protein-ingap-type-1 57Ratner RE, Feeley D, Buse JB, Fischer JS. Double-Blind, Placebo Controlled Trial of Islet Neogenesis Gene Associated Protein (INGAP) Therapy in Type 2 Diabetes (T2DM) Subjects. 65th Scientific Sessions of the American Diabetes Association. American Diabetes Association. https://professional.diabetes.org/abstract/double-blind-placebo-controlled-trial-islet-neogenesis-gene-associated-protein-ingap
留言 (0)