
記住我





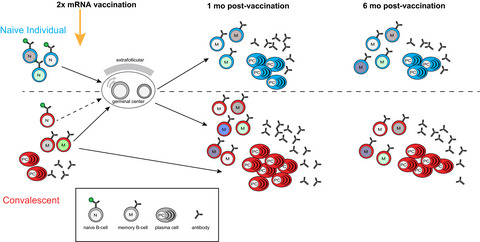


In a new study published in Science, Goel et al.1 evaluate immune responses against SARS-CoV-2 in vaccinated individuals and find evidence of durable immune memory for at least 6 months, irrespective of former infection (Figure 1).

Similar immune responses in naive individuals and COVID-19 convalescents receiving COVID-19 mRNA vaccines. (Top) Naive individuals receiving two doses of an mRNA vaccine generate a diverse somatically mutated circulating memory B cell population of a similar size to COVID-19 convalescents (Bottom) by 6 months post double-dose vaccination. More antibody-producing plasma cells are generated in the convalescents due to their pre-existing memory B cell population.
The SARS-CoV-2 virus causing COVID-19 is redefining human endeavor across the planet with considerable healthcare and economic burden. For immunologists, it has provided an unprecedented opportunity to learn the dynamics never-before-seen virus regarding transmissibility, epidemiology and the utility of vaccines in combating emergent health threats. Already in the first years of the pandemic, numerous studies have traced patient cohorts following SARS-CoV-2 infection for evidence of robust immune responses and memory. The formation of high amounts of pathogen-specific antibody is a key aspect of the protective immune response. In SARS-CoV-2 infection and vaccination, antibodies targeting the Spike protein are particularly important, as these can block virus binding to angiotensin conversion enzyme-2 on host cells, thereby neutralizing the virus.2 SARS-CoV-2 emerging variants of concern (VOC) contain mutations in the Spike protein, and particularly those in the receptor-binding domain (RBD) have been found to either enhance transmissibility (alpha and delta VOC) or to escape from antibody binding (beta and gamma VOC).
High-affinity antibodies are produced by plasma cells, terminally differentiated B cells that have cycled through a germinal center (GC) response. GCs are transient anatomical structures in secondary lymphoid organs in which activated B cells proliferate and receive help from T cells to mutate their genes encoding the pathogen-recognizing B cell receptor (BCR), the membrane-bound form of antibody. Mutations that improve pathogen recognition lead to the selective retention of B cells in the GC, such that the resultant pool of Bmem and plasma cells have improved affinity, and carry more mutations when they exit the GC at later times. In convalescence, antibody responses peak after ˜30 days and then contract. In contrast, circulating memory B (Bmem) cell numbers peak later (50–100 days) and remain more stable over 8 months.3, 4 Recent studies by Goel et al.5 showed that the first dose of an mRNA vaccine (Pfizer or Moderna) induces greater antibody responses in COVID-19 convalescents than formerly naïve individuals (FNI), demonstrating the functional relevance of this previously formed memory.
In this Science publication, Goel et al.1 followed up their convalescent and FNI cohort for several months after two doses of mRNA vaccine. Despite COVID-19 convalescents generating approximately 10-fold more peak antibody than FNI, these peaked at approximately similar times after the second dose, and then declined at similar rates. A parallel decline in antibody levels is intriguing, suggesting that the lifespans of plasma cells is the same, irrespective of former infection status.
One of the key questions about SARS-CoV-2 memory is whether vaccination will protect against VOC. The authors demonstrated that the elicited antibodies were able to neutralize Spike RBD from the Wuhan strain and VOC in the majority of individuals for at least 6 months, i.e. the duration of the study.1 Similarly, Bmem cell responses, as assessed by flow cytometry using an impressive suite of fluorescent tetramers of Wuhan and variant Spike proteins, increased after vaccination and remained high 6 months later. Importantly, the BCR of memory B cells in convalescents and FNI shared several common features, including V-gene usage patterns and somatic mutation densities, suggesting the protective immune response was qualitatively similar, irrespective of prior COVID-19 infection (Figure 1). The Bmem cells that recognized Spike RBD from multiple VOC tended to possess more mutations than those recognizing Wuhan Spike RBD alone, suggesting that ongoing GC processes are important for generating broadly protective immunity. This is an elegant demonstration of somatic evolution and diversification in response to a novel antigenic target.
The Bmem cell responses evoked during vaccination are supported by memory T cells (Tmem). The authors also provide insight into the dynamics of SARS-CoV-2-induced Tmem and elaborate further on this in a sibling publication.6 Vaccine-induced Tmem were identified following in vitro stimulation of peripheral blood mononuclear cells (PBMCs) with peptides from the Spike protein and the detection of activation-induced molecule (AIM) expression on T cells. Longitudinal analysis of blood samples with this AIM assay revealed that CD4+ and CD8+ Tmem cell responses peaked approximately 3 weeks after the second vaccine dose. Interestingly, the CD4+ T cell response declined at a similar rate to the initial antibody response, with a half-life of approximately 47 days. In contrast, CD8+ T cells contracted more rapidly, with a half-life of 27 days. Notably, the magnitude of the CD4+ T cell response soon after the second vaccine dose was positively correlated with the amount of antibody at 6 months, and could be a predictor of the antibody response. Overall, the study provides a holistic sense of COVID-19 responses: B cell responses are diverse, and presumably supported by T cell responses. Despite being quantitatively greater in convalescents, the functional properties and contraction of the B and T cell responses are reasonably parallel in convalescents and FNI following double-dose vaccination (Figure 1).
The Goel et al. study is one of three recent studies demonstrating cross-protection against VOC within the Bmem cell compartment by vaccination.1, 7, 8 The study affirms work by another group that broadly targeting BCR specificities tends to associate with more somatic mutations in the BCR.7, 8 However, by the nature of its timing and design, the study only assessed otherwise healthy individuals who had been infected with the Wuhan strain. This has changed considerably in 2021 with beta, gamma and especially the delta variant being more prevalent, making it intriguing to find out how vaccination with current Wuhan Spike protein vaccines will boost immune memory generated by infection with those VOC. Also, individuals with secondary health conditions or primary immunodeficiencies may not generate immunity in the same way as otherwise healthy individuals, so how vaccine-driven immune responses to VOC track in the most at-risk individuals for hospitalization and death will need to be addressed in the future.
While these studies suggest that VOC can be recognized well by vaccination responses to the original Wuhan Spike RBD, it remains possible that future evolution of VOC that evade immune protection would trigger the need to adapt COVID-19 vaccines in much the same way that yearly influenza vaccines are updated with new strains if these escape antibody responses. It is also possible that the protection engendered by vaccination reduces to non-protective levels each year, and we will thus require yearly boosters irrespective of cross-protection. Such a scenario would also raise the question of whether original antigenic sin, the concept that the original B-cell specificities recruited into the response will then dominate a response to escape variants, might limit the capacity of the immune system to respond to a novel mutant. From the cross-reactivity data in the current study,1 it would seem unlikely, but the specificities of vaccines may need to be adaptable in the future.
In the current study,1 modeling suggested that protective antibody amounts would diminish significantly in 7–12 months, suggesting on the one hand that a booster might be required after that time. On the other hand, neutralizing antibodies in blood do not protect against infection, which takes place in the upper airways. The delta strain, for example, is more prone to breakthrough infections in vaccinated healthcare workers than other strains, despite immune cross-reactivity.9 It would be important to assess whether protection against severe disease is more closely associated with antibody amounts or with Bmem and Tmem cell numbers. Further, exactly what immune response features might increase the duration of immune protection are currently unknown, but understanding them would be a leap forward in the field.
One minor caveat of the Goel study is that it maps only the response against variant RBD domains, whereas the antibody escape capacity of the beta variant is thought to result from mutations in other regions of the Spike protein. Thus, the breadth of immune memory recorded by RBD analysis might not reflect true neutralizing capacity. Outcomes for hospitalization and death upon re-infection may also differ from the elicited immune profiles. Certainly, in active SARS-CoV-2 infection, the magnitude of the immune response can predict the likelihood of hospitalization,10 making such correlations in a convalescent setting of interest. However, as the RBD by definition is integral to viral infection, the study does provide strong evidence for cross-recognition that will impair viral entry.
In summary, Goel et al.1 demonstrate the common evolution of B and T cell memory in COVID-19 convalescents and FNI, suggesting most of the benefit of tertiary boosting will come in the form of higher antibody amounts. Vaccination also leads to cross-protection against the Spike protein RBD domain of three circulating VOC, suggesting that a broad benefit of boosting with the current vaccines against VOC is likely. Thus, COVID-19 vaccination induces a memory response that is inherently diverse and inclusive, an outcome that is good for all.
Conflict of InterestThe authors declare no conflicts of interest.
AUTHOR CONTRIBUTIONSGemma E Hartley: Visualization; Writing-review & editing. Menno C van Zelm: Writing-original draft; Writing-review & editing. Marcus J Robinson: Writing-original draft; Writing-review & editing.
留言 (0)