
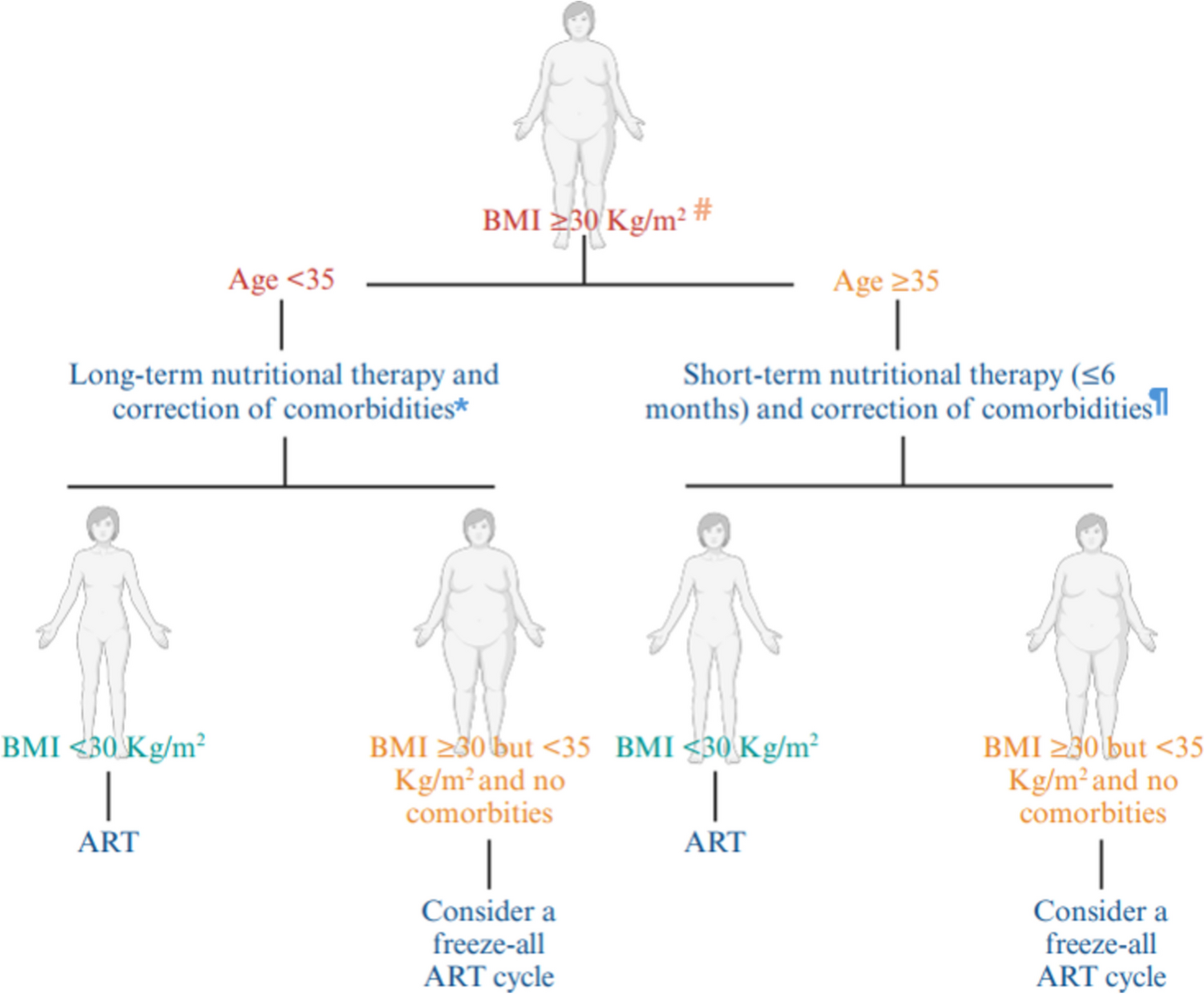
記住我







Bovine ovaries were collected from a slaughterhouse and transported to the laboratory in phosphate-buffered saline (PBS) within 3 h. Cumulus-oocyte complexes (COCs) were aspirated using an 18-gauge needle connected to a 10-mL syringe. The COCs were cultured in a 100-μL droplet of in vitro maturation medium (10 oocytes/drop) covered by paraffin oil (tissue culture grade; Nacalai Tesque) for 21 h. TCM-199 medium (Gibco, Grand Island, NY, USA) supplemented with 10% fetal calf serum (FCS) (5703H; ICN Pharmaceuticals, Costa Mesa, CA, USA), 5 mM taurine, and 10-ng/mL epidermal growth factor was used as the maturation medium. Then, the COCs were incubated with freeze-thawed semen from Japanese black cows, as described previously [14]. In vitro fertilization and in vitro culture (IVC) media were based on synthetic oviductal fluid, modified according to previous reports [15]. After fertilization, the COCs were incubated in IVC medium for 48 h in an atmosphere of 5% CO₂. Embryos cleaved over 8-cells were selected and used for experiments.
Vitrification and warming of 8-cell embryoVitrification medium TCM199 (pH 7.3) containing 6.25 mM HEPES and 20% FCS was referred to as the rinsing solution (RS). Equilibration was performed in RS containing 7.5% DMSO + 7.5% EG or 7.5% PG + 7.5 EG (equilibration solution; ES) for 5 min at 38.5 °C. Vitrification was then performed in RS solution containing 15% DMSO + 15% EG or 15% PG + 15% EG (vitrification solution; VS). Embryos were treated with VS for 30 s and placed on a Cryotop device (Kitazato Crop, Shizuoka, Japan) and plunged into liquid nitrogen. for warming, the Cryotop devices with embryos was immersed directly into RS containing 1.0 M sucrose and incubated for 1 min at 38.5 °C, followed by a step wise transfer into RS containing 0.5, 0.25, and 0 M sucrose for 3, 5, and 5 min, respectively.
ImmunostainingVitrified-warmed embryos were incubated for 1 or 5 days and cleaved embryos or blastocyst-stage embryos that appeared morphologically viable were selected and used for immunostaining. Embryos were fixed using 4% paraformaldehyde for 1 day and permeabilized using PBS containing 0.2% Triton X-100 for 30 min, followed by blocking in PBS containing 5% bovine serum albumin for 1 h. The embryos were incubated overnight with the primary antibody, followed by treatment with secondary antibody for 1 h. For immunostaining of 5mC and 5hmC, the embryos were treated with 1 N HCl for 1 h before blocking. The embryos were then mounted on glass slides and observed under a fluorescence microscope (SCR-038447; Dianova Hamburg, Germany). To determine the expression levels of 5mC and 5hmC in the 8-cell stage, all blastomere nuclei were measured. For blastocysts, non-overlapped 5 trophectoderm (TE) cells and five inner cell mass (ICM) cells were arbitrarily selected to measure 5mC expression levels. To evaluate the expression levels of other proteins (DNMT1, DNMT3A, TET1, and TET3) in cleaved embryos, the equatorial region of the whole embryo was evaluated. The primary and secondary antibodies used are described in Table 1. In each trial, average value of control (fresh, non-vitrified embryos) is defined as 1.0, and ratio of experimental groups were calculated.
Table 1 Antibodies used for immunostainingAssessment of lipid content in embryosVitrified-warmed embryos were cultured for 1 or 5 days and cleaved embryos or blastocyst-stage embryos that appeared morphologically viable were stained with Nile red (Wako, Tokyo, Japan) for 10 min. The embryos were mounted on a glass slide and the equatorial region of the embryos was imaged under a fluorescent microscope (SCR-038447; Dianova; Hamburg, Germany). The images were digitized using the ImageJ software (NIH).
RNA-seq and analysis of the dataIn vitro-developed blastocyst-stage embryos were used for RNA-seq analysis. Embryos were vitrified, warmed at 8-cell stage, and cultured up to the blastocyst stages for 5 days. Approximately 30 blastocysts developed from fresh DMSO + EG or PG + EG 8-cell stage vitrified embryos were used for RNA extraction. RNA quality and concentration were examined using the Agilent 2100 Bioanalyzer (Agilent Technologies, Palo Alto, CA, USA). The average RNA integrity number was 9.4 ± 0.4. cDNA was produced using the NEBNext single cell/low input RNA library prep kit (New England Biolabs). The quality and quantity were determined using an Agilent 2100 Bioanalyzer, followed by re-measurement using at Kapa library quantification kit (Kapa Biosystems, Wilmington, MA, USA). Sequencing was conducted using NextSeq1000 (Illumina, San Diego, CA, USA) with single read × 100 bp. Image analysis, base calling, and quality filtering were performed using RTA version 2.4.11 (Illumina) following the manufacture’s protocol, and the sequence data were converted to Fastq using bcl2fastq2 v2.20.0.422. To prepare the sequenced data, adapter sequences, ambiguous nucleotides, and low-quality sequences were removed. The remaining sequence data were aligned to the Bos taurus genome sequence (ARS-UCD1.2/bosTau9) to count sequence reads. Gene expression values were evaluated as transcripts per kilobase million. The process of sequence preparation, mapping to the reference genome, and differential gene expression analysis (P < 0.05) were performed using the CLC Genomics Workbench ver. 22.0.2 (Qiagen, Hilden, Germany). Pathways and Gene Ontology (GO) analyses of the differentially expressed genes (DEGs) were conducted using a functional annotation tool (DAVID, https://david.ncifcrf.gov/), where the Bos taurus genome was used as the reference. DEGs were compared using the principal component analysis (PCA) and K-medoids method. The raw RNA-seq data of the blastocysts were registered in DDBJ under accession numbers PRJDB18767 and Experiment DRX576597-576,605. DEGs between the control and DMSO groups (P < 0.05) were clustered into eight groups based on the differences between the DMSO and PG groups using the K-medoids method (CLC Genomics Workbench). K-medoids can assign the DEGs to clusters using an algorithm [16]. In K-medoids clustering, DEGs are clustered into K separate clusters. The procedures seek to assign DEGs to cluster such that distances between DEGs of the same cluster are small while distances between clusters are large. After clustering, DEGs with the greatest differential directional changes in DMSO/control vs. PG/DMSO groups were selected (Fig. 1a) and used for the prediction of Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways.
Fig. 1
Schematic design of K-medoids analysis of DEGs. a DEGs between control and DMSO groups (P < 0.05) were clustered into eight groups (a–h). a Gene groups (d and e) having the greatest adverse directional change between DMSO and PG compared with those between control and DMSO were selected. b Pathway enriched by the genes groups (d, e)
Experimental designIn the first experiment, 8-cell stage embryos were vitrified using DMSO + EG or PG + EG. The warmed embryos were cultured to the blastocyst stage, and the rate of development was examined (29 trials with 10–12 embryos each). A total of 324, 324, and 319 8-cell stage embryos were used for control, DMSO + EG, and PG + EG groups, respectively. In addition, fresh (control) or vitrified-warmed 8-cell stage embryos were incubated for 24 h when morphologically viable embryos were subjected to immunostaining for 5mC and 5hmC. Blastocysts derived from fresh or vitrified 8-cell stage embryos were immunostained for 5mC.
In the second experiment, to determine whether vitrification using DMSO or PG affected the expression levels of DNMT1/3A and TET1/3, fresh or vitrified-warmed 8-cell stage embryos were cultured for 1 day and then subjected to immunostaining. Furthermore, warmed 8-cell stage embryos were cultured to obtain blastocysts, in which lipid content was measured.
In the third experiment, 8-cell stage embryos were vitrified using DMSO or PG, and the warmed embryos were incubated for 5 days. A comprehensive gene expression analysis was conducted using blastocysts. Thirty embryos were prepared for each group, and this was repeated three times with a differential ovary series.
In these four experiments, we hypothesized that the oxidative properties of DMSO were causal factors for demethylation. 5 mM NAC (M8H6852; Nacalai Tesque, Inc; Kyoto, Japan) was added to the equilibration and vitrification solution containing DMSO + EG. Vitrified-warmed embryos were stained for 1 and 5 days, after which the embryos were stained for 5mC.The rate of developmental into the blastocyst stage was examined (29 trials with ten embryos).
Statistical analysisAll data were analyzed using the Shapiro–Wilk test. Three parametric datasets were analyzed using one-way analysis of variance (ANOVA), followed by Tukey’s post hoc test. Non-parametric data were analyzed using the Kruskal–Wallis and Steel–Dwass tests for multiple comparisons. Parametric data were analyzed using Student’s t test and non-parametric data were analyzed using the Mann–Whitney U test. Date with P < 0.05 was considered statistically significant.
留言 (0)