
記住我







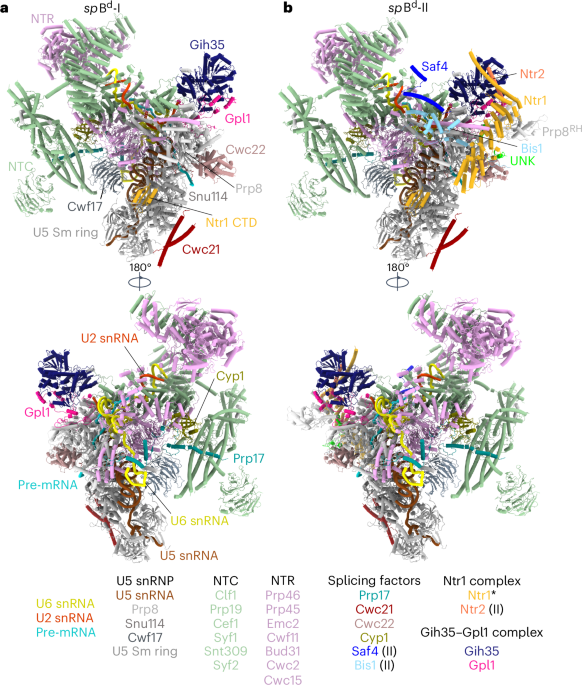
Our cryo-EM map of the supercomplex has an average resolution of 3.2 Å (according to local refined map resolutions; Extended Data Fig. 4) but well-ordered regions are locally resolved up to 3 Å (Extended Data Fig. 4). The supercomplex consists of one NDH and one PSI–LHCI unit (Fig. 1a–c), interacting directly at the terminus of the transmembrane module of NDH and Lhca6 of PSI–LHCI-2 (refs. 7,9). Our cryo-EM density map revealed 26 of the 29 NDH subunits10 and all 16 subunits of the PSI–LHCI-2 complex11, including 154 chlorophylls, 37 carotenoids and 46 lipids. NDH is organized into five modules: SubA, SubB, SubE, SubL and SubM. Modules SubA (NdhH–NdhO) and SubE (NdhU) form the peripheral arm. In the SubE module, which is specific to oxygenic photosynthesis, we identified subunit NdhU, while NdhS, NdhT and NdhV12,13,14,15 were absent.
Fig. 1: Cryo-EM structure of the NDH–PSI–LHCI-2 supercomplex from S. oleracea.
a–c, Lateral (a,b) and top (c) views of the cryo-EM map of the supercomplex (contour level: 0.38). Shades of cyan (module SubA, NdhH–NdhO) and magenta (module SubE, NdhU) indicate the NDH peripheral arm. The membrane arm is drawn in shades of blue (module SubM, NdhA–NdhG). Lumenal SubL subunits (PnsL1–PnsL5) are shown in shades of purple and subunits of the stromal SubB (PnsB1–PnsB5) in shades of gray. PSI–LHC-2 is colored in shades of green. Cryo-EM densities of lipids are salmon (PG), violet (MGDG), pink (SQDG) or green (DGDG). Carotenoids are orange and chlorophylls are bright green. The detergent belt is transparent gray. d, The peripheral arm contains a row of three 4Fe–4S iron–sulfur clusters (N6a, N6b and N2; Fe, orange; S, yellow) closely spaced for direct electron transfer. N2 is next to the PQ cavity that bifurcates into the PQ entry channel and the E-channel. The distance between the putative PQ molecule bound in the entry channel and N2 is too far for direct electron transfer. e, Structure of the PSI cofactors with corresponding map densities (contour level: 0.22). Subunits participating in electron transfer are indicated. PSI contains three 4Fe–4S centers (FB, FA and FX). The protein scaffold holds the special-pair chlorophylls (P700), pheophytins (A1A/B and A0A/B) and PhyQ in two near-symmetrical membrane-spanning branches that converge on Fe–S cluster Fx.
The membrane arm extends from SubB (PnsB1–PnsB5) on the stromal side to SubL (PnsL1–PnsL5) on the lumenal side. These subunits are unique to chloroplast NDH and absent in cyanobacteria10,16,17,18,19,20. On the stromal side, PnsB1 interacts with NdhB, NdhD, NdhH, NdhF, PnsB4 and PnsB5 (Fig. 2a–e). PnsB3 has four conserved cysteine residues (Extended Data Fig. 5d) expected to bind an iron–sulfur cluster21, although we did not find an Fe–S density in this position.
Fig. 2: The long N-terminal loop of PnsB1 interacts with NdhB residues likely to be involved in proton translocation.
a, Lateral view of the supercomplex. The subunits interacting with PnsB1 are color-coded as in Fig. 1. PnsB1 is shown as a ribbon; other subunits are shown as an outline. The highlighted region illustrates the points of contact between the long N-terminal loop and the subunits indicated. b, Main contact points between the PnsB1 N-terminal loop and the subunits below. Each group of electrostatic interactions is marked with a letter and shown in the corresponding panels (c–e).
The peripheral arm of plant NDH binds three iron–sulfur clusters precisely spaced for efficient electron transfer. NdhI coordinates two 4Fe–4S clusters and NdhK coordinates one (Fig. 1d). PQ binds in the PQ cavity at the junction between the peripheral and membrane arms (Fig. 1d).
Spinach PSI–LHCI-2 closely resembles the Pisum sativum PSI–LHCI complex22,23, consisting of 12 core subunits (PsaA–PsaL) and four LHCI antenna complexes (Lhca1, Lhca3, Lhca4 and Lhca6; Fig. 1b,c). As observed in NDH–PSI–LHCI-2 supercomplexes from other plants8,9, Lhca6 interacts with NDH, replacing Lhca2 (Fig. 1a–c). The PSI core subunits, PsaA and PsaB, form a conserved heterodimer22,23,24, with the reaction center situated at the interface of their C termini. This assembly includes two membrane-spanning branches, A and B, each featuring one chlorophyll P700, two pheophytins (A1A/B and A0A/B) and a phylloquinone (PhyQ) (Fig. 1e). The terminal electron acceptors are three 4Fe–4S clusters (FX, FA and FB), which subsequently reduce Fd. FA and FB are bound to PsaC, while FX is coordinated by four pseudosymmetrical cysteine residues from PsaA and PsaB (Extended Data Fig. 5c). On the stromal side, peripheral subunits PsaC, PsaD and PsaE form the Fd-docking site. PsaG and PsaI are unique to angiosperms and algae25. PsaF, PsaA and PsaB together create a potential docking site for the soluble electron transfer protein plastocyanin26,27.
In our PSI–LHCI-2 density map, we observed 154 chlorophylls, matching the count in PSI–LHCI-2 from A. thaliana but exceeding the 148 chlorophylls in PSI–LHCI-2 from H. vulgare (Supplementary Fig. 1a,c). Additionally, PSI–LHCI-2 from S. oleracea has more chlorophylls than PSI–LHCI-1 from both A. thaliana (152) and barley (148) (Supplementary Fig. 1d,e). This discrepancy is most apparent in subunit Lhca5, which in PSI–LHCI-1 interacts with NDH (chlorophylls labeled a, b and z in Supplementary Fig. 1d,e), suggesting that differences in pigment content reflect the variation in subunit composition and that the presence or absence of particular chlorophylls might be required for energy transfer in PSI–LHCI-1.
NDH–PSI–LHCI-2 interactionIn the spinach supercomplex, Lhca6, NdhF and PnsB5 stabilize the connection between NDH and PSI–LHCI-2 (Fig. 3a,b). Numerous electrostatic, polar and hydrophobic interactions contribute to supercomplex formation and stability, primarily on the stromal side (Fig. 3c–h). Lhca6 interacts with NdhF mainly through its transmembrane helix 2 (TMH2)–TMH3 loop (Fig. 3c–e). One electrostatic interaction was found on the lumenal side between the TMH1–TMH2 loop of Lhca6 and the N-terminal helix of NdhF (Fig. 3f). The extensive N-terminal stromal loop of PnsB5 engages with both PnsB1 (Extended Data Fig. 6a,b) and NdhD (Extended Data Fig. 6a,c,d), passing above NdhF (Extended Data Fig. 6a,c–e) and forming a hook around the N-terminal stromal loop of Lhca6 (Fig. 3g and Extended Data Fig. 6e). A detergent molecule, likely replacing a lipid, appears to reinforce the connection between TMH3 of Lhca6, the N-terminal stromal loop of PnsB5 and the α-helix motif linking TMH1 and TMH2 of NdhF (Fig. 3h). The loop connecting β-strands 4 and 5 of PnsB2 closely approaches the TMH2–TMH3 loop of Lhca6 (Fig. 3c). Contrary to expectations28, we did not observe direct interactions of NDH PnsB2 and PnsB3 with PSI–LHCI-2.
Fig. 3: The N-terminal loop of PnsB5 is central to supercomplex formation and stability.
a, Top view of the supercomplex. The subunits connecting the two complexes are shown as ribbons (NdhF, dark blue; PnsB5 dark gray; Lhca6, spring green) while the other subunits are shown as outlines. Key subunits are color-coded as in Fig. 1. b, Highlighted region illustrating the points of subunit contacts. c–h, Interactions in detail.
The NdhU subunitThe local resolution of the peripheral arm of spinach NDH was 3.3 Å, enabling detailed mapping of subunit interactions at the level of individual side chains in the SubA and SubE modules. Biochemical assays12,29,30 and cryo-EM structures of cyanobacterial NDH31,32 both revealed that SubE is necessary for the interaction of the complex with Fd. In addition to the subunits found in cyanobacterial NDH, we discovered a previously unresolved subunit (Fig. 4a and Extended Data Fig. 2) that we identified on the basis of its resolved side-chain sequence (Fig. 4a) as NdhU of the SubE module12. AlphaFold33 models for NDH subunits previously identified by mass spectrometry12,13,14 (Fig. 4b,c) suggested that either NdhT (Fig. 4c, yellow) or NdhU (Fig. 4c, magenta) was compatible with this new EM density (Fig. 4e). Both models featured a J-shaped domain that was visible in the map but side-chain densities of aromatic residues identified the subunit as NdhU unambiguously (Fig. 4d–h).
Fig. 4: Identification of NdhU from its cryo-EM density.
a, Cryo-EM map of the unidentified subunit. b, List of nuclear-encoded subunits and protein factors with established interactions with NDH observed in the model organism A. thaliana. c, Panel with the computational models of the proteins listed in b. The designated subunits were produced with AlphaFold33 from S. oleracea input sequences. d–h, On the basis of the AlphaFold predictions and the results from Yamamoto et al.12, we attempted to fit both NdhT and NdhU into the cryo-EM map shown in a. Side chains of NdhU (magenta) fit the density map well, firmly establishing the identity of this subunit. The cryo-EM map shown in a,e,g is drawn at a contour level of 0.3. In d,f,h the contour level is 0.45 to show the side chains more clearly.
NdhU has a flexible N-terminal loop and a J-shaped domain composed of four α helices (α1–α4), followed by a short helix (α5) (Extended Data Fig. 7a). An extended loop connects α5 to the transmembrane domain (Extended Data Fig. 7a). NdhU is centrally placed in the SubE module, establishing electrostatic interactions and salt bridges with subunits NdhI, NdhH, NdhK and NdhJ on the stromal side and potentially with NdhA on the lumenal side (Extended Data Fig. 7e). Its long N-terminal loop and J-shaped domain interact with NdhJ forming an aromatic cluster with NdhK, anchoring it to the peripheral arm (Extended Data Fig. 7b,c). Additionally, the loop connecting the soluble part to the NdhU TMH interacts with the surface of the SubA module (Extended Data Fig. 7d,f).
Bound lipidsWe modeled a total of 46 lipids into distinct densities in the spinach supercomplex map, of which 23 are in the NDH membrane arm and 23 are in PSI–LHCI-2 (Extended Data Fig. 8a,b). In NDH, we found densities for 14 phosphatidyl glycerol (PG), 5 monogalactosyl diacylglycerol (MGDG) and 3 sulfoquinovosyl diacylglycerol (SQDG) lipids (Extended Data Fig. 8a) but, surprisingly, no digalactosyl diacylglycerol (DGDG). Most lipids are associated with transmembrane subunits (Fig. 5, middle) and many fill gaps between neighboring subunits, acting as a hydrophobic adhesive (Fig. 5a–i). The negatively charged PG head groups connect loops and helices (Fig. 5a–c,g–i). Two adjacent PG molecules link the N-terminal loop of NdhL to NdhA (Fig. 5b), as in cyanobacterial NDH32 (Supplementary table 1a). Nested within the NDH core, we detected three well-defined SQDG molecules at the juncture between NdhB, NdhD and PnsB1 (SQDG 522/B; Fig. 5d), between NdhD and NdhF (SQDG 806/D; Fig. 5e) and between NdhF and PnsB5 (SQDG 1015/F; Fig. 5f). SQDG 1015/F, together with two PGs and a molecule of β-carotene8,32,34 (BCR 1003/D), connect NdhD, NdhF, PnsB5 and PnsB4 (Fig. 5f).
Fig. 5: Lipids in the NDH membrane arm.
Middle, subunits of the NDH membrane arm in outline, with NdhA, NdhB, NdhD and NdhF color-coded. Stromal and lumenal subunits were removed for clarity. Lipid densities are salmon (PG), violet (MGDG) or pink (SQDG). Detergent (t-PCCαM (A1H1M)) density is turquoise. a–i, Several lipids interact with more than one subunit, presumably enhancing supercomplex stability.
Of the 23 lipids modeled into the PSI–LHCI-2 complex, PG is the most abundant, followed by MGDG (Extended Data Fig. 8b). The lipid distribution within PSI–LHCI-2 is striking. While PG and DGDG are located at the interface between LHCIs and PSI or within the PSI core (Extended Data Fig. 9, middle), MDGD and SQDG are confined to the border between the LHCs and PSI (Extended Data Fig. 9, middle; violet and pink densities respectively). At the interface with Lhca1, Lhca6 and Lhca3 (Extended Data Fig. 9a,c,f,h), a prominent string of lipids connects the LHCI belt to PSI. MGDG in this region is consistent with the crystal structure of P. sativum PSI–LHCI22,23 (Supplementary table 1b). Some PG lipids bind closely to chlorophyll, creating a ligand for the central Mg2+ (Extended Data Fig. 9b–e). The symmetrical arrangement of one PG (PG 1064/a) and one DGDG (DGDG 850/b) in the PSI core is conserved across species (Extended Data Figs. 8b and 9e).
The PQ-binding pocketAt the juncture of the membrane arm and the peripheral arm, the HOLLOW software tool35 revealed a bifurcated cavity. This cavity comprises the PQ entry channel, E-channel and PQ-binding pocket, closely resembling the homologous Q entry channel, E-channel and Q-binding pocket of complex I (Figs. 1d and 6a,b). Within the PQ entry channel, we observed a distinct nonprotein density of approximately 26 Å in length (Fig. 6b, red) in a position that corresponds to the shallow site in mitochondrial complex I36,37. Guided by previously published structures of cyanobacterial NDH32 and complex I36,38, we modeled a PQ molecule in the density (Fig. 6c). The PQ sits at the entrance of the cavity surrounded by several charged residues, interacting with Y259 of NdhA through its carbonyl group (Fig. 6c). The PQ head group is directed toward the N2 Fe–S cluster at an approximate distance of ~24 Å (Fig. 1d), too far for direct electron transfer. Although we cannot rule out t-PCCαM (4-trans-(4-trans-propylcyclohexyl)-cyclohexyl α-maltoside), the detergent used for solubilizing and purifying the supercomplex, the density in the PQ pocket looks too long and bulky for a detergent molecule and PQ fits it almost perfectly (Fig. 6c).
Fig. 6: The PQ-binding pocket.
a, Lateral view of the peripheral arm with subunits forming the PQ chamber (NdhH, NdhK and NdhA) shown as cylinders and surrounding subunits in outline. b, Secondary-structure elements defining the PQ pocket. The PQ density is red and the cavity is shown as a gray surface. c, Close-up view of the PQ molecule modeled into the density (red) visualized at a contour level of 0.23. PQ interacts with Y259. d, TMH5–TMH6 loop of NdhA and the density map (light gray) at a contour level of 0.4. Residues making intrasubunit and intersubunit interactions are shown as sticks. e, Salt bridges formed between the NdhC TMH1–TMH2 stromal loop and surrounding subunits. The density of the NdhC TMH1–TMH2 loop is shown in gray at a contour level of 0.4. Backbone carbonyl groups involved in side-chain interactions are highlighted red in d,e.
The PQ-binding pocket is formed by subunits NdhA, NdhH and NdhK (Fig. 6a,b). As in cyanobacterial NDH32, the PQ pocket is lined by the four-helix bundle and the loop connecting the first two β-strands (β1–β2 loop) of NdhH. The highly conserved Y72 residue approaches Fe–S cluster N2 closely (Fig. 6b). The PQ pocket is further lined by the second helix (α2) and the loops α1–α2 and α3–α4 of NdhK and the long loop connecting TMH5 and TMH6 of NdhA. This loop engages in numerous intrasubunit and intersubunit interactions, including three salt bridges between NdhA and NdhH (Fig. 6d). Note that the TMH5–TMH6 loop is not fully resolved in PQ-bound cyanobacterial NDH32.
The special confinements of the PQ pocket are lined with bulky hydrophobic residues (Fig. 6b). In the NdhH β1–β2 loop, the highly conserved residues H23 and H19 are oriented outward (Fig. 6b), leaving the path to Y72 open. The tyrosine residue (Y39) in the NdhK α1–α2 loop is not conserved in complex I but present in both plant and cyanobacterial NDH.
The PQ-binding pocket is formed by subunits NdhC, NdhI, NdhJ, NdhL, NdhM, NdhO and NdhU (Fig. 6a,e), of which NdhC, NdhI, NdHL and NdhU link the membrane arm to the hydrophilic arm of NDH. The long stromal loop connecting the first and second TMHs of NdhC (TMH1–TMH2 loop; Fig. 6e) connects to NdhA and the soluble subunits NdhK, NdhH and NdhM with multiple salt bridges, reinforcing the interactions between the two arms of NDH (Fig. 6e).
The transmembrane proton pathwayThe transmembrane module of NDH comprises seven subunits (NdhA–NdhG). Of these, NdhA, NdhC, NdhE and NdhG form the E-channel37 (Fig. 7a,b) and NdhB, NdhD and NdhF correspond to the antiporter-like subunits of complex I38 (Fig. 7a). Together, they constitute the four proton-pumping units known from complex I5,37,39. Each antiporter-like subunit contains two discontinuous helices (TMH7 and TMH12, Fig. 7a) that are integral to the formation of conserved internal cavities within the membrane arm (Fig. 7a, gray surfaces).
Fig. 7: Proposed proton transfer path in the membrane arm of S. oleracea NDH.
a, Side view of NDH. From right to left, NdhA helices and NdhH and NdhK loops contributing to the formation of the PQ-binding pocket (gray surface), NdhC TMH1–TMH2, NdhG TMH3, NdhE TMH2–TMH3 and discontinuous TMHs of NdhB, NdhD and NdhF are shown as solid cylinders. Other subunits are shown as outlines. The 4Fe–4S N2 cluster is indicated. Charged residues of NdhA–NdhG and residues involved in proton translocation that are conserved between spinach NDH and A. thaliana complex I are shown as stick models and are color-coded. Positively charged residues are blue, negatively charged residues are red and key aromatic residues are white. PQ pocket, voids, channels and depressions within NDH that are likely to be relevant for proton translocation are shown as gray surfaces. The light-blue arrow delineates a continuous hydrophilic path from the Q-binding site to NdhF. Stromal half-channels above NdhB, NdhD and NdhF are indicated as dotted cyan arrows and labeled 1–3; the channel between NdhB and NdhD (magenta) is labeled 4. b, Top view of the E-channel with the relative map of voids presumably occupied by water molecules. Key residues are labeled. c, TMH3 of NdhG. Conserved residues in A. thaliana complex I important for proton translocation are shown in stick representation. The cryo-EM map (transparent cyan) is at a contour level of 0.4. d, Top view of the channel entry above NdhB shown as a surface (1 in a). Subunits of to the entry channel are shown as electrostatic potential surfaces (red, negative; blue, positive). The PnsB1 N-terminal loop is shown in cartoon representation. e,g, Side (e) and top (g) views of channel NdhB-D (4 in a). Polar residues defining the putative hydrophilic channel are shown as sticks. In e, the asterisk indicates the constriction point shown in f. f, Close-up view of the constriction in channel NdhB–NdhD. As in e, the asterisk indicates the constriction point. In g, elements from PnsB1, PnsB4 (not visible), PnsB5, NdhB and NdhD form a funnel, depicted as electrostatic potential surfaces (negative, red; positive, blue) above the stromal entry. The subunits involved in forming the funnel are shown in cartoon representation. Charged residues are represented as sticks.
As in cyanobacterial NDH32, we observed that NdhA, NdhH and NdhK subunits enclose a branched cavity (Fig. 7a, gray surface). The primary branch lined by NdhA helices forms the access channel for PQ toward the PQ entry channel and the PQ-binding site in subunits NdhH and NdhK (Fig.
留言 (0)