
記住我
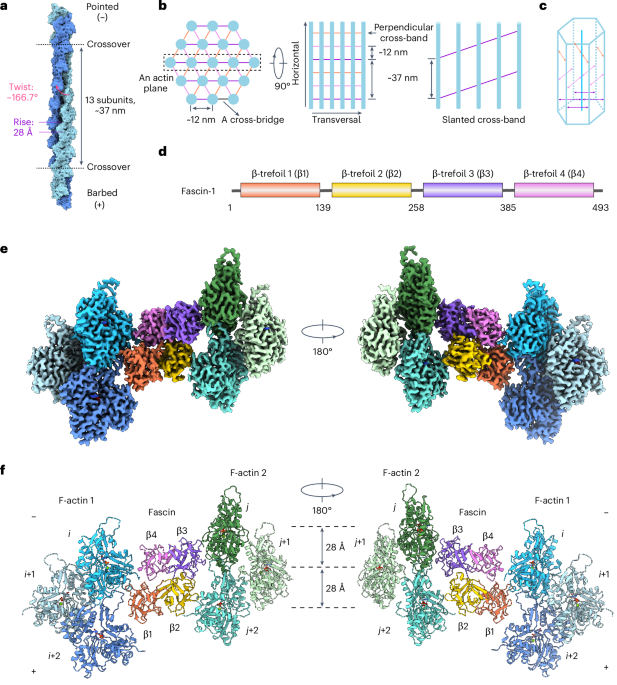

The coding sequence of human fascin-1 (GenBank, NM_003088.4) was obtained from Addgene (cat. no. 31207) and subsequently inserted into a pGEX-6p-1 vector (Cytiva, cat. no. 28954648) using Gibson Assembly (New England Biolabs, cat. no. E2611L) with the following primers: forward, 5′-CTGTTCCAGGGGCCCCTGGGATCCATGACCGCCAACGGCACAGC-3′, reverse, 5′-AGTCACGATGCGGCCGCTCGAGCTAGTACTCCCAGAGCGAGG-3′. The resulting construct was expressed in Escherichia coli BL21(DE3) cells (New England Biolabs, cat. no. C2527H) at 18 °C for 16 h after induction with 0.2 mM IPTG (GoldBio, cat. no. I2481C100). The cells were resuspended and disrupted in lysis buffer (25 mM Tris–HCl pH 8.0, 150 mM NaCl). After centrifugation at 48,000g for 30 min in a JA-25.50 rotor (Beckman Coulter), glutathione S-transferase (GST)-tagged fascin in the supernatant was enriched using GST affinity beads (Glutathione Sepharose 4 Fast Flow, Cytiva). The GST tag was removed by incubating the beads with 3C protease66 at 4 °C overnight. The cleaved target protein was purified on a HiTrap Q HP ion exchange column (Cytiva) and further polished on a Superdex 200 10/300 Increase column (Cytiva) equilibrated with 25 mM Tris–HCl pH 8.0, 150 mM NaCl and 3 mM DTT.
Chicken skeletal muscle actin was purified as previously described67 and stored at 4 °C in G-Ca buffer (2 mM Tris–HCl pH 8.0, 0.5 mM DTT, 0.1 mM CaCl2, 0.2 mM ATP, 0.01% NaN3). F-actin was polymerized at room temperature for 1 h by mixing 5 μM monomeric actin in G-Mg buffer (2 mM Tris–HCl pH 8.0, 0.5 mM DTT, 0.2 mM ATP, 0.1 mM MgCl2) with KMEI buffer (50 mM KCl, 1 mM MgCl2, 1 mM EGTA, 10 mM imidazole pH 7.0) supplemented with 0.01% Nonidet P40 substitute (Roche). The polymerized F-actin was stored at 4 °C overnight before grid preparation.
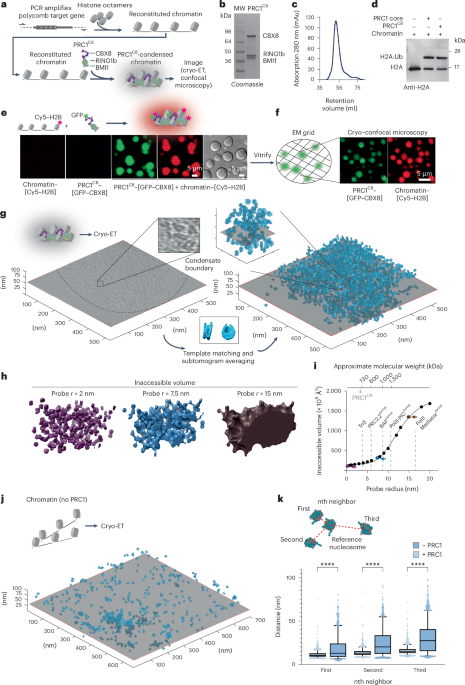
Cryo-electron microscopy and cryo-electron tomography sample preparationTo prepare fascin cross-linked F-actin bundle for cryo-EM analysis, 0.6 μM F-actin was incubated with fascin at a molar ratio of 1:2 at room temperature for 30 min. For cryo-ET analysis, F-actin bundles were prepared by incubating 0.3 μM each of F-actin and fascin at room temperature for 72 h. Then 4 μl of the specimen was applied to a freshly plasma cleaned (Gatan Solarus, H2/O2 mixture) C-flat 1.2/1.3 holey carbon Au 300 mesh grid (Electron Microscopy Sciences) in a Leica EM GP plunge freezer operating at 25 °C and 100% humidity. After 1 min of incubation, the grid was blotted from the back with a Whatman no. 5 filter paper for 4 s and plunge-frozen in liquid ethane cooled by liquid nitrogen.
Cryo-electron microscopy and cryo-electron tomography data acquisitionCryo-EM data were collected on a Titan Krios microscope (Thermo Fisher), operating at 300 kV and equipped with a Gatan K2-summit detector in super-resolution mode using SerialEM68. Frame sequences (videos) were recorded at a nominal magnification of ×29,000, corresponding to a calibrated pixel size of 1.03 Å at the specimen level (super-resolution pixel size of 0.515 Å per pixel). Each exposure was fractionated across 40 frames with a total electron dose of 61 e−/Å2 (1.53 e−/Å2 per frame) and a total exposure time of 10 s. The defocus values ranged from −0.8 to −2.2 μm. To alleviate the effects of potential preferential orientation of fascin within F-actin bundles, data were collected with the stage tilted at three different angles: 30°, 15° and 0°. At 30° and 15°, exposures were acquired by targeting a single hole per stage translation. At 0°, exposures were acquired using the beam tilt/image shift strategy, targeting nine holes per stage translation. A total of 6,288 micrographs were collected, with 1,258, 1,439 and 3,591 micrographs obtained at 30°, 15° and 0°, respectively.
Cryo-ET data were collected on a spherical-aberration (Cs) corrected Titan Krios microscope operating at 300 kV, equipped with a Gatan K3 detector and BioQuantum energy filter (slit width 20 eV). Tilt series were recorded from −60° to 60° with a tilt increment of 3°. The image stack at each tilt angle was acquired at a nominal magnification of ×26,000 corresponding to a calibrated pixel size of 2.6 Å at the specimen level (super-resolution pixel size of 1.3 Å per pixel). Each stack was fractionated into 12 frames with a total electron dose of 2.66 e−/Å2 (0.22 e−/Å2 per frame) and a total exposure time of 0.6 s. All 27 tilt series were collected at a nominal defocus of −4 μm.
Cryo-electron microscopy image processingFrame sequences were motion corrected, dose weighted and summed with 2 × 2 binning (to a 1.03 Å pixel size) using MotionCor2 (ref. 69). Contrast transfer function (CTF) estimation was performed with CTFFIND4 (ref. 70) using nondose weighted sums. Particle picking was performed using a previously reported neural network-based approach we developed for handling F-actin bundles65. Briefly, PDB 7R8V was expanded along its helical axis to produce a filament with 49 protomers and converted to a volume using the molmap command in Chimera71. Synthetic data were procedurally generated by spawning bundle pairs from two of copies of this volume featuring a horizontal displacement randomly sampled from a Gaussian distribution centered at 113.8 Å with an s.d. of 15.9 Å (estimated by manually measuring the interfilament distances of 36 bundles in our dataset) and a vertical displacement randomly sampled from a uniform distribution between −180 and +180 Å. A random angular skew sampled from a Gaussian distribution with a mean of 0° and an s.d. of 1.5° was also applied to the second filament.
To generate synthetic images, none, one, two or three of these bundle pairs were loaded while applying a translation in the x, y and z dimensions, each randomly sampled from a uniform distribution between −250 and +250 Å. This composite volume was projected along the z dimension to generate a noiseless projection. The first (rot) and third (psi) Euler angles were randomly sampled from a uniform circle in degree increments and the second (tilt) Euler angle was randomly sampled from a Gaussian distribution with a mean of 0° and an s.d. of 7.5°. These projections were corrupted by a theoretical CTF and pink noise using EMAN2 functions72. To generate training data for semantic segmentation, the noiseless projections were lowpass-filtered to 40 Å, a binarization threshold of 0.9 was applied and morphological closing was performed by first dilating by 66 Å then eroding by 33 Å. A denoising autoencoder neural network featuring the same architecture as previously described65 was trained using 150,000 noisy and noiseless projection pairs and a 90:10 training to validation split with a learning rate of 0.00005 until convergence after 14 epochs with a validation cross-correlation coefficient loss of 0.9296. This pretrained network was then trained for semantic segmentation using 50,000 projection pairs and a 90:10 training to validation split with a learning rate of 0.00005 until convergence after 13 epochs with a validation categorical cross-entropy of 0.787. The trained neural network for semantic segmentation was used for inference on the experimental images to identify pixels containing F-actin bundles. Tiles of 192 pixels spaced in 48-pixel increments from micrographs binned by 4 were passed through the neural network for semantic segmentation before being stitched back together via a maximum intensity operation. From these semantic maps, particle picks were generated by removing overlap within a 60 Å distance.
The coordinates of 3,056,360 picked particles were imported into RELION v.4.0 (ref. 73) and extracted with a box size of 448 pixels, then binned by 2. The extracted particles were imported into cryoSPARC v.4.2 (ref. 74) for reference-free two-dimensional classification. A subset of 2,231,774 particles were selected from classes featuring parallel actin filaments. A quarter of them (550,000) were used for ab initio 3D reconstruction to generate an initial reference. This reconstruction featured seven filaments arranged in a hexagonal lattice with several apparent interfilament densities corresponding to fascin cross-bridges (Extended Data Fig. 1a). The full selected set of 2,231,774 particles were then subjected to masked homogeneous refinement using a cylinder-shaped mask covering all 7 filaments. The aligned particles were subsequently reimported into RELION for 3D classification without image alignment. Particles from the class displaying the clearest hexagonal arrangement were selected, recentered on the fascin cross-bridge with the strongest density, re-extracted and reconstructed. An ‘H’-shaped mask covering the recentered fascin cross-bridge and its two associated filaments was created. The recentered particles were then subjected to masked 3D classification with a global search range for rot angles and a 10° local search range for psi and tilt angles. Particles with clear cross-bridge density were selected and subjected to another round of focused 3D classification with a mask covering the fascin cross-bridge and its two interacting actin subunits on each filament. Classes with improved fascin density were pooled and refined using the ‘H’-shaped mask, yielding a density map displaying strong fascin density in the mask and four neighboring weak fascin densities.
Particles were subjected to symmetry expansion with recentering on each of the additional fascin densities, followed by focused 3D classification. Particles corresponding to classes with strong cross-bridge density were then re-extracted without binning at a pixel size of 1.03 Å. After duplicate removal, CTF refinement, Bayesian polishing, 3D refinement and postprocessing, we obtained a consensus 3.4-Å resolution density map featuring streaking artifacts in fascin (Extended Data Fig. 1). To further improve the map quality, two rounds of multibody refinement were performed, alternately masking fascin and one filament as one body and the other filament as the other body (that is, F-actin 1 + fascin as body 1 and F-actin 2 as body 2; then F-actin 2 + fascin as body 1 and F-actin 1 as body 2). This greatly improved the density of each F-actin–fascin interface when it was in the body 1 mask. The final 3D refinements yielded postprocessed density maps with resolutions of 3.1 and 3.0 Å, respectively.
To generate a composite map containing both well-resolved fascin–F-actin interfaces, the two multibody maps were aligned on fascin. The poorly resolved pair of β-trefoil domains from each map was removed with the ‘split map’ command, and the remaining well-resolved density from the two maps was stitched together using the ‘vop maximum’ command, both in UCSF Chimera71.
To resolve a fascin cross-linked F-actin hexagonal bundle element, the original 3,056,360 picked particles were extracted in RELION using a box size of 448 pixels and binned by 8. Particles were imported into CryoSPARC and aligned by homogeneous refinement with a cylinder-shaped mask and a hexagonal bundle reference generated from the abovementioned ab initio 3D reconstruction. Aligned particles were subjected to several rounds of focused 3D classification in RELION to enrich particles with strong densities for all possible fascin molecules and F-actin filaments in a hexagonal bundle element. The 8,477 particles contributing to the class featuring the best density were re-extracted with a box size of 448 pixels and no binning, as well as a box size of 720 pixels binned by 2. Both particle stacks were reconstructed and refined, generating postprocessed density maps at 8.7 and 12.0 Å resolution, respectively. Four additional classes were refined that featured high-quality density for the central filament and discernable density for its six bound fascins, but varied quality for the peripheral filaments and cross-bridges.
Local resolution estimation was performed using the procedure implemented in RELION.
Measuring hexagonal bundle element filament parametersDensities corresponding to the seven actin filaments in the sub-nanometer resolution 460-Å bundle reconstruction were segmented using Chimera. The rotational phase offsets between each side filament and the central filament were measured by individually fitting the density of each side filament into the central filament using Chimera’s ‘Fit in Map’ tool. The consensus rise and twist of each filament was measured using both the relion_helix_toolbox75 in RELION v.4.0 and the iterative helical real space reconstruction program hsearch_lorentz76. To prepare the filaments for analysis with these programs, the segmented filament volumes were resampled such that the filament was centered and aligned along the proper axis. The programs were seeded with initial rise and twist estimates of 27.9 Å and −166.0°, respectively. For local twist measurements, atomic models of actin subunits (PDB 7R8V) were individually rigid-body fit into each filament density to generate a docking model, which was passed as input to an updated version of a custom script that we recently reported58.
Equivalent docking models generated from the centered and aligned volumes were furthermore used to perform axial shift measurements. The Cα of H plug residue I266 was selected as a fiducial, and the x, y, z coordinates of this atom in the central subunit of each model were recorded. Axial shift was calculated as the difference in this atom’s z coordinate between each of the side filaments and the central filament, equivalent to displacement along the filament axis.
Tilt series reconstructionMotion correction of individual frames and CTF estimation of combined tilt stacks were performed in WARP77. Data were exported for subsequent alignment and reconstruction using the IMOD software package78. Tilt series were binned by 3 to a final pixel size of 7.8 Å, aligned with the patch-tracking approach, then reconstructed using weighted back-projection with a SIRT-like filter equivalent to 12 iterations.
Synthetic tomography data generationSynthetic datasets approximating subtomograms were generated to train neural networks for denoising and semantic segmentation. The training datasets for the neural networks consisted of noisy source volumes simulating empirical data, along with paired target volumes of in silico-generated ground truth fascin–F-actin bundles. To produce these volumes, the seven-filament bundle element reconstruction was laterally expanded to produce a larger hypothetical lattice of 19 filaments bound by 84 fascins. Atomic models of F-actin (PDB 7R8V) and fascin cross-bridges were fit into this extended volume, and converted to volume data using the molmap command in Chimera. These docked components were used to generate plausible regions of bundles with varying architectures. For each simulated region, a random subset of the F-actin volumes was loaded. Fascins were loaded only if they were in a position to bind these filaments; to simulate sub-stoichiometric binding, a random subset of these fascins were rejected. All of the F-actin volumes were combined in a single volume, all of the fascin volumes were combined in a separate volume and both volumes were saved. A library of 200 volume pairs was procedurally generated using this approach.
The library was used to generate a dataset of 128 voxel training volumes at a sampling of 7.8 Å per voxel. Paired fascin and F-actin volumes were rotated about the rot and psi angles by random, uniformly sampled values between 0° and 359°, whereas the tilt angle was randomly sampled from a Gaussian probability distribution centered at 90° with an s.d. of 20°. The density was translated in the box along each dimension by a random uniform translation in the range of ±195 Å.
A set of 20,000 synthetic noisy volumes were generated by first projecting the repositioned volume at fixed angles corresponding to the experimental tilt series collection scheme (−60° to +60° with a 3° increment). These projections were corrupted by the CTF, and reconstructed back into a 3D volume using reconstructor class functions as implemented in the EMAN2 python package72. Empirical noise was extracted from each of our experimental tomograms by computing the average Fourier transform of 50 randomly sampled 128 voxel boxes. For each synthetic volume, one of these empirical noise boxes was randomly selected, multiplied element-wise by a white noise box of the same dimensions, normalized and scaled by a random scale factor that modulated the signal-to-noise ratio of each synthetic particle. The synthetic volume was then summed with its noise volume in Fourier space. To account for interpolation artifacts from CTF application or noise addition in Fourier space, the synthetic volumes were cropped to 64 voxels in real space. Each noisy particle had a corresponding noiseless ground truth particle, as well as binarized semantic maps calculated from the paired F-actin and fascin volumes.
Neural network training for tomogram denoising and semantic segmentationPretraining of a denoising autoencoder consisting of 3D convolutional layers in a U-net architecture (Extended Data Fig. 5) was performed with a single NVIDIA A100 graphics processing unit (GPU) with 80 GB of video RAM, using a learning rate of 0.0001. Training was run on the 20,000 pairs of noisy and ground truth volumes with a 90:10 training to validation split, until the network converged with a cross-correlation coefficient validation loss of 0.8659 after 20 epochs. Initial inference testing on experimental tomograms resulted in distorted denoised volumes. We reasoned that this was because the synthetic data did not model corruption of the experimental data with sufficient fidelity. We therefore continued to train the network using a domain adversarial neural network approach79. An additional network head was added for domain classification by forking the network output after the feature extraction layers (Extended Data Fig. 5a). The domain classification head consists of a gradient reversal layer and additional 3D convolutional layers, which are followed by flattening to a dense layer, then a binary classification layer with a sigmoid activation function. One hundred synthetic volumes and 100 volumes extracted from the experimental tomograms were used as the training set for the domain classifier. Adversarial training was performed by alternatively passing these data through the domain classifier head, followed by retraining the feature extractors with the denoising head using only the 100 synthetic volumes. This adversarial training was run for ten iterations; however, after multiple iterations, too many high-resolution details were lost, so the first iteration of adversarial training was used for denoising.
After using a domain adversarial neural network to complete training of the denoising autoencoder, a semantic segmentation network was trained using 10,000 volume sets and the autoencoder’s pretrained weights. The final layer was adapted to produce multichannel outputs, featuring a softmax activation layer and random initial weights. After training for ten iterations with a learning rate of 0.00001, the categorical cross-entropy loss of the semantic segmentation network was 0.0556.
Tomogram denoising and semantic segmentationThe trained neural networks were used to denoise and semantically segment empirical tomograms, once again using single A100 GPUs. The tomograms were extracted into 64-voxel tiles sampled every 32 voxels in x and y and every 4 voxels in z, normalized and passed as inputs to the neural network. The network outputs were masked with a 48-voxel cubic mask to minimize edge artifacts and stitched together via maximum intensity projection. Each input tomogram produced three outputs: a denoised tomogram, a semantic map of actin filaments and a semantic map of fascin molecules.
Measurement of F-actin rotational phase offsets in denoised tomogramsThe denoised tomograms were manually inspected to identify bundles with high-quality density, straight filaments and uniform fascin cross-bands spanning multiple crossovers. Subsequently, a 17-subunit F-actin model was generated from PDB 7R8V and fit into filament densities in the bundle using Chimera while maintaining identical axial register of the filaments across the bundle. Synthetic density maps were then generated for each fitted F-actin atomic model using the ‘molmap’ command in Chimera with a resolution cutoff of 6 Å. The rotational phase offset was measured by fitting the synthetic density map of the rotating filament to the reference filament using Chimera.
Subtomogram averagingSubtomogram particle picking required minimal postprocessing of the fascin semantic maps. A threshold of 0.9 was applied to each semantic map, and the centroids of objects larger than 50 voxels were designated as potential fascin picks. The center of the hole in the carbon film for each tomogram was manually picked, and potential fascin picks within a 740-voxel radius of the hole center were retained as picks. Subtomogram averaging was performed using RELION v.4.0. From 26 tomograms, 135,000 192-voxel pseudo-subtomograms were extracted at bin 3 (voxel size 7.8 Å) and cropped to a 64-voxel box. A small subset of 1,000 pseudo-subtomograms was used for initial model generation with three classes. Two of the three classes contained 82% of the particles, and the most populous class was selected. An initial global alignment of particles was performed using Class3D with one class. A subset of 5,000 particles with tilt angles outside the range 60–120° were subsequently excluded. The rot angle was then removed from the metadata, and a bimodal psi prior was used for subsequent 3D auto-refinement. An H-shaped mask (85% z-length) was used for focused 3D auto-refinement and subsequent processing of the central filaments and fascin bridge. Three-dimensional classification using two classes was attempted to remove partially decorated fascin monomers or picks of single filaments, but both classes were essentially identical, suggesting a minimal false-positive rate in picking.
After another round of local 3D auto-refinement, pseudo-subtomograms were sequentially re-extracted and subjected to local 3D auto-refinement at bin 2, then bin 1. This produced a reconstruction with a nominal resolution of 8.5 Å, yet the map appeared distorted. After two rounds of CTF refinement, FrameAlignTomo and local 3D auto-refinement, a final undistorted 6.7-Å reconstruction was obtained. Multibody refinement was conducted in an identical manner as described above for the single-particle analysis. Local resolution estimation was performed using RELION’s implementation.
Bundle hierarchical clustering based on subtomogram variability analysisPer-subtomogram deviations from the consensus reconstruction were measured using the Mahalanobis distance of each particle’s multibody refinement amplitude along the eigenvectors corresponding to rotations. To calculate filament interface scores, fascin cross-bridges were assigned to interfilament interfaces using a custom script. The filament interface score (FIS) was computed as:
where N is the number of fascin cross-bridges along the interface, L is the length of the interface and \(\bar\) is the average energy (Mahalanobis distance) of the cross-bridges along the interface. Graphs were constructed from 24 well-ordered bundle regions approximately 400 nm in length, where actin filaments correspond to nodes, and nodes are joined by an edge if the corresponding actin filaments were bridged by at least one fascin in the region. Edge weights were assigned as the filament interface scores.
Hierarchical clustering was performed using the ward linkage method, and the maximum distance was normalized across all graphs to enable direct comparisons. The metrics of transitivity, modularity and minimum, average and maximum cluster sizes were computed as a function of linkage distance for each graph.
Atomic model building, refinement and analysisPreviously determined structures of actin (PDB 7R8V)67 and fascin (PDB 3LLP)27 were rigid-body docked into each of the high-resolution multibody refinement-derived density maps in ChimeraX v.1.6.1 (ref. 80), then flexibly fitted with ISOLDE81. Atomic models were refined using phenix.real_space_refine82 alternating with manual adjustments in Coot83. Refined models were rigid-body fit into the composite map, split at the boundary between the well-resolved and poorly resolved β-trefoil domains in the multibody map from which the model was derived, merged and real-spaced refined. Model validation was conducted with MolProbity84 as implemented in Phenix. The structural rearrangements of fascin in different states were analyzed with the DynDom online server85.
Computational modelBased on the cryo-EM multibody refinement analysis (Fig. 5a), a computational model was developed to predict the 3D arrangement of F-actin arrays cross-linked by fascin. The code was implemented in MATLAB. This model iteratively adjusts the angular shift of each F-actin in the array to find the configuration corresponding to the maximum probability of forming fascin cross-bridges (Extended Data Fig. 4a), which is inferred from an energy function linked to cross-bridge geometry (Extended Data Fig. 4b). The angular shift between a pair of filaments determines the physically plausible fascin bridging positions. The favorability of a bridging position is furthermore linked to the extent a fascin cross-bridge must be deformed to accommodate the local binding geometry, as visualized in the multibody analysis. Both of these physical phenomena are simultaneously represented in the energy function, where distances between fiducial actin residues spanning the fascin–actin contact surface serve as a proxy for the local interface geometry (Extended Data Fig. 4b).
Derivation of geometric rulesFour pairwise distances between fiducial residues in fascin–actin contact sites were used to estimate cross-bridging energies: R95i and R95j+1, R95i and S350j+3, D24i+2 and S350j+3, and D24i+2 and R95j+1 (Extended Data Fig. 4b). The model assumes that the distances between the fiducial residues are independent. In addition, the distance between the center of geometry of these residues and the plane crossing the filament axes was incorporated to prevent unrealistic fascin distortions.
The distance-dependent cross-linking probability Pn(d) for each distance n (where n = 1, 2, …, 5), was calculated as follows:
$$_\left(d\right)=}^\right)}^}$$
Here, μ and σ represent the mean and s.d. of d, respectively. These parameters are calculated from multibody refinement analysis and are reported in Extended Data Table 2. Parameter m is the multiplication factor for the s.d. σ, which encodes cross-bridge plasticity. The total cross-bridging probability P is given by the product of the five Pn:
$$P=_\times_\times_\times_\times_$$
Distributions of these probabilities as a function of d are presented in Extended Data Fig. 4c,d. The normalized cross-bridge energy was calculated as:
Algorithm of growing bundlesEach filament in the model is identified by the angle that the first monomer forms with the origin of the domain. Filaments are represented as double-stranded helical structures composed of 14–19 monomers, with each monomer rotated by −166.7° and translated by 2.75 nm along the vertical axis.
The algorithm begins by placing the first filament (filament A) at a random position in the bundle and assigning it a random absolute rotation angle θA. In the first iteration, a second filament (filament B) is positioned 12.15 nm from filament A. The algorithm evaluates the probability P of forming a cross-bridge, P, for each possible rotation angle θB of filament B, which ranges from 0 to 359.9° in increments of 0.1°.
For each value of θB the probability P of forming a cross-bridge between filament A and filament B is calculated. This involves evaluating the probability for all pairs of protomers, specifically (i and i + 2) from filament A and (j + 1 and j + 3) from filament B. The probability P is defined as the product of five distance-dependent probabilities P1, P2, P3, P4 and P5, which are based on the distances between specific fiducial residues.
The algorithm then selects the rotation angle θB that maximizes the cross-bridge formation probability P, considering only angles where P exceeds a minimum threshold τ (the minimum threshold for fascin binding). Once filament B is added, the algorithm proceeds to add additional filaments. Each new filament is positioned within 12.15 nm of at least two existing filaments in the bundle. This process continues iteratively, with the algorithm optimizing the rotation angles and positions of new filaments to maximize the likelihood of cross-bridge formation by fascin.
Model parameterization based on sensitivity analysisModel parameterization involved adjusting two free parameters, m and τ, based on their effects on the distribution of P (the overall probability of forming a cross-bridge) and on the fraction of filament pairs that formed a fascin cross-bridge. For this analysis, filament pairs with randomized angular shifts were used. First, we evaluated the effect of m on the cross-bridge probability distribution. We tested how systematic variations in m affected the distribution of P. For each m value between 1 and 7 (with a step size of 1), and at each value of τ from 0.1 to 0.8 (with a step size of 0.1), 50 independent runs were performed, resulting in a total of 2,800 runs. The normalized cross-bridge energy distribution exhibited a biphasic relation with m: it increased as m increased from 1 to 4, and then decreased for m > 4 (Extended Data Fig. 4e). Next, we examined how systematically varying m affected the fraction of cross-bridges formed across runs. We observed a low fraction of cross-bridges for m < 4, with an average cross-bridge fraction <0.5 and a median fraction of 0 (Extended Data Fig. 4f). Based on these data, we identified m = 4 as the value that captured the variability in cross-linking probabilities and resulted in a high enough fraction of fascin bonds (Extended Data Fig. 4e,f).
The sensitivity of the model outputs was also assessed for τ. For these simulations, m was fixed at 4 as τ was varied between 0.1 and 0.8 (with a step size of 0.1). For each τ step, 50 model runs with randomized angular shifts were performed, totaling 400 independent runs. We found that for τ > 0.6, Ec = 0, indicating that τ was too restrictive to allow cross-bridges to form (Extended Data Fig. 4g). For τ ≤ 0.6, the distribution of Ec broadened, providing a range (0.1 ≤ τ ≤ 0.6) that produced fascin cross-bridging (Extended Data Fig. 4g). Further analysis of the mean and median cross-bridging fraction revealed that using τ < 0.4 was too permissive, allowing constitutive bond formation (Extended Data Fig. 4h). Therefore, the acceptable range for τ was identified as 0.4 ≤ τ ≤ 0.6. Based on this range, τ = 0.4 was selected for further simulations of higher-order fascin cross-linked assemblies.
Evaluating the effect of F-actin angular shift on cross-bridge energyWe analyzed pairs of filaments composed of 14 protomers (spanning a single crossover) to evaluate the effect of relative angular shifts on the probability of fascin cross-bridging, indicated as normalized energy, Ec. Systematic variation of the two filaments’ rotation angles between 0° and 180° identify regions without cross-bridging, corresponding to Ec = 0, and regions of cross-bridging with values of normalized Ec < 0 (Extended Data Fig. 4i). Consistent with the periodicity of F-actin, we observed a regular pattern of teardrop-shaped energy wells around minima (indicated by white dots in Extended Data Fig. 4j). To explore the implications of this pattern, we examined Ec with the absolute rotation angle of filament A held constant at 102.7° while rotating filament B between 60° and 140°. This revealed one global minimum in Ec and several local minima (Extended Data Fig. 4j). In this case, the global minimum corresponded to a fascin bridge in the ‘down’ pose, whereas the most favorable local minimum corresponded to fascin bridge in the ‘up’ pose at a nearly equivalent position (Extended Data Fig. 4k). This suggests that the two teardrops that intersect at a given filament A angle correspond to nearly isoenergetic ‘up’ and ‘down’ binding positions, with the relative favorability modulated by subtle changes to the interfilament rotation angle. To test the hypothesis of isoenergetic binding positions, we examined all minima in angular shift ranges that satisfy τ to form fascin bonds (yellow in Extended Data Fig. 4l). Consistently, when the rotation angle of filament A is greater than that of filament B (upper diagonal in Extended Data Fig. 4i), the Ec minima correspond to ‘down’ pose bridges (Extended Data Fig. 4m), whereas when the rotation angle of filament B is greater than that of filament A (lower diagonal in Extended Data Fig. 4i), the Ec minima correspond to ‘up’ pose bridges (Extended Data Fig. 4n), with adjacent minima corresponding to bridging of adjacent axially shifted protomers.
Graphics and additional analysisFigures and videos were generated with UCSF Chimera and ChimeraX. All statistical analysis and plotting were performed in GraphPad Prism. Custom code for tomogram denoising and downstream analysis was generated with the assistance of ChatGPT v.4.0.
Reporting summaryFurther information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
留言 (0)