
記住我








In recent years, abundant literature has been dealing with the complex and mutual connections of sleep and epilepsy. Those astonishingly firm links infiltrate each layer from clinical to network and cellular; suggesting an intrinsic relationship [1]. The distribution of seizures and interictal acivity occurring during the 24 h cycle is patient-dependent suggesting the lack of a universal rule [2]. However, the timing and intensity of ictal and especially interictal epileptic phenomena seem to follow the course of sleep homeostasis.
Some basic terms and concepts underlying the complex links of sleep and epilepsy:
Homeostatic regulation has major role. This general restorative force acting on all levels of body functioning provides multilevel control of sleep (sleep homeostasis). It maintains synaptic capacity—strength and number of synapses—closely related to plasticity and learning [3, 4].
Sleep homeostasis operates on sleep macro-levels e.g. sleepiness (high sleep pressure) and sleep ciclicity. It also acts on micro-levels, represented by sleep microstructure including the dynamics of sleep slow waves (SWs) within the framework of the cyclic alternating pattern (CAP) [5,6,7]
NREM sleep-oscillations range from high-frequency (80–200 Hz) hippocampal and cortical ripples (high frequency oscillations; HFO) through thalamo-cortical sleep spindles (12–15 Hz) and cortical (< 1Hz) SWs. SWs are constituted by active "up" states associating with spindles and ripples and disfacilitated "down" states [8]. The high-frequency neuronal firing including spindles and HFO favor synaptic normalization and is modulated by homeostatic forces as well.
SW, sleep spindles and ripples coalesce during NREM sleep, SW carrying an orchestrating role [9,10,11,12]. Thalamocortical SWs and sleep spindles [13] cooperate in memory processing and other cognitive functions [14, 15]. This is evidenced in multiple ways both in animals and humans [16,17,18,19,20,21].
The fine-tuned cooperation of hippocampal and cortical HFO with spindles, provides additional communication between the hippocampus and cortical regions for learning and memory processing [2, 19]
Slow wave (< l Hz) activity (SWA) is an inherent feature of NREM sleep, distinct from the focal slowing produced by brain lesions [22]. According to the synaptic homeostatic hypothesis, SW power increases during the day reaching its peak at the beginning of the “descending”- i.e. deepening—slope of the first sleep-cycle of the night [6, 7]. SWA parallels use-dependent sleep homeostasis [23].
The first sleep cycle of the night (about the first 60 min) is characterized by high homeostatic and sleep pressure; [23] reflected by augmented SWA. Then a decrease of the homeostatic pressure and SWA follows overnight. SWA is the product of the thalamocortical system; it has been proposed to be critical for synaptic homeostasis, metabolic regulation, and metabolic waste-clearance [14, 24].
A significant function of NREM sleep is recalibrating synaptic strength and numbers (termed synaptic upgrading or downgrading). NREM sleep modifies synaptic connections by different plasticity rules; [25], thus renewing functional capacity for the next day [26]. Also this function is marked by slow wave activity. The synaptic homeostasis hypothesis [6, 7] suggests an upgrading of synaptic strength (also termed upscaling) during the day and its downgrading (or downscaling) with deep nREM sleep. An opposite terminology (but apparently the same tendency) is proposed by Timofeev et al. [27], supposing the downgrading (also termed downscaling; weakening) of synapses during the day, and the restoration of synaptic strength (upgrading) during NREM sleep. Both curves are determined by homeostatic power.
Synaptic plasticity is the ability of neurons to modify the strength of their connections (efficacy). It is supposed to be a key substrate of learning and the development of neural circuits. Potentiated synapses are more excitable and may drive the system to a state of abnormal hyper-excitability—epilepsy—if not kept under control.
Synaptic plasticity comprises two opposing processes such as
the rapid acting anti-homeostatic positive feedback named ‘Hebbian’ after D.O. Hebb; strengthening active synapses by long-term potentiation (LTP) and diminishing less active ones by long-term depression (LTD) [28]; and
a slower acting homeostatic negative feedback mechanism - the actual synaptic homeostasis - proposed to balance the destabilizing influence of the Hebbian LTP and LTD. It protects neurons from becoming hyper-excitable or silent; it keeps synaptic strength at an appropriate level [29, 30], thus maintaining the functional capacity of neurons and networks for functioning.
The fine-tuned and flexible balance of those two types - Hebbian and homeostatic - of plasticity - means that any manipulation of neurons would induce a mixture of plasticity including local and global, Hebbian and homeostatic processes. The fine molecular mechanisms related to the activity-dependent adaptive cooperation of the two mechanisms have been studied recently [31].
Epilepsy and synaptic plasticity. The Hebbian and homeostatic plasticity are in flexible balance. Normally, any over-excitation or silence of neurons is immediately compensated by the synaptic homeostasis, ensuring the ideal, pre-set level of synaptic capacity. A failure of this balance in any direction is dysfunctional; if over-excitation dominates, an epileptic derailment may evolve [24, 32,33,34,35].
The ‘epilepsy homeostasis hypothesis’ [22], is one of the consequences of this theory on synaptic balance: after a seizure (necessarily representing extreme synaptic over-excitation), homeostatically driven slow waves follow immediately even in wakefulness; reducing and normalizing the aberrant hyper-activity, at the price of transient cognitive impairment.
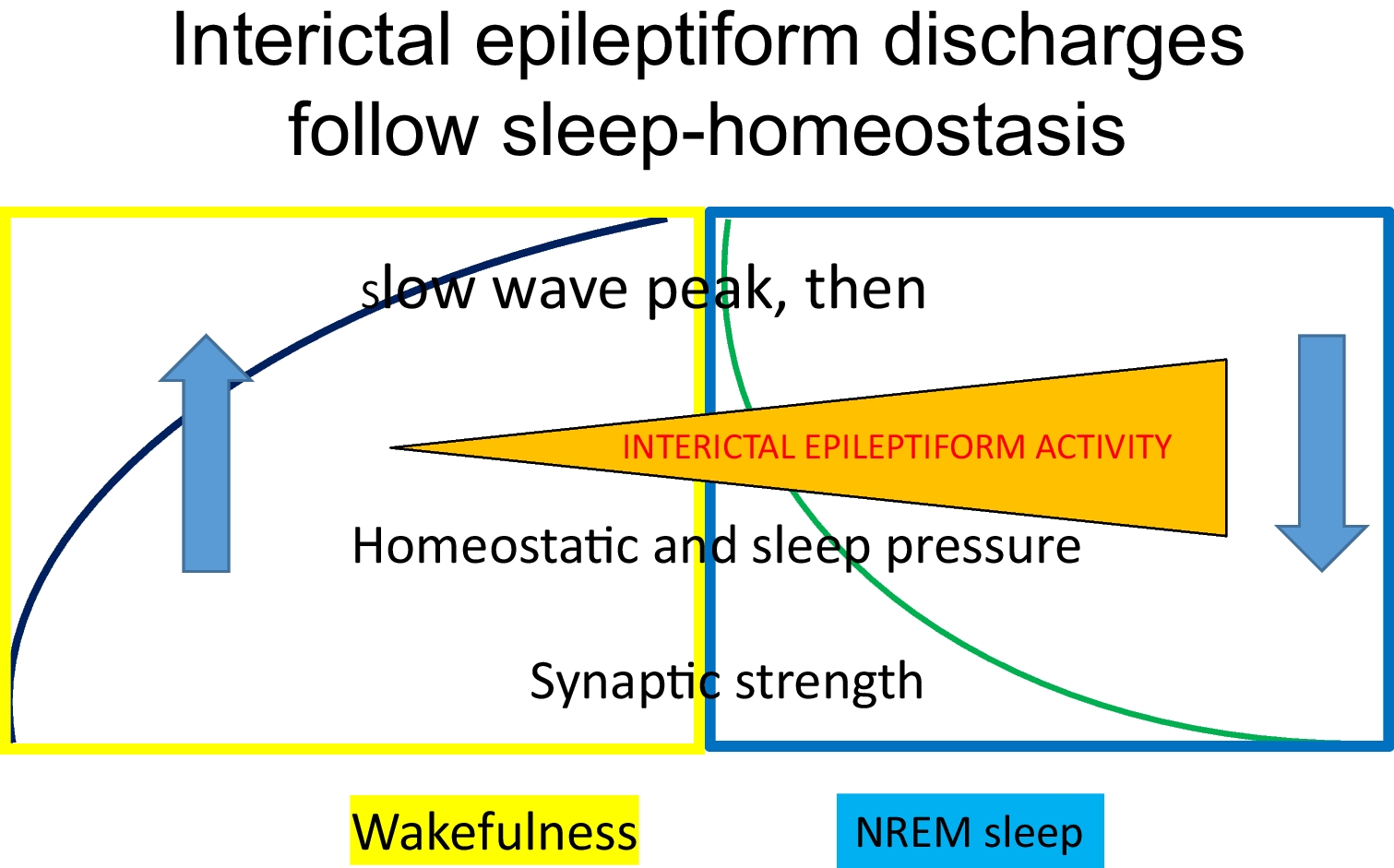
Seizures Promoted or Influenced By SleepThe seizure-provoking effect of sleep deprivation is well known; it is best seen in so-called idiopathic generalized epilepsies [36, 37]. Any sleep-disturbing factor or condition e.g.shift working, partying, obstructive sleep apnea, restless legs syndrome or pain-conditions, may increase the frequency of seizures This effect might be related to the evolving augmentation of homeostatic pressure, resulting in an increase of SWs that facilitate epileptic activity [38]. It is remarkable that paradoxically, the deprivation and occurrence of NREM sleep have the same (albeit different degree) seizure-inducing effect [39]. This may be resulted from the high homeostatic pressure and increased SW power shared by these conditions (Fig. 1).
Fig. 1
Interictal epileptiform discharges follow sleep-homeostasis
A substantial proportion of seizures occur during sleep [40,41,42]. In sleep-related hypermotor epilepsies (SHE) typically originating in the frontal lobes, most seizures evolve in NREM sleep. Those seizures link to frontal SWs and microarousals. There are numerous microarousals -sleep is fragmented- due to augmented arousal forces characterizing SHE [43]. SHE seizures favour phase N2 or N3 NREM sleep in the first sleep-cycles of the night; when sleep-promoting and wake-promoting forces meet.
Absence epilepsy attaches to NREM sleep in another way. Absences occur in a ‘vigilance-window’; in the transition-period from wakefulness to superficial NREM sleep, where homeostatic power prevails. Wakefulness and REM sleep inhibit them. The link of absences to the wakefulness-NREM sleep transitions has been experimentally supported [44]. Therefore, absence epilepsy has been considered the system-epilepsy of sleep promotion [45, 46].
Interictal Epileptic Discharges (IEDs) Strongly Link to NREM SleepIEDs cover > 40% of NREM sleep of children with electrical status epilepticus (ESES) and Landau-Kleffner syndrome [47], and, consistently with the spectral relation of self-limiting focal epilepsies (SeLFEs) with ESES [48], centrotemporal spikes (CTS) accumulate in the sleep of those malignant conditions. At the same time, there are hardly any IEDs in the waking EEGs in either of them. The sleep-accumulation of CTS has been convincingly supported by fMRI [49]. An important sleep-link of interictal epileptic activity has been described also in Lennox-Gastaut syndrome [50] medial temporal lobe epilepsy [51] and in idiopathic generalized epilepsies.
Paralleling the homeostatic pressure, IEDs accumulate in the first sleep cycles and on the deepening slopes of NREM sleep cycles [34, 41, 51]. More IEDs occur early at night-, and less in later periods of sleep [52]; following the homeostatic decay overnight. The effect of homeostatic regulation is seen during the day as well: the rate of IEDs increase from morning to afternoon and evening hours [53].
The link of IEDs to NREM sleep is not continuous. In most epilepsies, IEDs link to NREM sleep SWs (in the frameworks of the cyclic alternating pattern (CAP) A1) [54, 55]; or (in SeLFE), to sleep spindles [55, 56]. Moreover, even slow wave-phases influence the timing of IEDs (including epileptic ripples); normal high frequency oscillations (HFO) link to different phases of SW compared to spikes and pathologic HFO. IEDs prefer the transition-zone from the 'up' to the 'down' state of the SW, suggesting their link with slow wave synchronization rather than with excitability [38, 57].
Normal Sleep Patterns Transform to IEDs, Supporting the Idea That Epilepsies Enroot in NREM SleepIn certain epilepsies, reciprocal relations of IEDs and sleep spindles have been described. IEDs’ numbers anti-correlated with the numbers of sleep spindles and spike-waves (SWs) suggesting an epileptic transformation of normal NREM sleep patterns.
Such epileptic transformation has been early supposed by Gloor [58] in absence epilepsy. They noted that SWs were produced by the same thalamocortical system that normally produced sleep spindles. Beenhakker and Huguenard [59] elaborated how the normal spindle-generating burst-firing mode of the thalamocortial system switched to SW-producion, and recently, this mechanism has been further explored [60].
The transformation of sleep patterns to IEDs was shown also in medial temporal lobe epilepsy. Here, hippocampal sharp wave-ripple complexes (SPW‐Rs) deeply involved in memory consolidation, turn to epileptic spike-pathological ripples during NREM sleep [21, 61, 62]; IEDs’ number increases at the ‘expense’ of temporal sleep spindles [63].
An analogue anti-correlation of centrotemporal spikes (CTS) and sleep spindles has been described in self-limiting focal childhood epilepsy with centrotemporal spikes (SeLECTS; Rolandic epilepsy) [50, 54]. Here, an additional epileptic progress has been noticed: CTS occur alone in non-epileptic children; they are associated with HFO in Rolandic epilepsy; and in cases of progress to electrical status epilepticus in sleep (ESES) or Landau-Kleffner syndrome, the power and amplitudes of the associating HFO augment further [64].
In focal epilepsies, the number of sleep spindles was less than in normal controls, and it was less in the region of the epileptic focus compared to contralateral homologous areas [48]; providing further support to the sleep spindle to spike transformation.
The experimental data on posttraumatic epilepsy [27, 65, 66] showing the transformation of NREM sleep with extreme increase of SW down states (occurring less frequently even in wakefulness and REM sleep as well); confirm the sleep relatedness of epileptic derailment.
REM Sleep Impedes EpilepsyWhile the links of NREM sleep and epilepsies have been extensively studied, much fewer research focused on REM sleep’s protective role.
Compared to other sleep-stages and wakefulness, multiple pathways make REM sleep immune to seizures; occurring just exceptionally during this phase. Therefore, REM sleep has been considered a protecting agent against seizures [67, 68].
The cortical neuronal desynchronization during REM sleep—most marked during phasic REM [69]—may confine epileptic phenomena; reducing their propagation away from the source-generator. Therefore, IEDs appearing in REM sleep have high localizing value [57, 70,71,72]. Orexins may promote seizures, that may be partially attributed to their REM-supressing effect [73].
Due to the ‘antiepileptic’ effect of REM sleep, REM-related spikes have a high localizing value. There are just a few reports of REM sleep providing false localization [74].
We found but one case-study, where IEDs were maximal during REM sleep [75]. Based on a recent study, REM sleep affects the IEDs of mesiotemporal versus extratemporal epilepsies differently [76]
Congruently with REM sleep’s protective effect against IEDs, more seizures occurred during video-EEG monitoring if there were frequent “breakthrough” spikes during REM sleep. In other words, REM-related interictal activity could forecast seizure-risk and epilepsy-severity. [
留言 (0)