
記住我



The first discovered galectin (galectin-1) represents one of the best-studied and well-characterized galectins in both testis and epididymis. Galectin-1 is involved in a variety of cellular processes, including pathogen recognition, selective induction of Th1 and Th17 apoptosis, inhibition of T cell trafficking, expansion of tolerogenic dendritic and regulatory T cells, maintenance of maternal-fetal tolerance, induction of angiogenesis and suppression of autoimmunity [28, 29, 33,34,35,36].
Early reports demonstrated mRNA expression of L14 lectin (identified later as galectin-1) in the adult mouse testis [37]. Subsequent studies analyzing the galectin-1 expression during testis development revealed differential expression patterns in Sertoli cells from embryonic and pre-pubertal testis. During embryonic development before germ cell heterogeneity manifests, a cyclical pattern of galectin-1 expression arises. In the adult testis, Sertoli cells showed a stage-specific cyclical regulation with the highest abundance in spermatogenic stages X-XII and lowest levels at stages VIII-IX [38]. Other studies confirmed the localization in Sertoli cells, but found the protein additionally also in germ and Leydig cells using immunohistochemistry and in testicular extracts by Western blotting [39, 40].
For the rat testis, Dettin et al. confirmed the developmental regulation and stage-specific expression during spermatogenesis for galectin-1 [41]. Here, the testicular concentration of galectin-1 increased in an age-dependent manner in postnatal rats (9–60 days), with similar notions also in porcine and swamp buffalo testis [41,42,43,44]. Interestingly, during spermiation (stage VI – VIII) in rat, galectin-1 was localized to the apical part of Sertoli cells as well as on heads of mature spermatids and residual bodies. Following sperm release to the tubular lumen, galectin-1 localization was seen mainly at the basal part of Sertoli cells from where it spread out to the entire cell during progression of germ cell differentiation [41]. A similar pattern of galectin-1 localization was detected in rat Sertoli cells by Özbek et al., however, slight staining differences were observed in the germ cells. Here, additionally positive spermatocytes and round spermatids were recorded [43]. In agreement, our own data demonstrated that in normal rat testis galectin-1 was mainly found in Sertoli and germ cells [45]. The presence of galectin-1 in human Sertoli and germ cells as well as in peritubular cells was reported [46, 47]. Here, a polarized secretion in cultivated Sertoli cells was evident [46] that could occur via extracellular vesicles [48]. In summation (see Table 1 for overview, Fig. 1), reports indicate that the expression of galectin-1 starts in prenatal testis and increases during puberty, when neo-antigen harboring haploid germ cells first appear, a fact that could point to a role in germ cell development, but also in establishing testicular immune privilege.
Table 1 The table presents the cellular localization of galectin-1 and galectin-3 in mature testis and epididymis across different speciesFig. 1
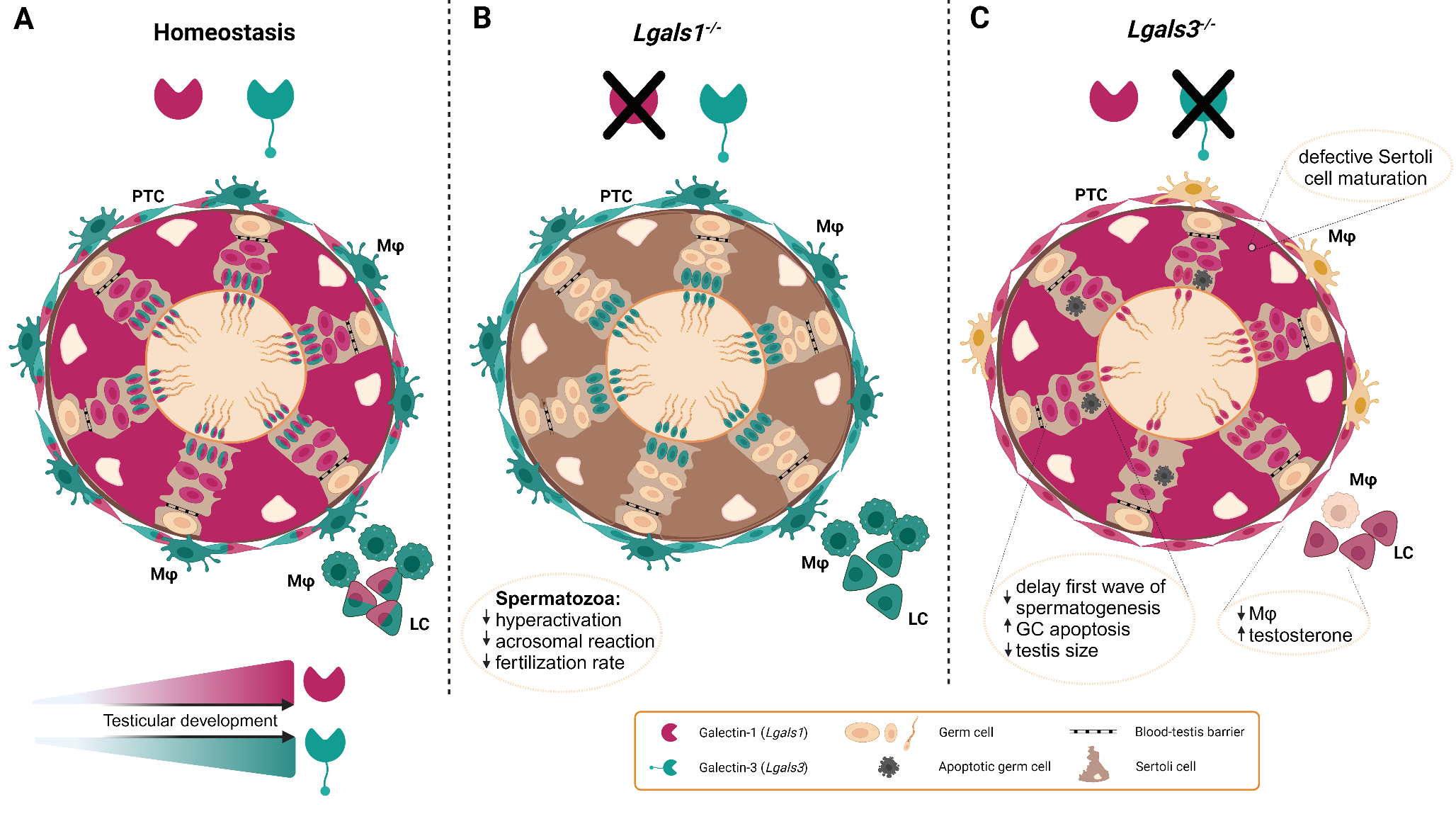
Murine model of testicular galectin-1 and galectin-3 expression patterns and the consequences of Lgals1 and Lgals3depletion on physiological function. (A) Color coded cellular expression of galectin-1 (fuchsia color) and galectin-3 (cyan color) in the mouse testis. Galectin-1 and 3 levels increase in an age-dependent manner in the testis and are expressed by peritubular, Leydig and germ cells with galectin-3 only present in later germ cell stages. Sertoli cells express galectin-1, but not galectin-3. In contrast, macrophages show galectin-3, but not galectin-1. (B) In Lgals1-/- mice, sperm functional parameters are affected as indicated. (C)Lgals3-/- mice are fertile, however, the testis exhibits impairments as noted in the figure. PTC: peritubular cell, LC: Leydig cell, Mφ: macrophage. Image was created using BioRender
The latter is supported by an increasing body of evidence that demonstrates an immunoregulatory function in autoimmune disease models mediated through galectin-1 effects on immune cells [28]. In this context, galectin-1 secreted by murine Sertoli cells co-cultured with dendritic cells has been identified as a factor that induces the differentiation of tolerogenic dendritic cells. This is achieved by decreasing the expression of surface molecules (CD80, CD83, CD86, CCR7, CD11c) crucial for T cell activation and dendritic cell migration to lymph nodes. Furthermore, galectin-1 enhances the secretion of immunoregulatory IL-10 and TGF-β1, while reducing the secretion of pro-inflammatory TNF and IL-12p70 by immature dendritic cells in a dose-dependent manner [49]. By these means, galectin-1 is considered as a crucial mediator of the immunotolerogenic environment created by Sertoli cells in the testis, playing a significant role in maintaining testicular immune privilege. This is well exemplified by the successful application of Sertoli cells in co-transplantation studies (e.g. with pancreatic islets). Additionally, galectin-1 directly inhibits the activation of B and T cells and induces regulatory T cells [18, 50, 51].
In the testicular interstitial space, Leydig cells are both producers and target cells of galectin-1. Stimulation of cultivated Leydig cells from rat with recombinant galectin-1 inhibited testosterone production and caused a reduction in cell viability by eliciting apoptosis [52]. In contrast, the effects of added galectin-1 on growth and cell survival on the mouse MA-10 Leydig cell tumor line were biphasic depending on the concentration. High levels of galectin-1 induced apoptosis by mitochondrial and death receptor pathways, while low concentrations promoted cell proliferation and survival [53].
In relation to the presence and function of galectin-1 in the epididymis, the available data is very scarce. In rat epididymis, the presence of galectin-1 was demonstrated by immunohistological staining in the smooth muscle layers surrounding the epididymal ducts and in the blood vessels, while the epithelial cells were negative [43]. In murine epididymis, the expression of galectin-1 was investigated in protein extracts from all regions revealing a gradual increase from caput to distal parts (cauda). Moreover, galectin-1 was detected in fresh epididymal spermatozoa from cauda, but not from caput suggesting transfer from epithelial secretions and a role in sperm maturation This is supported by the presence of soluble galectin-1 in the epididymal fluid from cauda [40]. Galectin-1 was also found in human seminal plasma [54, 55].
Functional investigations on the role of galectin-1 in testis and epididymis were mainly derived from mice deficient in Lgals1 (Fig. 1). Caudal spermatozoa from Lgals1−/− mice exhibited defects in acquiring hyperactivation, an important step required for fertilization, i.e. the penetration of the egg coat. Moreover, Lgals1−/− sperm revealed a decreased ability for a progesterone-induced acrosome reaction, another process necessary to enter the egg. Mechanistically, this was related to a deficiency in sperm membrane potential hyperpolarization. Named effects were rescued by addition of exogenous recombinant galectin-1. These in vitro data are supported by lower fertilization rates in Lgals1−/− mice compared to wild type counterparts [40], emphasizing the relevance for galectin-1 in sperm function and fertilization. Of note, in contrast to the mouse, spermatozoa from rat cauda epididymidis showed very strong galectin-1 immunostaining around the plasma membrane of the head indicating species-specific differences in the presence of this lectin in spermatozoa from different regions of the epididymis [41].
Role in pathological conditionsSince galectin-1 is involved in the modulation of immune responses and shows anti-inflammatory properties in autoimmune diseases, investigations on the role of galectin-1 were prompted in a mouse model of experimental autoimmune epididymo-orchitis (EAEO). EAEO is a well-established rodent model of autoimmune-based inflammation mimicking pathology observed in the human testis and epididymis that ultimately can lead to infertility [51]. The pathology in the testis is characterized by production of pro-inflammatory and pro-fibrotic mediators (e.g. TNF, CCL2, activin A), immune cell infiltration, disruption of spermatogenesis, thickening of the lamina propria likely contributing to loss of the adluminal compartment and subsequent aspermatogenesis. Later stages of the disease show disruption of the BTB, extensive necrosis in seminiferous tubules and fibrotic remodeling [51, 56]. Our own data demonstrated that in the rat EAEO testis total expression of galectin-1 was downregulated, an observation likely based on the concomitant loss of galectin-1 expressing germ cells. Primary rat Sertoli cells demonstrated increased production of galectin-1 after challenge with TNF, a cytokine elevated in EAEO testis. Moreover, galectin-1 enhanced TNF-stimulated production of Il1a, Il6 and Ccl2 through MAP kinase activation in Sertoli cells [45]. In contrast, in a murine model of EAEO, the protein expression of this lectin was unchanged [39].
Induction of EAEO in Lgals1−/− mice led to a significant reduction in the incidence and severity of the disease as compared to wild type, showing only focal tubular damage and lower number of apoptotic germ cells. Surprisingly, these effects are opposite to established inhibitory function of endogenous galectin-1 in other immune privileged site such brain as documented by high susceptibility of Lgals1−/− mice to development of experimental autoimmune encephalomyelitis [35]. However, the role of galectin-1 in this model seems to be context- and cell-dependent as intraperitoneal administration of exogenous recombinant galectin-1 during the course of EAEO significantly reduced the severity of the disease. Therefore, a dual role of endogenous and exogenously administered galection-1 is evident in the control of autoimmune-based testicular inflammation [39].
Interestingly, under normal conditions testicular macrophages do not express galectin-1, while in EAEO testis, CD68+ macrophages located near granulomas show galectin-1 expression. It is plausible that galectin-1 secreted by newly arriving macrophages is implicated in enhanced TNF induced inflammatory effects in Sertoli cells as mentioned above [45].
Galectin-1 binds to LacNAc on branches of N- and O-glycans, which are synthesized by the synchronized activity of glycosyltransferases. There are three important post-translational mechanisms to form galectin-1 binding sites including: (a) activity of core 2 glucosaminyl (N-acetyl) transferase 1 (Gcnt1) for synthesis of core 2 O-glycans, which are the backbone of galectin-1 ligands, (b) suppression of ST6 beta-galactoside α-2-6-sialyltransferase 1 (St6gal1) activity, that abrogates galectin-1 binding to some terminal N-acetylglucosamines by adding α-2-6-sialic acid and (c) branching of N-glycans by mannosyl (α-1,3-)-glycoprotein ß-1,2-N-acetylglucosaminyltransferases (Mgat) like Mgat5 [57]. The experiments demonstrated that transcripts of St6gal1 were upregulated with a concomitant downregulation of Mgat5 in rat EAEO testis. In line with these findings, the binding of Sambucus nigra agglutinin, recognizing terminal α-2-6 sialic acid residues, was increased in EAEO testis. From a mechanistic perspective, the in vitro binding of galectin-1 to Sertoli and peritubular cells following an inflammatory challenge with TNF was reduced due to changes in the glycan composition. These findings indicate that α-2-6-sialylation of O- and N-glycans increases in inflamed testis and galectin-1 modulates inflammatory responses in Sertoli cells by enhancing the pro-inflammatory activity of TNF [45].
In seminal plasma, the levels of galectin-1 were significantly elevated in azoospermic patients [54, 55].
Galectin-3Role in homeostasisGalectin-3 is the only member of the chimera type of galectins, which consists of one CRD connected to the amino-terminal domain that mediates oligomerization into pentamers [29]. Intracellularly, galectin-3 is localized in the nucleus and cytoplasm, but is also found outside of the cell. Galectin-3 influences different inflammatory processes through its interaction with certain immune cell populations such as neutrophils, monocytes or macrophages, thereby facilitating their recruitment and migration to the inflammatory site. It also interacts with a variety of cell types involved in wound healing including monocytes, macrophages, neutrophils, keratinocytes, and fibroblasts (reviewed in [28]).
However, the testicular or epididymal function of galectin-3 is largely unknown. Galectin-3 expression at the protein and gene levels were found in the testes of human, boar, bull, rat and mouse [58,59,60,61,62].
In the healthy testis, Sertoli cells, germ cells, peritubular cells and macrophages mainly express galectin-3. Similar to galectin-1, the expression levels of galectin-3 increases during testicular development [43, 58, 59]. In rat, at postnatal day (PND) 5 before puberty, only macrophages were positive for galectin-3, with peritubular cells following from PND 20 after commencement of spermatogenesis. Sertoli and Leydig cells were seen positive both at PND 50 and 70 [43].
Here, it needs to be mentioned that some species-specific differences in galectin-3 expression were reported (Table 1). In relation to cellular localization of galectin-3 in the testis, species specific differences were noted for mouse, bull and boar as compared to rat and human testis (summarized in Table 1) ([43, 47, 58, 59, 61,62,63,64], own unpublished data). In contrast to rat and human, Sertoli cells from mouse, bull and boar do not express galectin-3 and in these species the presence of this lectin is limited to peritubular cells, macrophages and/ or late stages of germ cells (Table 1). In cultivated porcine Sertoli cells galectin-3 expression was stimulated in a dose-dependent manner by FSH, epidermal growth factor and TNF [58], while in Leydig cells galectin-3 is involved in the regulation of steroidogenesis and secretion of testosterone. Depletion of galectin-3 in murine Leydig cells resulted in increased expression of enzymes crucial for testosterone biosynthesis, such as Cyp11a1, Hsd3b1, and Cyp17a1. This upregulation was associated with elevated secretion of testosterone and androstandiol [65].
Analysis of the physiological function of galectin-3 built largely on constitutive knock-out mice for galectin-3 (Lgals3−/−). In this context, Lgals3−/− mice showed (i) a delay in the first wave of spermatogenesis, (ii) decreased number of germ cells at PND 5 and 15 as well as (iii) an impaired maturation of Sertoli cells. Although adult animals are fertile, the testicular weight was significantly reduced compared to age-matched wild type controls. To determine the causes of testicular atrophy, testicular apoptosis was monitored and demonstrated an elevated rate of germ cell apoptosis due to a perturbed Bcl-2, Bax and Bak expression ratio, molecules responsible for live/death balance in cells. Interestingly, the expression of enzymes involved in steroid biosynthesis as well as levels of serum testosterone were increased in Lgals3−/− mice. Moreover, galectin-3 seems to play a role in the control of the testicular macrophage population since the number of F4/80+CD11b+ macrophages within the population of CD45+ immune cells was reduced in Lgals3−/− testis [65]. An apoptosis-related function of galectin-3 in the testis was highlighted in studies showing upregulated expression of galectin-3 in the testes of Nucling-deficient mice. Nucling interacts with galectin-3 and mediates apoptosis by inhibiting its expression via NF-κB signaling [66]. Altogether, the studies point to an important role of galectin-3 in the regulation of spermatogenesis, steroidogenesis and maintenance of the testicular macrophage population.
In the epididymis and ejaculate, galectin-3 is present in epididymal cells (narrow, basal, principal cells), ejaculated spermatozoa and extracellular vesicles in human seminal plasma [60,61,62]. Expression levels of galectin-3 were seen lower in caput than in the distal parts (corpus, cauda) of the epididymis in human, bull and rat [43, 61, 62]. In development, at PND 5 in all parts of the epididymis (initial segment, caput, corpus and cauda) only interstitial macrophages in the connective tissue between the epididymal duct were positive for galectin-3, while at PND 20, intraepithelial macrophages additionally acquired expression. During adulthood (PND 50 and 70), in addition to macrophages, narrow cells in the initial segment, as well as principal and basal cells in the corpus and cauda epididymidis, began to express galectin-3 [43]. A similar expression pattern was observed in the bull epididymis. Interestingly, bull spermatozoa, which were negative for testicular galectin-3, acquired its expression in the caput epididymis and subsequently lost it during their passage through the corpus and cauda [62]. In combination, these data point to a role of galectin-3 in epididymal sperm maturation, but it needs to be noted that firm evidence is lacking. Variations in the galectin-3 presence in the immature and mature testis and epididymis may be related to the hormonal status, progression of spermatogenesis, expression of neo-antigens on the surface of spermatozoa and development of immune tolerance to haploid germ cells.
Role in pathological conditionsGalectin-3 has been discussed as a marker of aggressiveness in testicular tumors, with LGALS3 transcripts down-regulated in malignant Sertoli cell tumors, but seen increased in non-seminomatous testicular germ cell tumors [59]. In human biopsies obtained from infertile men with different diagnosis of testicular impairment (Sertoli cell only syndrome (SCOS), arrest of spermatogenesis at the meiotic or spermatid stage) an increased expression of galectin-3 was accompanied by transfer of the protein in Sertoli cells from the cytoplasm to the nucleus [58]. In line with these data, nuclear staining of Sertoli cells in SCOS was also reported in earlier studies by Wollina et al. [47]. In the serum and seminal plasma from azoospermic patients the levels of galectin-3 were significantly upregulated as compared to fertile men [55].
Other galectinsIn a recently published study, the expression of galectin-9 was for the first time reported on the surface of testicular immune cells (CD3+, CD4+, CD8+ T cells, NK cells) from mice using flow cytometry measurements [67]. Galectin-9 is a ligand for the T-cell immunoglobulin and mucin-domain containing-3 (TIM-3) receptor. Both, in mouse and human, binding of galectin-9 to TIM-3 leads to apoptosis of Th1 and Th17 cells and induces immune tolerance [30]. Furthermore, galectin-7 was postulated as a marker of immature Sertoli cells [
留言 (0)