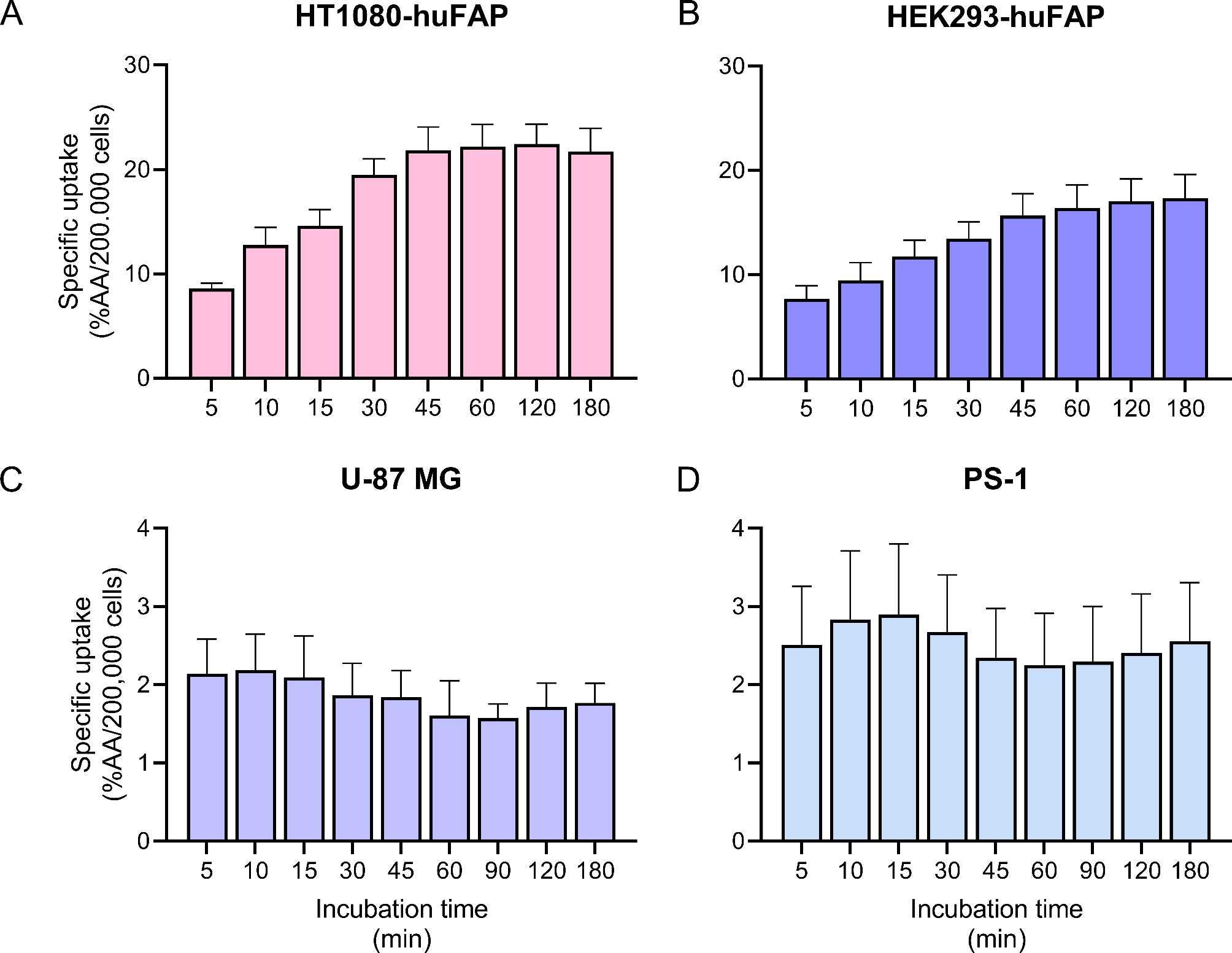
Cancer cell lines transduced to (over-)express human FAP are the most commonly used models to evaluate the potential of novel FAP-targeted radiopharmaceuticals in preclinical studies. However, these models do not accurately mimic the patient situation where the FAP-expression is most often found on CAFs present in the tumor stroma, and it is unclear if these transduced cells mimic the physiological expression and behavior of FAP. To compare tracer behavior in commonly used cell lines, we determined uptake and localization of [111In]In-FAPI-46 and RTX-1370S in HT1080-huFAP, HEK293-huFAP, U-87 MG, and an immortalized FAP-expressing fibroblast PS-1. In line with expectations, the highest FAP expression was found in the two transduced cell lines HT1080-huFAP and HEK293-huFAP, resulting in 6–10 fold higher radiotracer uptake in vitro and 2-fold higher radiotracer binding ex vivo compared to the endogenously FAP-expressing U-87 MG and PS-1. Even though HEK293-huFAP cells demonstrated the highest FAP expression, this was not resembled by the uptake of [111In]In-FAPI-46, which might indicate that not all of the expressed FAP was available for tracer binding. Alternatively, it might be that the different cell lines have different binding kinetics. The lower levels of FAP expression and [111In]In-FAPI-46 uptake observed for PS-1 in vitro and U-87 MG ex vivo are most likely more representative for patients with FAP-positive CAFs infiltrating the tumors (e.g., colon, head and neck cancer, or non-small cell lung cancer), or U-87 MG even in vitro for cancer cells with FAP-expression (e.g., sarcoma, mesothelioma) [15], as it is questionable if the huFAP transduced models with homogeneous over-expression of FAP mimic the physiological expression levels of patients with these cancers.
In addition to the level of uptake, the cellular processing of the radiotracer can influence its performance for FAP-TRT. In line with previously published studies, our in vitro data demonstrated that [111In]In-FAPI-46 is rapidly and almost fully internalized [7, 13]. In contrast, confocal microscopy demonstrated that RTX-1370S was at least partly membrane-bound for HEK293-huFAP cells, whereas it is mainly internalized in the other three FAP-expressing cell lines. Unexpectedly, incubating the cells at 4 °C did not alter the ratio between the membrane-bound and internalized [111In]In-FAPI-46 for any of the four cell lines. However, a statistically significant decrease in total internalized [111In]In-FAPI-46 fraction was observed for HT1080-huFAP and HEK293-huFAP. In contrast, confocal microscopy demonstrated that RTX-1370S remained more membrane-bound at 4 °C in both cell lines. Enzymatic activity, and hereby the internalization rate, was expected to decrease at 4 °C, compared to the physiologically relevant temperature of 37 °C [26]. However, if FAP internalization were a passive process, this would be less impaired at 4 °C than active transport internalization [27]. Additionally, the observed difference in [111In]In-FAPI-46 and RTX-1370S localization could be caused by the Alexa Fluor™ 568 group that contributes to the larger size of the fluorescent tracer, and that can lead to a higher hydrophilicity of RTX-1370S over FAPI-46. Thus the observed differences could also be the result of differences between the tracers. This has also been observed in a study by Millul et al., who compared multiple FAP-targeted radiotracers using two huFAP transduced cell lines. The study demonstrated complete internalization of FAPI-46, while FAP-2286 remained membrane-bound in both cell lines [13]. Similar to our study, the researchers removed the surface-bound radiotracer with an acid glycine buffer (pH 2.8), which is commonly used to interrupt target-ligand interactions, assuming that this interaction is pH dependent [28]. However, studies have shown that using an acid wash to separate the membrane-bound fraction is not always effective and reliable, and is dependent on the protein’s sensitivity to pH and the type of target-ligand interaction [29]. It has been confirmed that binding affinity of FAP for a ligand can be pH dependent [30], but it is unclear at which pH the interaction between a FAP-targeted small molecule and FAP could be disrupted. This target-ligand pH dependency could be different for the cyclic peptide FAP-2286 and FAPI-46, and further research could help identify if this acid wash is the optimal strategy to separate membrane-bound small molecule FAP inhibitors from FAP. Thus, based on our data it remains inconclusive if the observed differences in localization of [111In]In-FAPI-46 and RTX-1370S are due to differences in tracer behavior, the difference in the tracer concentration (1 nm and 10 nm, respectively), or due to suboptimal separation of membrane-bound radiotracer. Nevertheless, confocal microscopy did demonstrate a different localization pattern for RTX-1370S in HT1080-huFAP and HEK293-huFAP cells, indicating that tracer behavior is impacted by the model.
There are multiple explanations possible for the observed differences between HT1080-huFAP and HEK293-huFAP. Although both have been transduced to express the huFAP gene, the transductions were performed using different vectors and protocols, which could introduce differences in the expression levels of FAP. Additionally, the huFAP introduced in the HEK293 cell line is fused with GFP, and this fusion protein could alter the internalization kinetics of FAP. The impact of the GFP fusing affecting the FAP internalization is also supported by the observation of a high GFP signal around the nucleus, and almost 30% of the FAP-GFP not co-localizing with RTX-1370S. Thus, it seems that not all FAP is available for tracer binding, potentially due to FAP accumulation in the endoplasmic reticulum. This is in line with our data demonstrating the highest levels of FAP protein and mRNA expression in HEK-293-huFAP cells, which did not result in the highest [111In]In-FAPI-46 uptake. Thus, our data demonstrates various differences in FAP expression, internalization, and processing. Because these mechanisms are currently not fully understood, it is difficult to select the model with the highest translational value.
During our study, we observed that the FAP expression of PS-1 cells increased with higher passage numbers. This trend was observed with consecutive uptake studies, hence the higher SD in our results compared to that of the other cell lines. By IHC it was determined that FAP-expression was higher in passage 27 than in passage 22. The PS-1 cell line is hTERT immortalized, and a study by Park et al. found that hTERT expression can increase FAP expression by interacting with a transcription factor that is involved in regulating FAP expression (i.e., EGR1) [31]. Thus, endogenous expression should be interpreted carefully in the case of (hTERT) immortalization of primary cells, and ideally FAP expression prior to immortalization should be determined to establish if FAP expression was impacted by hTERT introduction.
Almost no studies described isolated primary human CAFs as a model to evaluate FAP-targeted radiotracers. Although these would have the highest translational value, it is practically challenging and difficult to reproduce. However, for FAP-TRT studies, a monoculture of CAFs would still not be adequate to study the complex aspects of indirect radiation of cancer cells after radiotracer binding to the CAFs. Thus, more advanced models such as co-cultures or 3D organoid models are necessary to determine and compare the FAP-TRT efficacy of radiotracers in vitro. Although these models are more complex, their value is extensively being researched [32,33,34]. Indirect co-cultures are the most user-friendly alternative to mono-cultures, as it allows separation of the two cell types while including the effects of the secretions and cross-talk between CAFs and the cancer cells. Nevertheless, the distance between the cells is still not representative of TRT in the clinical situation, and thus suboptimal to evaluate the efficacy of FAP-TRT. Alternatively, direct 2D co-cultures consisting of CAFs and cancer cells mimicking a variation of cancer types are extensively being researched to study the interaction between CAFs and cancer cells [33, 35, 36]; however, to our knowledge, no studies have been published describing TRT in these direct co-culture models to date. More complex 3D spheroid and organoid models are also becoming an attractive alternative to 2D cell culture. For example, Schuth et al. developed a personalized primary pancreatic ductal adenocarcinoma co-culture model to study the impact of CAFs on cancer cell response to chemotherapy [32]. However, studies describing TRT in such advanced models are lacking, implying that using radiotracers in a 3D organoid model is complicated.
For the evaluation of novel radiotracers, in vivo data remains essential to determine therapeutic efficacy and tumor-to-background ratios. The U-87 MG cells are an attractive model for FAP-TRT research because these cells stimulate attraction of murine fibroblasts in vivo. Thus, the level of FAP expression in this model depends on the attraction of enough murine fibroblasts, resulting in a heterogenic FAP expression. Despite this lower and heterogenic FAP-expression, we only observed a 2-fold lower binding of FAPI-46 to U-87 MG xenografts compared to HT1080-huFAP and HEK293-huFAP xenografts with in vitro autoradiography, whereas an almost 10-fold higher in vitro cell uptake was observed. One limitation of this research was the availability of fresh frozen HEK293-huFAP xenografts, hence no evaluation of the heterogeneity of binding in this CDX model could be performed. In a future study it would be valuable to include a large sample set with a variety of huFAP transduced cell lines to evaluate their differencesin vitro and in vivo, and to determine the impact on FAP-targeted radiotracer evaluation in more detail. Nevertheless, U-87 MG CDXs seem more suitable for in vivo TRT-efficacy studies, likely with a higher translational value compared to a xenograft established with a FAP over-expressing cell line. Next to U-87 MG, other cell lines have been used to create a FAP-positive CDX based on attracting murine fibroblasts, including the pancreatic cancer cell lines Panc-1 and PDAC299 [37,38,39]. PDAC299, for example, demonstrates high histopathological similarities with the stroma-dense pancreatic ductal adenocarcinomas in patients. Even though these xenografts can demonstrate resemblance to a patient’s tumor, the FAP expression in healthy organs remains different between mice and man, especially due to the higher levels of soluble FAP in mice [40]. Even more advanced in vivo models are available, such as the genetically engineered KPC mouse model for PDAC [41]. Although this model resembles patient histology more closely, these models are more labor-intensive, expensive, and not available for all tumor types. Studies using any in vivo model based on attraction of muFAP-expressing CAFs should carefully consider the radiotracer’s affinity for huFAP and muFAP. FAPI-46 demonstrated high and comparable affinity for both muFAP and huFAP, but FAP-2286, for example, does demonstrate a notable difference in affinity between the two isoforms [15, 21]. All in all CDXs and other in vivo models remain an approximation of the clinical situation, but with increased understanding of the FAP-expressing models, the most representative model can be selected.




留言 (0)