
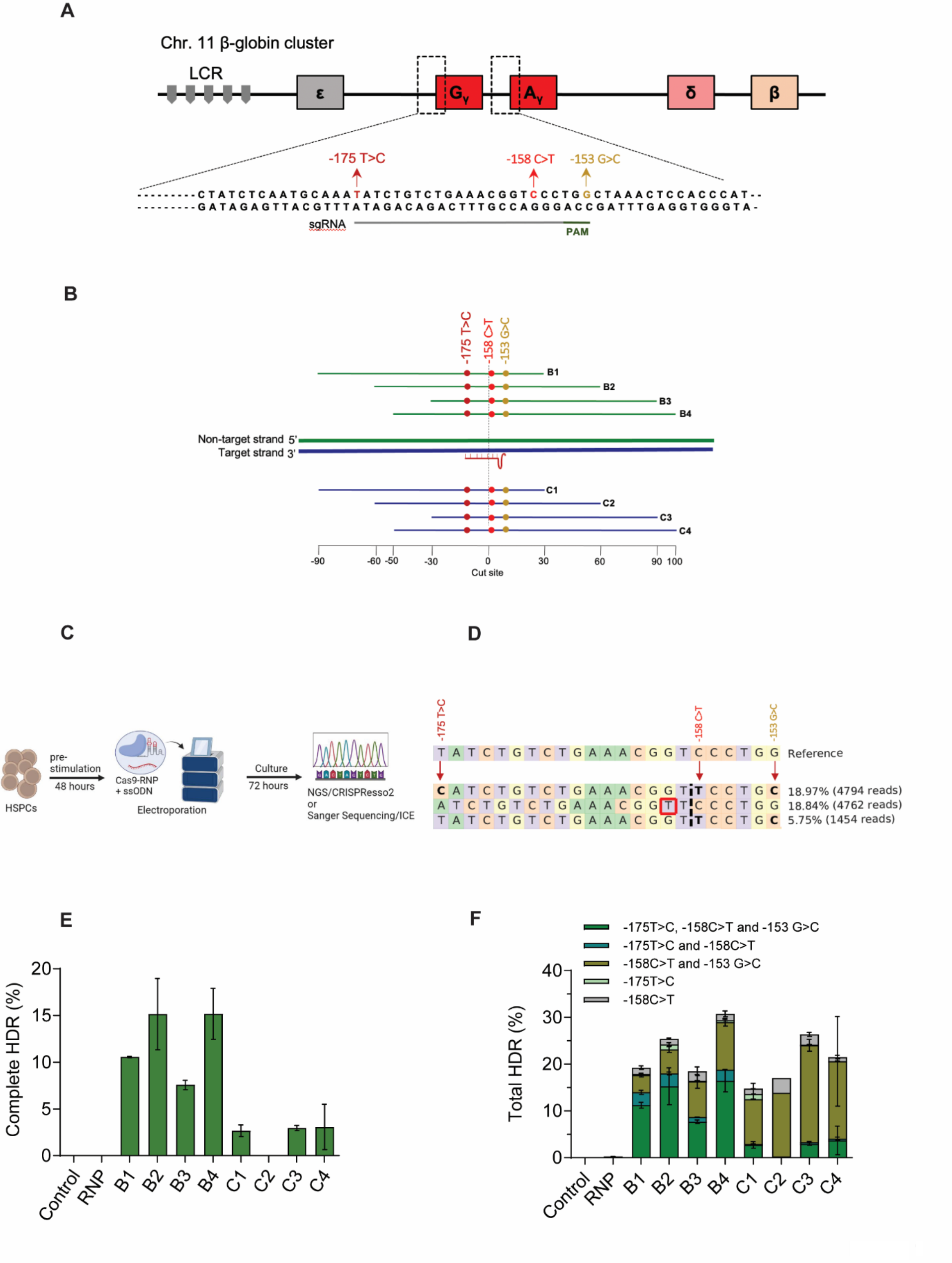
記住我





Human-induced pluripotent stem cell line UC was assessed for pluripotency prior to differentiation. Immunofluorescence and flow cytometry analysis showed that more than 99% of the cells co-expressed the pluripotency markers NANOG and OCT4 (Fig. 1B). It has been reported that a high dose of CHIR induces mesoderm, whereas a low dose induces endoderm [15]. To achieve co-differentiation of both endoderm and mesoderm from hPSCs in a single dish, we tested different combinations of ActA and CHIR (Fig. 1A) to precisely manipulate WNT signaling over three days (day 1–3).
Fig. 1
Optimization of co-differentiation of endoderm and mesoderm from hPSC. (A) A schematic representation of the differentiation protocol from Day 0 to Day 3. The endoderm differentiation scheme was adapted from Chen et al. Our optimized protocol facilitates the simultaneous induction of both endoderm and mesoderm. (Created in BioRender: https://BioRender.com/j57p699 ). (B) Immunofluorescence and flow cytometry analysis confirmed that most Day 0 hPSCs expressed NANOG and OCT4. Scale bar, 50 μm. (C) Quantitative PCR analysis. High levels of CHIR99021 promote the optimal proportion of mesodermal and endodermal co-differentiation compared to protocols with increased or decreased CHIR99021 concentrations. (D) Quantitative PCR analysis, (E) flow cytometry, and (F) immunofluorescence showed that 2.5% mTeSR1 induces an appropriate proportion of mesoderm and endoderm. (n = 3, *p < 0.05, **p < 0.01, ***p < 0.001, ns: no statistical differences)
Initially, we evaluated various concentration of CHIR within a three-day treatment protocol. Consistent with previous studies, the expression of mesoderm markers (TBXT+, MIXL1+) increased with higher concentrations of CHIR, while the expression of endoderm markers (SOX17+, FOXA2+) decreased (Figure S1A). Consequently, we choose the low dose of CHIR for further testing. Both the dosage and duration of CHIR treatment are critical for determining the fate of cells towards mesoderm or endoderm. The combination of ActA with decreasing concentrations of CHIR has been shown to generate highly purified DE [23]. We then sought to identify the optimal processing time for CHIR application. Compared to decreasing concentration, an incremental or constant concentration of CHIR favored mesoderm induction and suppressed endoderm differentiation. However, the constant concentration had a reduced effect on endoderm suppression (Fig. 1C). Therefore, maintaining a constant concentration of CHIR for three days appears to be an effective strategy for the simultaneous induction of both endoderm and mesoderm.
In our previous study, we demonstrated mTeSR1 (mT) contributes to the co-differentiation of endoderm and mesoderm for generating vascularized hepato-biliary organoids [25, 27]. To access the effect of mT on the co-induction of endoderm and mesoderm, various ratios of mT were added into the basic medium for three days. The expression of endoderm makers decreased with increasing ratios of mT, while the expression of mesoderm markers increased (Fig. 1D-F). Immunoflurescence and flow cytometry further confirmed that a 2.5% mT treatment achieved an optimal proportion of mesodermal (~ 30%) and endodermal (~ 70%) cells (Fig. 1F).
Bone morphogenetic protein 4 (BMP4) has been reported to induce mesoderm differentiation from hPSCs. Therefore, we tested the effect of supplementing BMP4 during the first three days of differentiation. Unexpectedly, BMP4 did not show a positive effect in our protocol (Figure S1B). The initial cell density of hPSCs can significantly affect differentiation efficiency. We tested different initial cell densities, applying the same ActA and CHIR treatment for three days. Immunofluorescence analysis of the mesoderm marker TBXT revealed that starting differentiation at cell densities between 80% and 100% resulted in stable mesodermal cell populations by day 3 (Figure S1C).
In summary, by manipulating WNT signaling and supplementing with mTeSR1, we achieved the co-induction of both endoderm and mesoderm from a single culture of hPSCs. This approach effectively prepares the cells for subsequent co-differentiation into pancreatic progenitors and endothelial cells.
Promotion of endothelial cells differentiation by VEGFA in pancreatic progenitors specializationWe used Chen’s protocol as a control, which effectively differentiates DE into pancreatic progenitors (PP) [23]. To generate vascularized pancreatic progenitors (vPP) from mesoderm and endoderm, we initially applied the same supplement as in Chen’s protocol from day 4 to day 10 (Figure S2A). By day 10, compared to Chen’s protocol, there was a significant increase in the expression of endothelial cell makers (CD34+, CDH5+) (Figure S2C). However, expression of the key pancreatic progenitor marker PDX1 was lower, even though NKX6.1 and SOX9 levels were higher than those in the control group (Figure S2C). The proportion of ECs was approximately 7% (Figure S2B). Therefore, we need to identify a strategy to promote EC differentiation without hindering PP specialization.
VEGFA and FGF2 are key differentiation factors that promote angiogenesis [28].We investigated the effectiveness of supplementing these angiogenic factors to facilitate EC differentiation during PP specialization (day 4–7). A high dose of VEGFA increased mRNA expression of CD31, while FGF2 did not have the same effect (Figure S2F). We then combined VEGFA and with the PP differentiation protocol to generate both ECs and PPs, referred to as the vPP protocol (Fig. 2A).
Fig. 2
Generation of Vascularized Pancreatic Progenitors (vPP). (A) A schematic representation of the PP and vPP differentiation protocol. The PP differentiation scheme was adapted from Chen et al. Our vPP protocol facilitates the simultaneous induction of both EC and PP from Day 4 to Day 10. (Created in BioRender: https://BioRender.com/d21p796). (B) Flow cytometry analysis of EC marker CD34. (C and D) Quantitative PCR analysis of EC markers (CD34, CD31, CDH5), PP markers (PDX1, SOX9, NKX6.1), and pericyte markers (PDGFRβ, α-SMA). (E) Immunostaining of generated PP (day 10) against PDX1 and CD31, comparing the vPP and Chen’s protocols. Scale bar, 250 μm. (F) Cell counting analysis. Statistical analysis of cell counts after re-inoculation for 3 days with an equal number of cells on Day 10 under both protocols. (G) Immunostaining of generated PP (day 10) against Ki67, comparing the vPP and Chen’s protocol. Scale bar, 100 μm. (n = 3, *p < 0.05, **p < 0.01, ***p < 0.001, ns: no statistical differences)
Compared to Chen’s protocol, the expression of endothelial cells makers (CD31+, CD34+, and CDH5+) significantly increased, with no adverse impact on PP differentiation markers (PDX1+, SOX9+, and NKX6.1+) (Fig. 2C). The proportion of ECs increased to 13.9% (Figure S2B). Additionally, pericytes, identified by the molecular markers PDGFRβ and α-smooth muscle actin (SMA), were more prevalent in vPPs (Fig. 2D, Figure S2D).
In contrast to prior methods that primarily focused on producing pure PPs, our approach successfully controlled the co-development of DE and mesodermal lineages within the same culture, leading to the formation of vascularized pancreatic progenitors (Fig. 2E, Figure S2E). The robustness of vPP differentiation was maintained, with both cell proliferation ability and cell numbers significantly improved compared to the original protocol (Fig. 2F-G).
To verify these results, we repeatedly induced vPP using H1 human embryonic stem cell lines, consistently achieving the same outcomes (Figure S5).
Transcriptome analysis of vascularized pancreatic progenitorsTo evaluate the transcriptomic profile of vPPs, we performed RNA sequencing on cells at day 3 and day 10 of vPP differentiation. Cells from Chen’s protocol and hPSCs were used as controls. Principal components analysis (PCA) and heatmaps demonstrated a clear distribution of all samples (Figure S3A and B).
Our optimized protocol, applied from day 1 to day 3, enhanced the expression of several makers involved in mesoderm development, including SFRP2, SMAD3, TBXT, LEF1, and DKK1 (Fig. 3A and C, Table S5), while it suppressed the expression of makers associated with endoderm development, such as APELA, BMP4, GATA4, GATA6, and HNF1B (Fig. 3A and D, Table S5). Gene ontology (GO) enrichment analysis revealed that differentially expressed genes (DEGs) were significantly involved in the biological process of mesoderm and endoderm development (Fig. 3B). This was further supported by gene set enrichment analysis (GSEA). Compared to controls, the gene set related to mesoderm formation was significantly up-regulated (P value = 0.024, FDR value = 0.1142, ES = 0.557), while the set related to endodermal cell differentiation was down-regulated (P value = 0, FDR value = 0.0026, ES=-0.792) (Fig. 3E-F). To further understand the underlying mechanisms, KEGG enrichment analysis of DEGs showed that pathways involved in oxidative phosphorylation (P value = 0, FDR value = 0, ES = 0.574) and linoleic acid metabolism (P value = 0.0050, FDR value = 0.0289, ES = 0.626) were significantly up-regulated in our optimized protocol in comparison to the previous method. Conversely, the PI3k-AKT signaling pathway (P value = 0, FDR value = 0.0305, ES=-0.4954) and FOXO signaling pathway (P value = 0.0028, FDR value = 0.0408, ES=-0.5061) were significantly down-regulated (Fig. 3G).
Fig. 3
Characterization of endoderm and mesoderm co-differentiation via RNA sequencing. (A) Heatmap representation of DEGs of endoderm and mesoderm co-differentiation from Optimized Protocol compared to control protocol (Chen’s Protocol). (B) Gene Ontology (GO) enrichment analysis of DEGs involved in mesoderm and endoderm development. (C and D) Up-regulation of mesoderm-associated genes and down-regulation of endoderm markers in the optimized protocol. (E and F) Gene set enrichment analysis (GSEA) indicating significant up-regulation of mesoderm formation and down-regulation of endodermal cell differentiation. (G) GSEA for KEGG pathway analysis
We further analyzed DEGs in vPPs, compare to PPs from Chen’s protocol [23]. Most genes related to endothelium development were up-regulated (Fig. 4A and C, Table S5). GO enrichment analysis of up-regulated DEGs in vPPs highlighted functions involved in angiogenesis (GO: 0001525), heart development (GO: 0007507), and positive regulation of angiogenesis (GO: 0045766) (Fig. 4B). GSEA suggested that the vPP protocol may promote processes related to hematopoietic cell lineage, vasculature development, and angiogenesis. Importantly, we did not find any down-regulated DEGs impacting pancreatic development.
Fig. 4
Characterization of vascularized pancreatic progenitors (vPP) via RNA sequencing. (A) Heatmap showing upregulated endothelium development-related genes in vPP from vPP Protocol compared to PP from Chen’s protocol. (B) GO enrichment analysis of up-regulated DEGs in vPP. (C) Up-regulation and down-regulation of endothelium development-related genes in vPP protocol, compared to Chen’s protocol. (D) GSEA results suggesting vPP protocol promotes processes of hematopoietic cell lineage, vasculature development, and angiogenesis
Key transcription factors in vascularized pancreatic progenitor differentiationTranscription factors (TFs) are specialized proteins that bind to specific regions of DNA and regulate gene transcription. The roles of TFs in gene regulation during vPP differentiation were further elucidated through bioinformatics analysis. On day 3, 154 TF genes were significantly differentially expressed, with 71 TF genes up-regulated in our optimized protocol compare to the control (Fig. 5A, Table S5). The biological processes associated with these up-regulated TF genes were linked to mesoderm development and vasculogenesis (Fig. 5C). By day 10, 59 TF genes were significantly differentially expressed, with 33 TF genes up-regulated in our optimized protocol compared to the control (Fig. 5B, Table S5). These upregulated genes continued to be associated with mesoderm development and vasculogenesis (Fig. 5D).
Fig. 5
Transcription factor analysis for vascularized pancreatic progenitors (vPP). (A) Volcano plot for differentially expressed TF genes of endoderm and mesoderm co-differentiation from Optimized Protocol compared to control protocol (Chen’s Protocol) on day 3. (B) Volcano plot for differentially expressed TF genes in vPP Protocol compared to PP from Chen’s protocol on day 10. (C) GO enrichment and gene correlation analysis of up-regulated differentially expressed TF genes associated with mesoderm development and vasculogenesis on day 3. (D) GO enrichment and gene correlation analysis of up-regulated differentially expressed TF genes associated with mesoderm development and vasculogenesis on day 10
Potency of vascularized pancreatic progenitors to differentiate into β-cellsTo demonstrate the potential of vascularized pancreatic progenitors to differentiate into β-cells, we applied the late-stage differentiation medium from Chen’s protocol and compared these cells with those directly differentiated using the same protocol. We examined markers of mature β-cells (NKX6.1, INS, and MAFB), immature β-cells (PDX1 and SOX9), and ECs (CD34). The expression levels of NKX6.1 and INS were significantly up-regulated in cells at day 27 that differentiated from vPP compared to the control, while other markers showed no significant differences (Fig. 6A). Immunofluorescence demonstrated co-expression of INS and MAFA in cells at day 27 (Fig. 6B). Flow cytometry analysis revealed that approximately 60% of vPP differentiated into insulin-producing β-cells by day 27, with the ratio of C-peptide+ cells being higher than in the control group (Fig. 6C). Insulin secretion in the culture supernatant at the end of the culture period was higher in cells differentiated from vPP compared to the control (Figure S4A). Transmission electron microscopy and immunofluorescence confirmed the presence of insulin-secreting granules in the β-cells derived from vPP (Figure S4B and C). These observations suggest that vPP can differentiate into insulin-producing β-cells and may do so more efficiently than direct differentiation.
Fig. 6
Differentiation of vPP into β-Cells. (A) Quantitative PCR analysis of mature β-cells (NKX6.1, INS and MAFB), immature β-cells (PDX1 and SOX9), and ECs (CD34) at day 27. (B) Immunofluorescence images showing co-expression of INS and MAFA in vPP-derived β-cells at day 27. (C) Flow cytometry analysis of insulin-producing β-cells (C-peptide+ cells), indicating a higher ratio in vPP-derived β-cells compared to the control. (n = 3, *p < 0.05, **p < 0.01, ***p < 0.001, ns: no statistical differences)
留言 (0)