
記住我







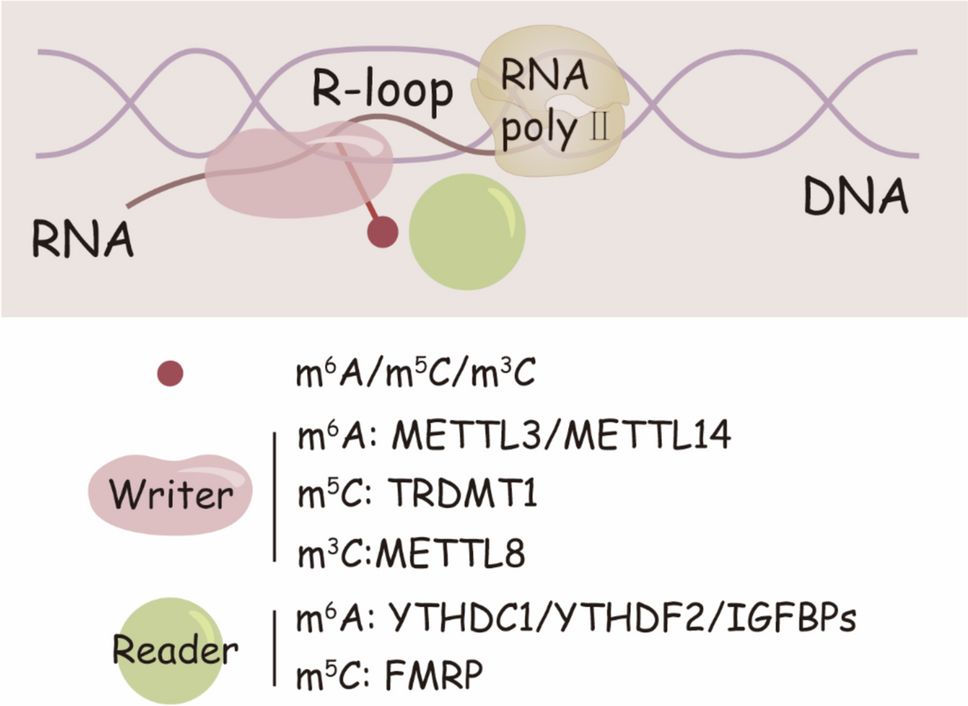
The proper formation and regulation of R-loops are essential for maintaining genomic stability. However, excessive accumulation or incomplete resolution of R-loops can lead to transcription-replication conflicts (TRCs), resulting in DNA replication obstacles, DSBs, and genomic instability [43]. RNA methylation plays a pivotal regulatory role by controlling both the formation and resolution of R-loops, ensuring their dynamic balance. When this balance is disrupted, abnormal R-loop accumulation can directly compromise genomic integrity (Fig. 2).
Fig.2
RNA methylation dysregulation promotes R-loop accumulation and genomic instability. When RNA methylation writers and readers, such as METTL3/METTL14 (m6A writer) and YTHDC1 (m6A reader), are dysregulated, the formation and resolution of R-loops become impaired. This results in increased R-loop accumulation, leading to TRCs and DSBs, both of which contribute to genome instability. Such genomic instability is a key factor in the development of diseases, including cancer and neurological disorders
For example, in patients with MDS harboring DDX41 mutations, excessive R-loop accumulation is a significant contributor to genomic instability. Under normal conditions, DDX41 interacts with METTL3/METTL14 and YTHDC1 to regulate R-loop formation and dissolution. However, mutations in DDX41 impair this regulatory function, leading to R-loop accumulation and an increased risk of DNA damage [38].
During transcription termination, R-loop accumulation is closely associated with genomic instability. m6A modifications, mediated by METTL3, regulate R-loop formation and resolution, ensuring proper transcription termination. Inadequate m6A modification can prevent RNA polymerase II from pausing effectively, resulting in transcriptional elongation, increased TRC frequency, and a higher incidence of DSBs, thereby exacerbating DNA damage and genomic instability [30, 37]. Additionally, the inactivation of DDX21 further impairs R-loop resolution, exacerbating transcriptional elongation and worsening genomic damage [37].
R-loop accumulation in telomeric regions is also strongly linked to genomic instability. TERRA forms R-loops to maintain telomere stability, and METTL3 stabilizes TERRA-associated R-loops via m6A modification. However, insufficient m6A modification destabilizes TERRA, reducing R-loop formation and leading to telomere shortening and genomic instability [31, 32].
Moreover, m3C modifications also play a role in R-loop-induced genomic instability. SUMOylated METTL8 promotes R-loop formation through m3C modification, particularly in GC-rich regions. Dysfunctional METTL8 or disrupted SUMOylation decreases R-loop formation, increasing genomic instability and transcriptional dysregulation [41].
In summary, excessive R-loop accumulation is a key driver of genomic instability and DNA damage. RNA methylation regulates the dynamic balance of R-loops and plays a central role in maintaining genomic stability. Dysregulation of either m6A or m3C modifications can lead to increased R-loop accumulation, significantly raising the risk of DNA damage. This underscores the critical role of methylation in regulating R-loops and its profound impact on genomic integrity.
When RNA methylation writers and readers, such as METTL3/METTL14 (m6A writer) and YTHDC1 (m6A reader), are dysregulated, the formation and resolution of R-loops become impaired. This results in increased R-loop accumulation, leading to TRCs and DSBs, both of which contribute to genome instability. Such genomic instability is a key factor in the development of diseases, including cancer and neurological disorders.
The role of R-loops and RNA methylation in DNA damage repairFollowing DNA damage, particularly during DSBs, complex repair mechanisms are activated, and the formation and resolution of R-loops play crucial regulatory roles in this process. RNA methylation, especially m6A and m5C modifications, significantly influences the progression and efficiency of the repair process by regulating the dynamic balance of R-loops.
m6A modification plays a pivotal role in DNA damage repair. Studies have demonstrated that METTL3 regulates R-loop formation and resolution via m6A modifications. After a DSB, if METTL3 or m6A modifications are insufficient, the recruitment of key repair proteins such as RAD51 and BRCA1 is impaired, leading to delayed DNA damage responses and a substantial reduction in repair efficiency [33, 38]. Inadequate m6A modification can also result in the accumulation of DNA-RNA hybrids, further obstructing the initiation of normal repair pathways, exacerbating the damage caused by DNA breaks [34].
m5C modifications are equally important in DNA damage repair. TRDMT1 stabilizes R-loops by generating m5C modifications, facilitating the binding of repair proteins. Cells deficient in TRDMT1 show significantly reduced homologous recombination repair efficiency, with repair proteins such as RAD51 and RAD52 failing to localize effectively to sites of DNA damage induced by reactive oxygen species (ROS) [39]. Additionally, FMRP, through its interaction with TET1, regulates m5C demethylation, promoting R-loop resolution. A lack of or reduced expression of FMRP leads to abnormal accumulation of m5C-modified R-loops, delaying their resolution and impeding DSB repair, thereby increasing the risk of persistent DNA damage [25].
R-loop resolution depends not only on RNA methylation but also on the action of specific enzymes. Methylation modifications assist in the recruitment of RNase H1, a key enzyme responsible for resolving R-loops. By preventing the excessive accumulation of R-loops, RNase H1 ensures proper DNA end resection and smooth progression of homologous recombination repair. For instance, ARID1A recruits METTL3 to methylate R-loops with m6A, promoting RNase H1 binding and accelerating R-loop resolution, which is crucial for efficient DNA repair [35].
In summary, RNA methylation plays a central role in DNA damage repair by regulating R-loop formation, accumulation, and resolution. By facilitating R-loop resolution and recruiting repair proteins, methylation ensures timely and efficient DNA damage repair. Mutations or functional disruptions in these methylation pathways can impair normal R-loop resolution, ultimately threatening genomic stability.
The impact of methylation and R-loops on DNA damage repair pathway selectionIn response to DNA damage, the selection of the appropriate repair pathway is crucial, particularly in balancing homologous recombination (HR) and non-homologous end joining (NHEJ). RNA methylation, especially m6A and m5C modifications, significantly influences R-loop formation and resolution, thereby modulating the recruitment of repair proteins and determining the choice between different repair pathways (Fig. 3).
Fig.3
R-loop regulation influences the choice between HR and Alt-NHEJ during DNA damage repair. In the presence of DSBs, R-loops may form at break sites, affecting the repair pathway selection. m6A and m5C modifications can facilitate HR by recruiting repair proteins such as RAD51/RAD52. In contrast, the failure to resolve R-loops can lead to the activation of Alt-NHEJ, mediated by PARP1 and associated proteins
m6A modification plays a key role in repair pathway selection. Studies have shown that cells lacking METTL3 exhibit a marked reduction in HR repair efficiency, while NHEJ repair remains relatively unaffected [33]. This suggests that m6A modification is essential for HR repair, and in its absence, cells shift their preference toward NHEJ. Furthermore, a reduction in the binding capacity of YTHDC1 to RAD51 compromises RAD51 recruitment to DNA damage sites, further weakening HR repair efficiency [38].
ARID1A ensures the preferential selection of HR repair by regulating m6A modifications on R-loops. ARID1A recruits METTL3 to DNA damage sites, promoting m6A modification of R-loops, and facilitates the recruitment of RNase H1 to dissolve R-loops, ensuring the smooth progression of HR repair [35].
m5C modification also plays a critical role in repair pathway selection. Studies have shown that TRDMT1-mediated m5C modifications at DNA damage sites enhance the binding of repair proteins such as RAD52 and RAD51 to R-loops, thereby promoting HR repair [39]. In TRDMT1-deficient cells, NHEJ repair is enhanced, indicating that m5C modification favors the high-fidelity HR repair pathway. Additionally, TRDMT1-mediated m5C modification has been shown to inhibit the alternative non-homologous end joining (Alt-NHEJ) pathway. m5C modification suppresses the activation of PARP1, reducing the recruitment of Alt-NHEJ repair proteins like XRCC1 and LIG3, thereby promoting HR repair. PARP inhibitor studies further support the role of m5C modification in repair pathway selection, demonstrating that m5C limits Alt-NHEJ activity and favors HR repair [40].
FMRP also facilitates HR repair by regulating m5C demethylation and R-loop resolution. Unlike other repair factors, FMRP enhances HR repair efficiency by promoting RAD51 recruitment independently of the BRCA1/BRCA2 pathway, highlighting the unique regulatory role of m5C modification in ensuring the efficiency of HR repair [25].
In conclusion, RNA methylation influences DNA damage repair pathway selection by regulating R-loop dynamics and the recruitment of repair proteins. Disruptions in methylation modifications not only interfere with R-loop homeostasis but also lead to improper repair pathway selection, ultimately compromising genomic stability.
In the presence of DSBs, R-loops may form at break sites, affecting the repair pathway selection. m6A and m5C modifications can facilitate HR by recruiting repair proteins such as RAD51/RAD52. In contrast, the failure to resolve R-loops can lead to the activation of Alt-NHEJ, mediated by PARP1 and associated proteins.
留言 (0)