
記住我
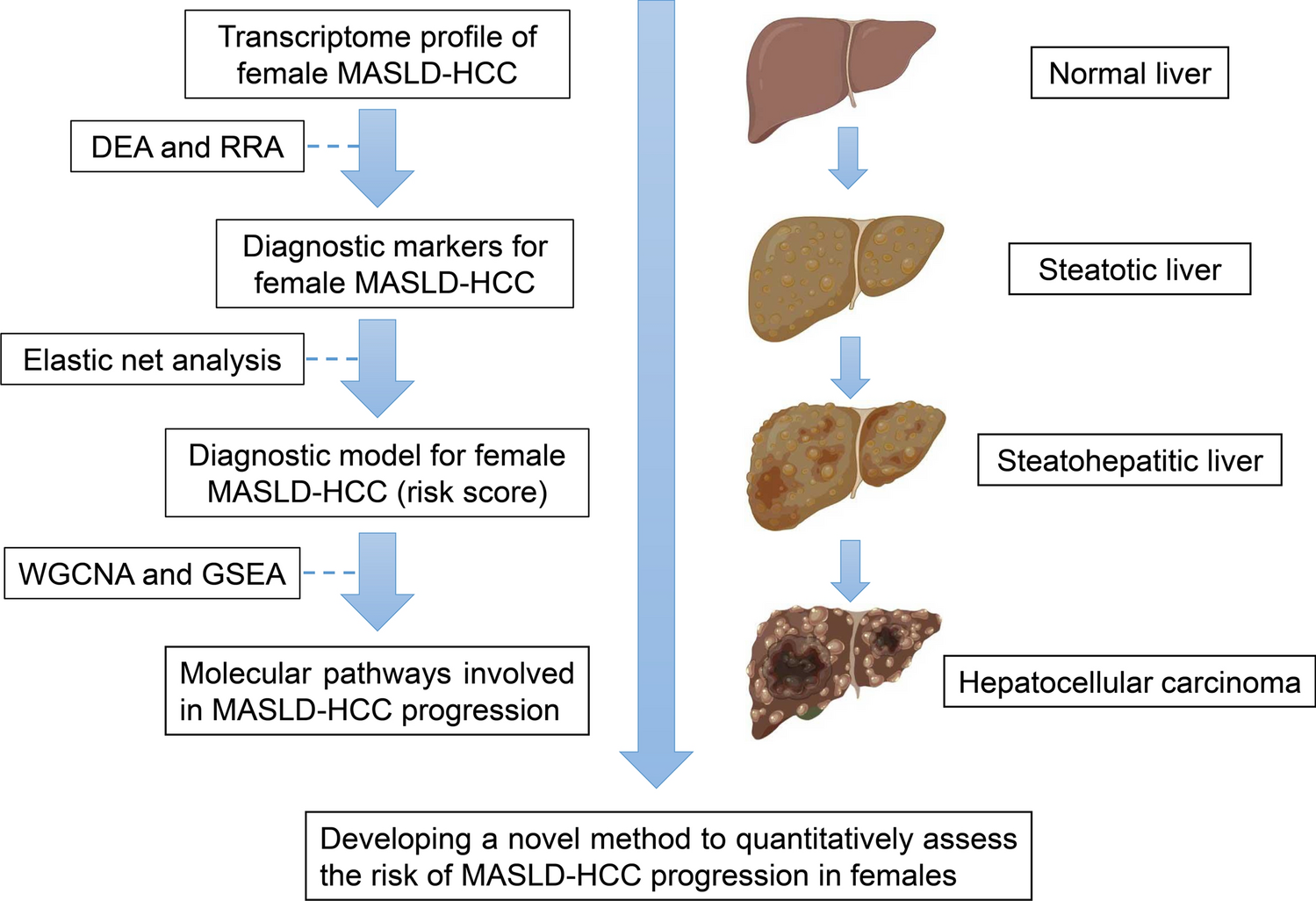





All 145 genes were identified as co-expressed AIGs by Venn diagram (Fig. 1A). 33 genes were found to have potential prognostic value by univariate Cox regression analysis (Supplementary Fig. 1). Unfortunately, however, of these 28 genes, AGER, CTSS, IGF2, EGFR, EPOR, and IKBKB were not differential expressed in bladder cancer versus paraneoplastic + normal tissue (Supplementary Fig. 2A–G), and CALCA, CCN2, EDNRB, JUN, LRP1, PENK, TGFB3, IL33, S100A6, GHR PTPN11, IGF1, NGF, NGFR, NR3C1, A2M, FGFR1, ELN, PPARG, AGTR1 and PDGFRA whose expression levels of contradicted the prognostic status (Supplementary Fig. 2A–G). Ultimately, GBP2, PLCG2, NFKB1, PTK2B and IL7 were found to be lowly expressed in bladder cancer by unpaired expression analysis (Fig. 1B), and PLCG2, NFKB1 and IL7 were also lowly expressed in paired expression analysis (Fig. 1C). Low expression of the above five genes in BLCA was correlated with poor OS by prognostic K-M plots (p < 0.05) (Fig. 1D), and moreover, low expression of GBP2, PTK2B and IL7 was associated with poor PFI and poor DSS (Fig. 1E, F). Collectively, this study initially identified GBP2, PLCG2, NFKB1, PTK2B and IL7 as tumor suppressor gene with significant prognostic value in BLCA.
Fig. 1
Identification of five aging-immune related genes in the TCGA-BLCA cohort. A Preliminary screening of potential prognostic AIGs by Venn diagram; unpaired expression (B) and paired expression (C) of GBP2, PLCG2, NFKB1, PTK2B, IL7; overall survival (D), progress free interval (E) and disease-specific survival (F) of GBP2, PLCG2, NFKB1, PTK2B, IL7
3.2 Identification of pathway enrichment for co-expression AIGsBy STING, the protein interaction network of 35 co-expressed AIGs was identified, as well as EGFR, VEGFA, IGF1, CTGF and JUN were found to be at the centre of the network (Fig. 2A). Through GO analysis, the above genes were found to be enriched in positive regulation of MAPK cascade, response to peptide and positive regulation of fibroblast proliferation, regulation of epithelial cell proliferation, neuroinflammatory response (BP); vesicle lumen, platelet alpha granule, collagen-containing extracellular matrix, transport vesicle, CD40 receptor complex (CC); Signaling receptor activator activity, integrin binding, protein serine/threonine/tyrosine kinase activity, according to the protein binding and G protein-coupled receptor binding (MF), whose the p.adjust < 0.05 and qvalue < 0.05 (Fig. 2B). KEGG analysis showed that the functions enriched in MAPK signaling pathway, Proteoglycans in cancer, Chemical carcinogenes-reactive oxygen species, and PD-L1 expression and PD-1 checkpoint pathway in cancer, Leukocyte transendothelial migration, Prostate cancer and Renal cell carcinoma et al. (Fig. 2C). Subsequently, further analysis of the five tumor suppressor genes with significant prognostic value revealed weak correlations for GBP2, PLCG2, NFKB1, PTK2B and IL7, with NFKB1 at the centre (Fig. 2D). The GO-KEGG analysis of the above genes indicated that they were mainly enriched in B cell differentiation, response to tumor necrosis factor and lymphocyte differentiation (BP); apical dendrite, cell leading edge (CC); growth factor activity, ionotropic glutamate receptor activity (MF); Chemokine signaling pathway, NF − kappa B signaling pathway, B cell receptor signaling pathway (KEGG), whose the p.adjust < 0.05 and qvalue < 0.05 (Fig. 2E, F).
Fig. 2
Identification of pathway enrichment for aging-immune related genes. 35 protein interaction networks co-expressing AIGs (A), GO (B) and KEGG (C) analysis; protein interaction networks for five AIGs (D), GO (E) and KEGG (F) analysis
3.3 Correlation of the hub AIGs NFKB1 and IL7 with the BLCA clinicopathological featuresIn the TCGA-BLCA disease cohort, low expression of NFKB1 and IL7 was found to be associated with the development of lymph node metastasis (N1&2&3) and distant metastasis (M1) (Fig. 3B, C). Additionally, low expression of PLCG2 and PTK2B was associated with the development of lymph node metastasis (N1&2&3) (Fig. 3B) and low expression of GBP2 was associated with high T-stage (T3&4) (Fig. 3A). Low expression of PLCG2 and PTK2B correlated with high stage (Stage III&IV) and high grade (Fig. 3E, F). Notably, using 70 years as the advanced age cut-off, only NFKB1 and IL7 were significantly underexpressed in advanced age patients (> 70 years) (Fig. 3D). The remaining clinicopathological features with NFKB1 and IL7 were shown in Supplementary Table 1. Combined with the study objectives and the correlation between paired analyzes and clinicopathological features, this study tentatively identified NFKB1 and IL7 as hub AIGs in bladder cancer. Further exploration revealed 1,3,5 year diagnostic AUC curves of 0.428,0.482 and 0.443 for NFKB1, and 0.372,0.428 and 0.448 for IL7 respectively (Fig. 3G), as well as the time dependent AUC curves for NFKB1 and IL7 are shown in Fig. 3H. Interestingly, after combined age groups, the diagnostic AUC curves for NFKB1 and IL7 were significantly higher at 0.595 and 0.582 respectively (Fig. 3I). Therefore, the study constructed a prognostic nomogram of the three elements of NFKB1, IL7 and age with concordance = 0.592, p < 0.05 (Fig. 3J), and its 1,3,5 year prognostic calibration curve was well fitted to the ideal curve, especially for 3 years (Fig. 3K). By K-M curves, high expression of this combined indicator was found to be significantly associated with poor OS, showing a pro-carcinogenic effect (Fig. 3L), which would be somewhat contradicted by the initial diagnosis of NFKB1 and IL7 as suppressor genes in bladder cancer (Fig. 1B, D, 3A–F). In order to reconfirm in the expression of IL7 and NFKB1 in bladder cancer, the GSE121711, GSE45184 and GSE3167 datasets were used in this study. In GSE3167, IL7 was significantly under-expressed in tumor patients, while the trend in GSE121711 and GSE45184 was not statistically significant (Supplementary Fig. 3A–C); In GSE121711, NFKB1 was significantly under-expressed in tumor patients, while the trend in GSE45184 was not statistically significant (Supplementary Fig. 3D, E). The above results were consistent with the TCGA-BLCA results, which demonstrated that IL7 and NFKB1 were lowly expressed in bladder cancer.
Fig. 3
Association of aging-immune related genes with clinicopathological features. Correlation of five AIGs with T (A), N (B), M (C), age (D), stage (E) and grade (F); G 1,3,5 year diagnostic AUC curves for NFKB1 and IL7; H time dependent AUC curves; I diagnostic AUC curves for NFKB1 and IL7 combined with age; J prognostic nomogram for NFKB1, IL7 combined with age; K prognostic calibration curves; L overall survival
3.4 Investigation of the correlation between hub AIGs with immune infiltration, ferroptosis and m6A methylationA negative correlation between NFKB1 expression and TMB score was found (pSpearman = 0.12, p < 0.05), while there was no correlation for IL7 (Fig. 4A). TIDE scores were lower in patients with low NFKB1 expression by ICB response analysis, who had good immune checkpoint blockade (ICB) therapy efficacy and longer survival, while there was no correlation with IL7 (Fig. 4B). Further exploration of the relationship between NFKB1 and IL7 and immune checkpoint-related genes revealed that the NFKB1 low expression group (G1) had significantly lower expression of CD274, CTLA4, HAVCR2, LAG3, PDCD1, PDCD1LG2 and TIGIT and higher expression of SIGLEC15 (Fig. 4C); the IL7 low expression group (G1) had significantly lower expression of CD274, CTLA4, HAVCR2, LAG3, PDCD1, PDCD1LG2 and TIGIT (Fig. 4C). Subsequently, the correlation between NFKB1 and IL7 expression and immune infiltration was assessed by the xCELL algorithm suggesting that the NFKB1 low expression group (G1) had elevated T cell CD4+ Th1, Eosinophil, T cell NK infiltration, while T cell CD4+ Th2, Mast cell, Macrophage Monocyte, T cell CD4+ memory, and T cell regulatory infiltrates were low (Fig. 4D). In contrast, T cell CD4+ Th2, Mast cell, T cell CD4+ effector memory, Myeloid dendritic cell activated, Macrophage, Macrophage M1, Monocyte, T cell CD4+ memory, T cell regulatory, T cell CD8+, B cell infiltration are low in the IL7 low expression group (G1) (Fig. 4D). Subsequently, the dryness score revealed a significantly higher degree of dryness in the NFKB1 and IL7 low expression group (G1) than in the high expression group (G2) and the normal group (Fig. 4E).
Fig. 4
Immunological correlation analysis of NFKB1 and IL7. A Correlation of NFKB1 and IL7 with TMB scores; B ICB response; C immune checkpoint-associated genes; D immune infiltration heat map under the xCELL algorithm; E dryness score. C1 gene low expression group, C2 gene high expression group
Subsequently, the ferroptosis-related genes SLC7A11, ATP5MC3, ACSL4, ATL1, NFE2L2, NCOA4, PCAT3, EMC2, CS, HSPA5, CARS1, CISD1, SAT1, DPP4, and CDKN1A were explored to be less expressed in the NFKB1 low expression group (G1) (Fig. 5A). SLC7A11, ACSL4, NFE2L2, NCOA4, LPCAT3, EMC2, HSPA5, CISD1, SAT1, DPP4, and CDKN1A were significantly lower in the IL7 low expression group (G1) (Fig. 5B). Meanwhile, most of the m6A methylation-related genes in the NFKB1 low expression group (G1) showed a significant low expression, except for METTL3 and IGF2BP1 which were not significant (Fig. 5C); the situation within the IL7 low expression group (G1) was similar (Fig. 5D).
Fig. 5
Correlation of NFKB1 and IL7 with ferroptosis and m6A methylation. A Ferroptosis-related genes; B m6A methylation-related genes. C1 gene low expression group, C2 gene high expression group
To further clarify the signaling pathways of NFKB1 and IL7 in bladder cancer, this study performed single-gene GSEA pathway enrichment analysis of IL7 and NFKB1, respectively. The correlation results suggested that there were multiple identical pathways for IL7 and NFKB1, such as immunotherapy-associated pathways (e.g., CANCER_IMMUNOTHERAPY_BY_PD1 AND CTLA4_BLOCKADE), various immune cell-associated pathways (e.g., IMMUNOREGULATORY_INTERACTIONS_BETWEEN_A_LYMPHOID_AND_A_NON_LYMPHOID_CELL, NEURAL_CREST_CELL_MIGRATION_IN_CANCER, and NATURAL_KILLER_CELL_MEDIATED_CYTOTOXICITY), and critical signaling pathways (e.g., JAK_STAT_SIGNALING_PATHWAY, INTERFERON_SIGNALING, andPI3K_AKT_SIGNALING_IN_CANCER) (Supplementary Fig. 4A, B).
3.5 Construction of aging-immune related prognostic clusters and risk-prognosis modelsAs only two genes, NFKB1 and IL7, had the highest group mean concordance based on K = 2, cluster analysis was performed to obtain two two clusters, with 153 persons for AIG-high risk and 141 persons for AIG-low risk (Fig. 6A). Analysis of expression differences by clusters suggested that NFKB1 expression was higher and IL7 expression was lower in the AIG-high risk group relative to the AIG-low risk group (Fig. 6B). Combined with the OS results, patients with the AIG-high risk cluster had a significantly shorter OS (Fig. 6C), and therefore the AIG-high risk cluster (C1) was identified as the carcinogenic group, while the AIG-low risk cluster (C2) was the carcinogenic group. Cox regression analysis was used to construct a risk prognostic model based on NFKB1 and IL7 to obtain the optimal model AIC = 1884.9481 and Riskscore = (0.0581)*NFKB1 + (− 0.2285)*IL7. Analysis of the relationship between NFKB1-IL7 and survival status of BLCA patients showed that the high risk group had a lower number of survival states and relatively lower NFKB1 and IL7 expression (Fig. 6E). Analysis in the TCGA-BLCA cohort suggested that OS was shorter in the high risk group (Fig. 6E), and that the ROC-1, 3- and 5-year OS were 0.631, 0.567 and 0.539 respectively (Fig. 6E). The DCA curve further showed that the combined prognostic model of NFKB1 and IL7 had more application and better prognosis than the single gene prediction model (Fig. 6D). The prognostic model was then explored for immune correlation by the quanTIseq algorithm, which revealed a significant negative correlation with macrophage M1 and macrophage M2 expression (Fig. 6F).
Fig. 6
Prognostic risk model for the combination of NFKB1 and IL7. A Heat map at K = 2 with 153 AIGs-high and 141 AIGs-low; B Differential expression of NFKB1 and IL7 in different clusters; C OS under clusters; D DCA curve analysis of NFKB1, IL7 and risk prognostic model; E Distribution of risk scores, survival of TCGA-BLCA patients status and heat map of hub AIGs, as well as overall survival and 1,3,5-year diagnostic AUC curves; F Correlation of the risk prognosis model with macrophage M1 and M2 polarization
3.6 Characterization of differential expressed genes and associated pathways in different clustersTo explore the molecular mechanisms underlying the differences in prognosis between the different clusters, 123 genes were identified, of which 54 were up-regulated and 69 were down-regulated (FC ≥ 1, p < 0.05) in the two clusters (Fig. 7A, B). KEGG analysis of the different genomes suggested that up-regulated genes were clustered in retinol metabolism, bile secretion and cytokine-cytokine receptor interaction (Fig. 7C); down-regulated genes were enriched in circadian entrainment, dopaminergic synapse and dopaminergic synapse, dopaminergic synapse and cytokine-cytokine receptor interaction (Fig. 7D). GO analysis revealed that up-regulated gene aggregation was involved in epidermal cell differentiation, cell junction and monocarboxylic acid transmembrane transporter activity (Fig. 7E). The down-regulated genes were enriched in premrna binding, ion channel complex and transmembrane transporter complex (Fig. 7E). Moreover, GSEA analysis further showed that AIG-high risk (C1) and AIG-low risk clusters (C2) on HYPOXIA, APOPTOSIS, ANGIOGENESIS, GLYCOLYSIS and signaling pathways (such as PI3K_AKT_MTOR_SIGNALING, TGF_BETA_SIGNALING, P53_PATHWAY, TNFA_SIGNALING_VIA_NFKB, etc.) had significant differences (Fig. 7G, H), which might be the underlying mechanism that leads to the different prognosis.
Fig. 7
Identification of differential expressed genes and pathway identification in different clusters. A, B Volcano and heat maps of DEGs in different clusters; KEGG analysis of up-regulated (C) and down-regulated (D) DEGs. GO analysis of up-regulated (E) and down-regulated (F) DEGs. G, H GSEA analysis of DEGs
3.7 Exploration of the landscape of mutations and immune infiltration in different clustersThrough somatic mutation maps, this study found significant differences in gene mutations of AIG-high risk (C1) and AIG-low risk cluster (C2). The top 10 mutant genes of AIG-high risk cluster were TP53, TTN, KMT2D, MUC16, KDM6A, ARID1A, PIK3CA, KMT2C, SYNE1 and ZFHX4 (Fig. 8A). The AIG-low risk cluster were TP53, TTN, KMT2D, KDM6A, MUC16, ARID1A, SYNE1, RB1, HMCN1 and PIK3CA (Fig. 8B). Among them, the mutation rate of hub genes (TP53, TTN) was significantly lower in the AIG-high risk cluster than AIG-low risk cluster. Subsequently, based on the possibility that AIGs may regulate immune response activation or silencing, the tumor immune microenvironment was assessed by xCELL, TIMER, quanTIseq, CIBERSORT and EPIC algorithms under different clusters, and neutrophils were found to be significantly high in the AIG-high risk cluster (C1) in multiple algorithms (Fig. 9A–E), and CD4 T cells, CD8 T cells and DC cells, among others, differed in the different clusters (Fig. 9A–E).
Fig. 8
Gene mutations in different clusters. Visualization of the first 10 mutated genes in the AIGs clusters, A AIGs-high group; B AIGs-low group
Fig. 9
Immune infiltration landscape under five algorithms in different clusters. A xCELL algorithm; B TIMER algorithm; C quanTIseq algorithm; D CIBERSORT algorithm; E EPIC algorithm. C1 is the AIGs-high group and C2 is the AIGs-low group
3.8 Verification of IL7 and NFKB1 in BCa by cell phenotype and immunohistochemistryImmunohistochemical experiments of bladder cancer patients in our medical center showed that both IL7 and NFKB1 were underexpressed in tumor tissues (Fig. 10A, B), and the conclusion was consistent with the above bioinformatics analysis results. Then, the relative wound healing rate of cells was found to be significantly lower after OE-IL7 and OE-NFKB1 by cell scratch assay (Fig. 10C). CCK8 experiments suggested weakened cell proliferation after OE-IL7 and OE-NFKB1 (Fig. 10D). In addition, the number of cell clones decreased after OE-IL7 and OE-NFKB1, compared with the NC group (Fig. 10E). Moreover, the cell migration ability was decreased after OE-IL7 and OE-NFKB1 (Fig. 10F). Collectively, the above studies further confirmed IL7 and NFKB1 as anti-oncogenes in BCa.
Fig. 10
IL7 and NFKB1 were proved to influence bladder cancer progression. A IL7 immunohistochemistry; B NFKB1 immunohistochemistry; C Scratch tests; D CCK8 assay; E clone formation assay; F transwell assay
留言 (0)