
記住我



Tph2 deficiency in rats caused severe growth retardation, together with moderate impairments in somatosensory reflexes and thermoregulatory capabilities, partially aggravated by CN, as compared to SN. Developmental milestones and somatosensory reflexes were assessed every second day during the first two weeks of life (Fig. 2A). This included the assessment of body weight gain (Fig. 2B) and thermoregulatory capabilities (Fig. 2C). Throughout the first two weeks of life, body weight gain was strongly affected by genotype (genotype: F2,82 = 101.743; p < 0.001; age × genotype: F2.865,117.462 = 157.843; p < 0.001). Specifically, body weight increased substantially in Tph2+/− pups and Tph2+/+ littermate controls, while body weight gain was reduced in Tph2−/− pups. Whereas there was no genotype difference on P2, differences in body weight started to emerge from P4, resulting in body weight values of Tph2−/− pups being 20–40% lower by P14 than those observed in Tph2+/+ littermates (Fig. 2D,D′). Surprisingly, the growth retardation phenotype displayed by Tph2−/− pups was aggravated by CN, as reflected in even further reduced body weight gain in Tph2−/− pups exposed to CN, as compared to SN (nesting × genotype: F2,82 = 5.094; p = 0.008; age × nesting × genotype: F2.865,117.462 = 6.563; p < 0.001). Moreover, the aggravated growth retardation phenotype in Tph2−/− pups exposed to CN is in contrast to the increased body weight gain seen in Tph2+/− pups and Tph2+/+ littermates exposed to CN, possibly reflecting higher levels of competition between pups under CN, as compared to SN. Of note, a similar pattern was obtained irrespective of whether body weight values were expressed as percentages relative to all Tph2+/+ littermates or separately for Tph2+/+ littermates exposed to SN versus CN, respectively.
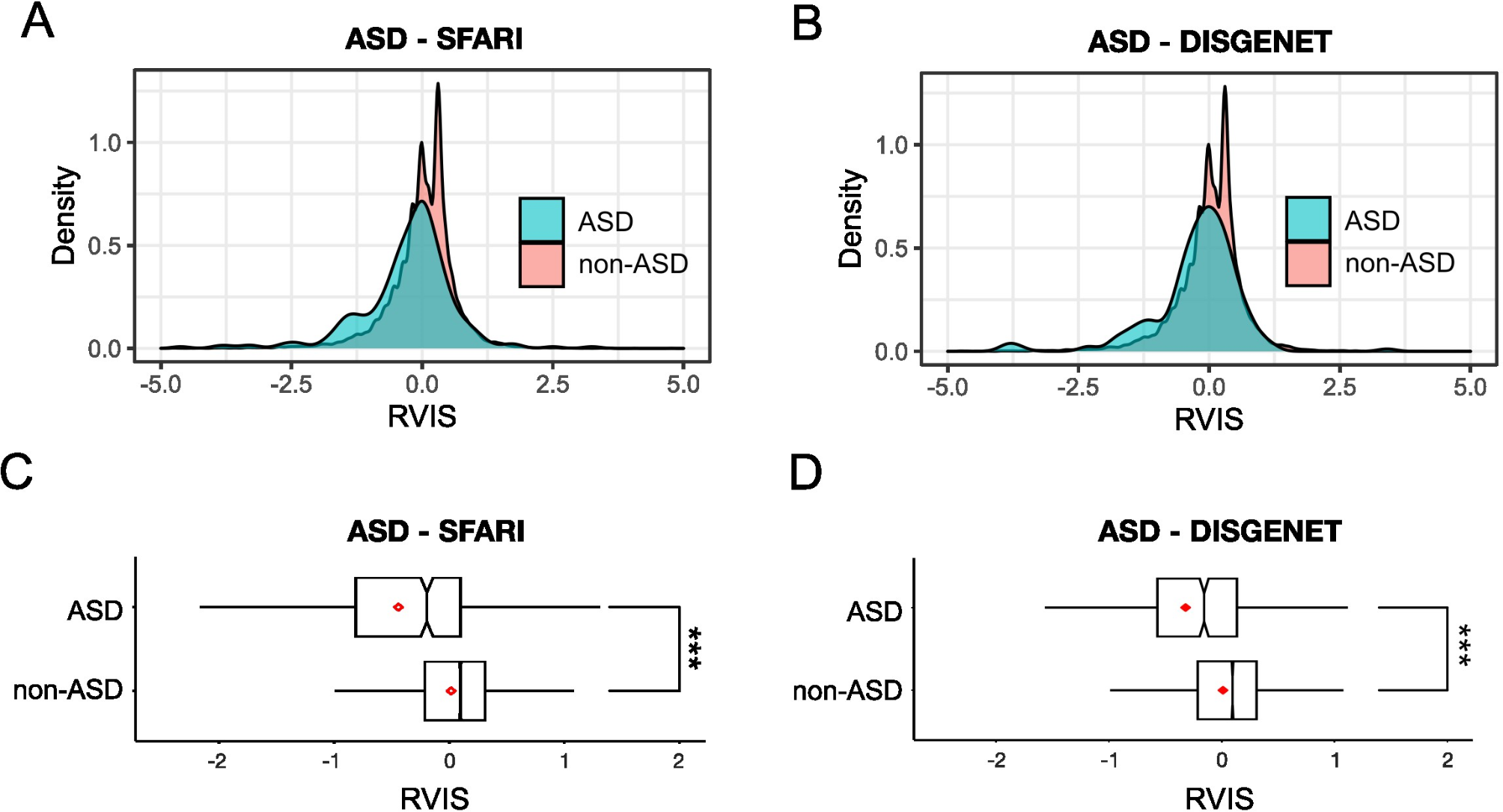
Fig. 2
Tph2 deficiency in rats causes severe growth retardation, together with moderate impairments in somatosensory reflexes and thermoregulatory capabilities, partially aggravated by communal nesting (CN), as compared to standard nesting (SN). (A) Overview of the assessment of developmental milestones, somatosensory reflexes, and thermoregulatory capabilities in Tph2−/− (KO) and Tph2+/− (HET) rat pups, as compared to Tph2+/+ (WT) littermate controls. (B) Exemplary image of WT and KO rat pups on P10. (C) Exemplary thermal image of WT, HET, and KO across P2 to P14 obtained during body temperature measurements. (D) Absolute body weight of WT, HET, and KO rat pups across P2 to P14, depending on nesting condition. (D′) Normalized body weight of HET and KO rat pups across P2 to P14 relative to WT littermate controls, depending on nesting condition. (E) Body temperature before the 10-min isolation period for WT, HET, and KO rat pups across P2 to P14, depending on nesting condition. (F) Body temperature after the 10-min isolation period for WT, HET, and KO rat pups across P2 to P14, depending on nesting condition. (G) Body temperature change during the 10-min isolation period for WT, HET, and KO across P2 to P14, depending on nesting condition. (H) Upright latency in the negative geotaxis assay for WT, HET, and KO rat pups across P2 to P14, depending on nesting condition. SN = standard nesting (blue), CN = communal nesting (orange). P = postnatal day. Data are expressed as mean ± SEM. Effect of genotype: ***(black) WT vs. KO p < .001; **(black) WT vs. KO p < .01. ***(gray) HET vs. KO p < .001; *(gray) HET vs. KO p < .01; *(gray) HET vs. KO p < .05. Effect of nesting: + + + p < .001; + + p < .01; + p < .05. Interaction nesting x genotype: @@ p < .01; @ p < .05. N(WT-SN) = 16, N(HET-SN) = 18, N(KO-SN) = 19, N(WT-CN) = 12, N(HET-CN) = 12, N(KO-CN) = 17
Growth retardation in Tph2−/− pups was associated with impairments in thermoregulatory capabilities. Body temperature was assessed before and after a 10-min social isolation period under room temperature. When measuring body temperature right after removal from the nest before isolation, prominent genotype differences were evident, with Tph2−/− pups having lower body temperature values than Tph2+/− pups and Tph2+/+ littermate controls (genotype: F2,82 = 21.851; p < 0.001; Fig. 2E). The developmental pattern was similar to body weight gain (age × genotype: F10.366,424.997 = 1.906; p = 0.040). While no genotype differences were seen on P2, body temperature differences were evident during later stages of development. However, in contrast to growth retardation, effects of Tph2 deficiency on thermoregulatory capabilities were not aggravated by CN, as compared to SN (nesting × genotype: F2,82 = 0.193; p = 0.825; age × nesting × genotype: F10.366,424.997 = 0.894; p = 0.542). A similar result pattern was found after the 10-min isolation period, with prominent genotype differences in body temperature starting at P4 (genotype: F2,80 = 45.522; p < 0.001; age × genotype: F9.315,372.605 = 4.940; p < 0.001), irrespective of CN (nesting × genotype: F2,80 = 0.163; p = 0.850; age × nesting × genotype: F9.315,372.605 = 0.794; p = 0.626; Fig. 2F). Comparing the body temperature before and after the 10-min isolation period showed that rat pups typically lost between 3 °C to 6 °C on P2, while the typical temperature loss on P14 was less than 1 °C, reflecting improved thermoregulatory capabilities with age. Improved thermoregulatory capabilities with age were seen irrespective of genotype and CN (genotype: F2,80 = 0.695; p = 0.502; age × genotype: F8.715,348.587 = 1.150; p = 0.327; nesting × genotype: F2,80 = 0.397; p = 0.673; age × nesting × genotype: F8.715,348.587 = 0.674; p = 0.727), suggesting that the genotype differences in body temperature evident before isolation were maintained but not exacerbated throughout isolation.
Growth retardation in Tph2−/− pups was further associated with delays in several developmental milestones (Table 1). Firstly, growth retardation was not only reflected in body weight but also body length (genotype: genotype: F2,82 = 35.864; p < 0.001; age × genotype: F7.685,315.088 = 73.624; p < 0.001). Moreover, eye opening was delayed in Tph2−/− pups (genotype: F2,82 = 7.905; p = 0.001; age × genotype: F12,492 = 6.579; p < 0.001). The visibility of a milk spot in the belly was likewise affected by genotype, with less visible milk spots in Tph2−/− pups, as compared to Tph2+/− pups and Tph2+/+ littermate controls, particularly during the first week of life (genotype: F2,82 = 2.329; p = 0.104; age × genotype: F8.171,335.031 = 4.007; p < 0.001). However, those phenotypes were not modulated by CN (all p-values > 0.50). Pinnae detachment, incisor eruption, and fur development were not affected by genotype (all p-values > 0.50).
Table 1 Developmental milestonesFinally, the developmental delay displayed by Tph2−/− pups was associated with a broad variety of alterations in somatosensory reflexes (Table 2). Firstly, the grasping reflex was found to be more prominent in Tph2−/− pups than in Tph2+/− pups and Tph2+/+ littermate controls (genotype: F2,82 = 8.046; p = 0.001; age × genotype: F6.982,286.243 = 3.902; p < 0.001). Secondly, forelimb placing was reduced in Tph2−/− pups (genotype: F2,82 = 131.532; p < 0.001; age × genotype: F10.137,415.624 = 4.782; p < 0.001; Supplementary Fig. 1A). Thirdly, the righting reflex was affected by genotype, with Tph2−/− pups being able to flip over onto the abdomen faster, particularly during the first week of life, possibly because of their lower body weight (genotype: F2,82 = 3.082; p = 0.051; age × genotype: F6.286,257.712 = 2.343; p = 0.030; Supplementary Fig. 1B). Fourthly, level screen was slightly impaired in Tph2−/− pups during the first week of life (genotype: F2,82 = 0.160; p = 0.852; age × genotype: F8.697,356.580 = 3.765; p < 0.001; Supplementary Fig. 1C). Fifthly, negative geotaxis was impaired in Tph2−/− pups. While all genotypes started to turn around on a 45° grid to reach an upright position in the second week of life, there was evidence for a regression in Tph2−/− pups on P12 and P14 (genotype: F2,82 = 2.109; p = 0.128; age × genotype: F7.053,289.161 = 11.043; p < 0.001; Fig. 2H). Sixthly, an increase in vertical screen behavior was observed in Tph2−/− pups (genotype: F2,82 = 7.281; p = 0.001; age × genotype: F7.968,326.676 = 1.798; p = 0.077; Supplementary Fig. 1D). Seventhly, Tph2−/− pups needed more time to withdraw from an elevated edge in the cliff avoidance task during the second week of life (genotype: F2,82 = 0.681; p = 0.509; age × genotype: F10.595,434.396 = 2.118; p = 0.020). Finally, Tph2−/− pups were less able to hold onto a bar resulting in a reduced bar holding time, especially during the second week of life (genotype: F2,82 = 6.745; p = 0.002; age × genotype: F7.282,298.548 = 2.676; p = 0.010). However, except for forelimb placing, alterations in somatosensory reflexes were not modulated by CN (nesting × genotype: F2,82 = 4.830; p = 0.010; all other p-values > 0.50). Sex had no prominent modulatory effects on the alterations caused by Tph2 deficiency in developmental milestones, somatosensory reflexes, or thermoregulatory capabilities, except for grasping reflex and forelimb placing (not shown in detail).
Table 2 Somatosensory reflexesTaken together, Tph2 deficiency led to severe growth retardation, as reflected in reduced body weight gain. Moreover, Tph2 deficiency was associated with impairments in thermoregulatory capabilities and a delay in developmental milestones, most notably eye opening. While CN aggravated the growth retardation phenotype, it did not aggravate the impairments in thermoregulatory capabilities nor the delay in developmental milestones associated with Tph2 deficiency. The effects of Tph2 deficiency on somatosensory reflexes were moderate and mostly not modulated by CN.
Isolation-induced pup ultrasonic vocalizationsTph2 deficiency in rats led to severe deficits in socio-affective communication during the first two weeks of life, as evidenced by reduced call emission rates, regardless of CN. Socio-affective communication in pups was assessed through isolation-induced USV during the 10-min isolation period every second day (Fig. 3A; for exemplary spectrograms, please see Fig. 3B). The average number of calls emitted over the 7 test days was reduced in Tph2−/− pups, as compared Tph2+/− pups and Tph2+/+ littermate controls. This reduction was seen irrespective of CN (genotype: F2,82 = 198.971; p < 0.001; nesting × genotype: F2,82 = 0.334; p = 0.717; Fig. 3C). While Tph2+/− pups and Tph2+/+ littermates emitted ~ 1500–1600 isolation-induced USV on average under SN conditions, Tph2−/− pups emitted only ~ 800–900 isolation-induced USV. This strong reduction of ~ 50% in Tph2−/− pups was also seen under CN conditions. Whereas isolation-induced USV emission rates again ranged between ~ 1500–1600 in Tph2+/− pups and Tph2+/+ littermates, CN did not ameliorate the socio-affective communication deficit displayed by Tph2−/− pups and their emission rate remained at ~ 800–900 isolation-induced USV.
Fig. 3
Tph2 deficiency in rats leads to severe deficits in socio-affective communication during the first two weeks of life, as evidenced by reduced call emission rates. (A) Overview of the assessment of isolation-induced ultrasonic vocalizations in Tph2−/− (KO) and Tph2+/− (HET) rat pups, as compared to Tph2+/+ (WT) littermate controls. (B) Exemplary spectrograms of isolation-induced ultrasonic vocalizations emitted by WT, HET, and KO on P12, depending on nesting condition. (C) Average number of isolation-induced ultrasonic vocalizations emitted across P2 to P14. (D) Average total calling time across P2 to P14. Developmental trajectories of (C′) the number of isolation-induced ultrasonic vocalizations and (D′) total calling time across P2 to P14. SN = standard nesting (blue), CN = communal nesting (orange). P = postnatal day. Data are expressed as mean ± SEM. Effect of genotype: ***(black) WT vs. KO p < .001; **(black) WT vs. KO p < .01. ***(gray) HET vs. KO p < .001; Effect of nesting: + p < .05. N(WT-SN) = 16, N(HET-SN) = 18, N(KO-SN) = 19, N(WT-CN) = 12, N(HET-CN) = 12, N(KO-CN) = 17
When analyzing the developmental trajectories, it was evident that this phenotype emerged around P2. While isolation-induced USV emission rates increased substantially in Tph2+/− pups and Tph2+/+ littermate controls across development from ~ 700–900 USV on P2 to ~ 1700–2200 USV on P14, there was no such prominent increase seen in Tph2−/− pups. Isolation-induced USV emission increased only slightly from ~ 600–700 USV on P2 to ~ 800–1100 USV on P14. This difference in the developmental trajectories resulted in very prominent genotype differences in isolation-induced USV emission rates in the second week of life (age × genotype: F8.778,359.913 = 10.869; p < 0.001). CN did not affect the strong effect of Tph2 deficiency on developmental trajectories (age × nesting × genotype: F8.778,359.913 = 1.112; p = 0.354; Fig. 3C′). In correspondence with the number of calls, total calling time showed a similar pattern. Across the 7 test days, Tph2−/− pups spent less time calling than Tph2+/− pups and Tph2+/+ littermates (genotype: F2,82 = 170.602; p < 0.001; nesting × genotype: F2,82 = 0.113; p = 0.893; Fig. 3D). Starting from P4, total calling time was lower in Tph2−/− pups, as compared to Tph2+/− pups and Tph2+/+ littermates. Similar to call numbers, most prominent genotype differences were seen during the second week of life (age × genotype: F9.019,359.797 = 6.318; p < 0.001; age × nesting × genotype: F9.019,359.797 = 1.020; p = 0.424; Fig. 3D′). Of note, CN did not affect isolation-induced USV emission rates and total calling time (all p-values > 0.50). Sex had a mild modulatory effect on the alterations caused by Tph2 deficiency in isolation-induced USV emission rates and total calling time, with stronger reductions in male than female Tph2−/− pups (not shown in detail).
The deficits in socio-affective communication driven by Tph2 deficiency in rats were also reflected in alterations of acoustic features of USV emitted by Tph2−/− pups. This included all four main acoustic features, i.e. call duration, peak frequency, peak amplitude, and frequency modulation (Fig. 4A). Tph2−/− pups emitted slightly longer isolation-induced USV than Tph2+/− pups and Tph2+/+ littermate controls, irrespective of CN (genotype: F2,82 = 3.209; p = 0.046; age × genotype: F9.157,375.450 = 0.943; p = 0.489; nesting × genotype: F2,82 = 0.326; p = 0.722; age × nesting × genotype: F9.157,375.450 = 0.763; p = 0.653; Fig. 4B, B′). Moreover, isolation-induced USV emitted by Tph2−/− pups were characterized by higher peak frequency under SN and CN conditions (genotype: F2,82 = 18.358; p < 0.001; nesting × genotype: F2,82 = 0.895; p = 0.413; Fig. 4C). This genotype difference in peak frequency was driven by a prominent alteration in the developmental trajectories. While there was a continuous gradual decrease in peak frequency seen across development in Tph2+/− pups and Tph2+/+ littermates, a U-shaped developmental pattern was evident in Tph2−/− pups. In the latter, isolation-induced USV with relatively low peak frequencies were seen during the first week of life similar to Tph2+/− pups and Tph2+/+ littermates, yet a rather abrupt increase in peak frequency occurred around P10 in Tph2−/− pups. Due to a further increase in Tph2−/− pups on P12 and P14, genotype differences in peak frequencies reached up to 8 kHz on P14 (age × genotype: F8.847,362.724 = 18.412; p < 0.001; age × nesting × genotype: F8.847,362.724 = 0.477; p = 0.887; Fig. 4C′). In contrast to peak frequency, Tph2−/− pups emitted isolation-induced USV characterized by lower peak amplitudes, as compared to Tph2+/− pups and Tph2+/+ littermates (genotype: F2,82 = 15.716; p < 0.001; nesting × genotype: F2,82 = 0.557; p = 0.575; Fig. 4D). Genotype differences started to emerge from around P6 and prominent differences were evident throughout the second week of life (age × genotype: F8.565,351.175 = 3.137; p = 0.001; age × nesting × genotype: F8.565,351.175 = 0.766; p = 0.642; Fig. 4D′). Finally, frequency modulation of isolation-induced USV emitted by Tph2−/− pups was reduced, regardless of CN (genotype: F2,82 = 14.497; p < 0.001; nesting × genotype: F2,82 = 1.179; p = 0.313; Fig. 4E). The phenotype started to emerge around P4 and persisted until P14 (age × genotype: F6.394,262.145 = 4.389; p < 0.001; age × nesting × genotype: F6.394,262.145 = 1.158; p = 0.329; Fig. 4E′). Of note, CN led to genotype-independent alterations in the acoustic features of isolation-induced USV. Specifically, reductions in peak frequency (nesting: F1,82 = 9.356; p = 0.003; age × nesting: F4.423,362.724 = 0.757; p = 0.566; Fig. 4C′) and frequency modulation (nesting: F1,82 = 8.002; p = 0.006; age × nesting: F3.197,262.145 = 3.958; p = 0.007; Fig. 4E′) were seen in the pups exposed to CN, particularly in the later phase of development (all other p-values > 0.50). Sex had a mild modulatory effect on the alterations caused by Tph2 deficiency in peak frequency, with more prominent genotype differences in male than female Tph2−/− pups under CN but not SN conditions (not shown in detail).
Fig. 4
Tph2 deficiency in rats is results in changes in the acoustic features of isolation-induced ultrasonic vocalizations. (A) Overview of the acoustic features of isolation-induced ultrasonic vocalizations assessed in Tph2−/− (KO) and Tph2+/− (HET) rat pups, as compared to Tph2+/+ (WT) littermate controls: call duration, peak frequency, peak amplitude, and frequency modulation. (B) Average call duration across P2 to P14. (C) Average peak frequency across P2 to P14. (D) Average peak amplitude across P2 to P14. (E) Average frequency modulation across P2 to P14. Developmental trajectories of (B′) call duration, (C′) peak frequency, (D′) peak amplitude, and (E′) frequency modulation of isolation-induced ultrasonic vocalizations emitted across P2 to P14. SN = standard nesting (blue), CN = communal nesting (orange). P = postnatal day. Data are expressed as means ± SEM. Effect of genotype: ***(black) WT vs. KO p < .001; **(black) WT vs. KO p < .01; *(black) WT vs. KO p < .05. ***(gray) HET vs. KO p < .001; **(gray) HET vs. KO p < .01; *(gray) HET vs. KO p < .05. Effect of nesting: + + + p < .001; + p < .05. N(WT-SN) = 16, N(HET-SN) = 18, N(KO-SN) = 19, N(WT-CN) = 12, N(HET-CN) = 12, N(KO-CN) = 17
Moreover, Tph2 deficiency in rats affected call clustering. Detailed spectrographic analyses of more than 150,000 individual isolation-induced USV on P12 revealed multiple clusters of call subtypes. In Tph2+/+ littermate controls, two call clusters were revealed by plotting peak frequency against call duration. One prominent cluster was characterized by relatively low peak frequencies roughly between 40 and 50 kHz. A second cluster was characterized by relatively high peak frequencies roughly between 60 and 70 kHz. Call durations in the low-frequency cluster were much longer than in the high-frequency cluster. Although both clusters were present in Tph2−/− pups, the typical peak frequency associated with each of the two clusters was shifted up by ~ 5 kHz. Moreover, the relative prevalence of the two call clusters was affected by genotype. While the low-frequency cluster was much more prominent than the high-frequency cluster in Tph2+/+ littermates, this difference in relative prevalence was smaller in Tph2−/− pups due to a more prominent high-frequency cluster, ranging roughly from 65 to 85 kHz. As in Tph2+/+ littermates, call durations in the low-frequency cluster were much longer than in the high-frequency cluster. Call clustering in Tph2+/− pups was more similar to Tph2+/+ littermates than Tph2−/− pups. CN slightly reduced the prevalence of the high-frequency cluster (Fig. 5A-F).
Fig. 5
Tph2 deficiency in rats is associated with changes in the clustering of subtypes of isolation-induced ultrasonic vocalizations. (A-L) Density plots depicting the distribution of individual isolation-induced ultrasonic vocalizations in Tph2−/− (KO) and Tph2+/− (HET) rat pups, as compared to Tph2+/+ (WT) littermate controls, depending on nesting condition, i.e. WT-SN (A, G; ~ 35,000 calls), WT-CN (D, J; ~ 24,000 calls), HET-SN (B, H; ~ 34,000 calls), HET-CN (E, K; ~ 25,000 calls), KO-SN (C, I; ~ 20,000 calls), and KO-CN (F, L; ~ 14,000 calls). SN = standard nesting (blue), CN = communal nesting (orange). Color coding reflects frequencies as percentages. Individual isolation-induced ultrasonic vocalizations depicted here were recorded on postnatal day 12
When plotting peak frequency against frequency modulation, four call clusters were revealed in Tph2+/+ littermate controls. For the low-frequency cluster, two call subtypes were present, differing in frequency modulation. Frequency modulation was either below ~ 15 kHz or above ~ 20 kHz, ranging up to ~ 35 kHz. For the high-frequency cluster, another two call subtypes were present, again differing in frequency modulation. Frequency modulation was either below ~ 15 kHz or above ~ 25 kHz, ranging up to ~ 35 kHz. The low-frequency cluster characterized by low levels of frequency modulation was most prevalent. The high-frequency cluster characterized by high levels of frequency modulation was least prevalent. While all four call clusters were present in Tph2−/− pups, the two call clusters characterized by high levels of frequency modulation were less prevalent than in Tph2+/+ littermates. In particular, the high-frequency cluster characterized by high levels of frequency modulation was almost absent. This indicates that the increased prevalence of the high-frequency cluster in Tph2−/− pups compared to Tph2+/+ littermates was exclusively driven by isolation-induced USV characterized by low levels of frequency modulation, i.e. below 15 kHz. Again, call clustering in Tph2+/− pups was more similar to Tph2+/+ littermates than Tph2−/− pups. CN slightly reduced the prevalence of the high-frequency clusters, with the high-frequency cluster characterized by high levels of frequency modulation being virtually absent in Tph2−/− pups exposed to CN (Fig. 5G-L).
Finally, Tph2 deficiency affected the temporal organization of isolation-induced USV emission. This is suggested by sequential analyses through correlating the call durations of given isolation-induced USV with the call durations of previous ones. In general, correlations between a given isolation-induced USV were highest with the previous one (N-1) and gradually declined when correlating a given isolation-induced USV with the one two before (N-2) or even three before (N-3). This pattern was evident in all genotypes. In Tph2+/+ littermate controls, however, such correlations decreased across developmental stages from P4 over P8 to P12, with higher variability at later stages possibly reflecting a higher level of temporal organization or complexity. This was seen for all three types of correlations (N-1, N-2, and N-3). Interestingly, no such change was seen in Tph2−/− pups. There, the correlations remained mostly unchanged from P4 over P8 to P12, with evidence for a mild inverted U-shaped pattern. This indicates that the temporal call emission pattern remains highly stereotypic even at later developmental stages, with values on P12 being similar to P4, lacking of the increase in temporal organization or complexity seen in Tph2+/+ littermates. Tph2+/− pups displayed an intermediate phenotype for N-1 (age × genotype: F3.715,152.33 = 5.838; p < 0.001; Fig. 6A), N-2 (age × genotype: F3.358,137.698 = 4.09; p = 0.006; Fig. 6B), and N-3 (age × genotype: F3.171,129.992 = 4.133; p = 0.007; Fig. 6C). Importantly, the highly stereotypic temporal call emission pattern displayed by Tph2−/− pups was primarily seen under conditions of close temporal proximity (N-1 and N-2), while under conditions of low temporal proximity (N-3) correlations were lower than in Tph2+/+ littermates (genotype: F2,82 = 6.81; p = 0.002; Fig. 6C). Of note, similar results were obtained for SN and CN conditions, with developmental changes being slightly more prominent under SN conditions (not shown in detail).
Fig. 6
Tph2 deficiency in rats affects the temporal organization of the emission of isolation-induced ultrasonic vocalizations. (A-C) Correlation coefficients (r) between call durations reflecting the sequential organization of isolation-induced ultrasonic vocalizations in Tph2−/− (KO) and Tph2+/− (HET) rat pups, compared to Tph2+/+ (WT) littermate controls. Correlation coefficients (r) indicate the level of correlations between the durations of a given isolation-induced ultrasonic vocalizations with the call durations of the previous ones (A, N-1), the ones two before (B, N-2), and the ones three before (C, N-3) on postnatal days (P) 4, 8, and 12. SN = standard nesting (blue), CN = communal nesting (orange). Data are presented as mean ± SEM. Effect of age: *** p < .001; ** p < .01: * p < .05
Taken together, Tph2 deficiency led to a prominent reduction in the emission of isolation-induced USV. Moreover, Tph2 deficiency was associated with acoustically-altered isolation-induced USV characterized by untypically long call durations and high peak frequencies but low peak amplitudes together with low levels of frequency modulation. Call clustering and the temporal organization of isolation-induced USV were also affected by Tph2 deficiency, with differences in the relative prevalence of call clusters being less prominent and the sequential call emission pattern being more stereotypic in Tph2-deficient rat pups. The effects of Tph2 deficiency on isolation-induced USV were mostly not modulated by CN.
Homing test as a proxy maternal affiliationTph2 deficiency in rats resulted in severely impaired maternal affiliation, as assessed by the pup’s preference for its mother’s odor in the homing test (Fig. 7A). This impairment was ameliorated by CN. As expected, the large majority of rat pups displayed a strong preference for the one third of the test apparatus with soiled bedding containing the mother’s odor over the one third of the test apparatus with clean bedding without the mother’s odor (zone: F2,81 = 890.722; p < 0.001; Fig. 7B). This preference was modulated by Tph2 deficiency, with Tph2+/− pups and Tph2+/+ littermate controls but not Tph2−/− pups displaying strong preferences for the zone with soiled bedding containing the mother’s odor (genotype: F2,81 = 72.631; p < 0.001; zone × genotype: F2,81 = 130.372; p < 0.001). Importantly, this preference was affected by CN (zone × nesting: F1,81 = 5.052; p = 0.027; zone × nesting × genotype: F2,81 = 2.051; p = 0.135). Specifically, under SN conditions, Tph2+/− pups and Tph2+/+ littermates displayed a prominent preference for soiled bedding, while no preference was seen in Tph2−/− pups. While Tph2+/− pups and Tph2+/+ littermates spent ~ 91% and ~ 88% of time on average, respectively, in the one third of the test apparatus with soiled bedding, Tph2−/− pups spent only ~ 39% of time on average in soiled bedding, i.e. only slightly above chance level of 33%. CN exerted beneficial effects in enhancing maternal affiliation, as reflected in a stronger preference for soiled bedding. Beneficial effects of CN were primarily evident in Tph2−/− pups, leading to ~ 51% of time spent in soiled bedding, i.e. clearly above chance level of 33%, reflecting a prominent increase of ~ 12%, as compared to SN. In contrast, the beneficial effects of CN on Tph2+/− pups and Tph2+/+ littermates were mild, likely due to the strong preference already shown under SN conditions. Tph2+/− pups and Tph2+/+ littermates spent ~ 92% and ~ 93% of time on average, respectively, in the one third of the test apparatus with soiled bedding, reflecting an increase of merely 1% and 5%, respectively, as compared to SN. Of note, no evidence for genotype differences were obtained for line crossing (genotype: F2,82 = 1.544; p = 0.220; nesting: F1,82 = 2.456; p = 0.121; nesting × genotype: F2,82 = 0.506; p = 0.605), suggesting that the maternal affiliation deficits displayed by Tph2−/− pups were not due to locomotor impairment (Fig. 7C).
Fig. 7
Tph2 deficiency in rats leads to maternal affiliation deficits, which can be rescued through communal nesting (CN), as compared to standard nesting (SN). (A) Overview of the homing test as a proxy for maternal affiliation in in Tph2−/− (KO) and Tph2+/− (HET) rat pups, compared to Tph2+/+ (WT) littermate controls. (B) Maternal affiliation as measured through the time spent in the clean bedding zone versus the soiled bedding zone containing maternal odors, depending on nesting condition. (C) Total number of line crossings, depending on nesting condition. (D) Total number of isolation-induced ultrasonic vocalizations, depending on nesting condition. (E) Total calling time, depending on nesting condition. SN = standard nesting (blue), CN = communal nesting (orange). Data are expressed as mean ± SEM. Effect of genotype: *** p < .001; Effect of zone: ### p < .001, ## p < .01. N(WT-SN) = 16, N(HET-SN) = 18, N(KO-SN) = 19, N(WT-CN) = 12, N(HET-CN) = 12, N(KO-CN) = 17
Consistent with the reduction in isolation-induced USV, USV emission rates during the homing test were affected by genotype. Tph2−/− pups again displayed a reduced rate of isolation-induced USV emission, as compared to Tph2+/− pups and Tph2+/+ littermate controls (genotype: F2,82 = 9.915; p < 0.001; nesting × genotype: F2,82 = 1.334; p = 0.269; Fig. 7D), suggesting a robust phenotype independent of test context. However, total calling time was found to be comparable to the levels seen in Tph2+/− pups and Tph2+/+ littermates (genotype: F2,82 = 1.943; p = 0.150; nesting × genotype: F2,82 = 1.297; p = 0.279; Fig. 7E). This may indicate that the homing test environment caused alterations in acoustic features, as compared to the test environment used for studying the developmental trajectories of isolation-induced USV.
Taken together, Tph2 deficiency led to severely impaired maternal affiliation, yet this impairment was ameliorated by CN.
Maternal preference testIn order to close the communicative loop between mother and pup, maternal preference was assessed by exposing one Tph2+/+ littermate control and one Tph2−/− pup simultaneously to their mother (Fig. 8A). Maternal preference was depending on the genotype of the pup, with mothers showing a preference for Tph2+/+ littermate controls over Tph2−/− pups (pup genotype: F1,22 = 6.721; p = 0.017; Fig. 8B). While CN had no prominent modulatory effect (nesting: F1,22 < 0.001; p = 0.995; nesting × pup genotype: F1,22 = 2.059; p = 0.165), it is worth noting that the mothers displayed a more prominent preference for Tph2+/+ littermates over Tph2−/− pups under CN conditions (t11 = 2.239; p = 0.047) than under SN conditions (t11 = 1.323; p = 0.213).
Fig. 8
Tph2 deficiency in rats causes a reduction in maternal preference, which is emphasized through communal nesting (CN), as compared to standard nesting (SN). (A) Overview of the maternal preference test to close the communicative loop between mother and pup and to compare maternal preferences between Tph2−/− (KO) rat pups and Tph2+/+ (WT) littermate controls. (B) Time spent by the mother in proximity to WT versus KO rat pups. Of note, the individual data points of an individual WT rat pup and an individual KO rat pup are connected by a line in order to indicate that the two connected rat pups were simultaneously exposed to their mother. Mothers were always exposed to their own pups. SN = standard nesting (blue), CN = communal nesting (orange). Data are expressed as mean ± SEM. Effect of pup genotype: *p < .05. N(SN) = 12 WT-KO pairs, N(CN) = 12 WT-KO pairs
Taken together, Tph2 deficiency led to a reduced ability to attract the mother, particularly under CN.
留言 (0)