
記住我

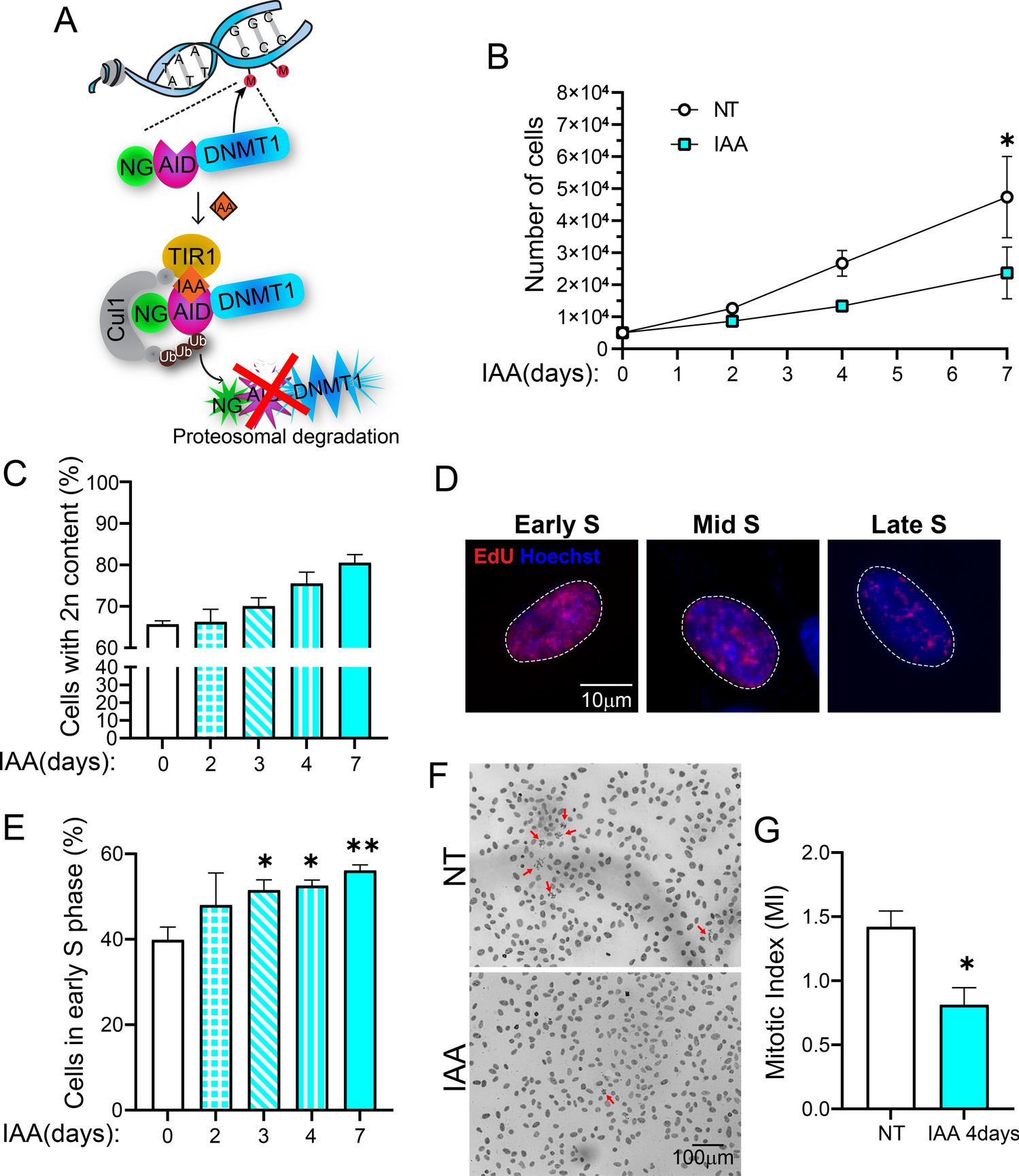






IQGAP1 possesses six functional domains (Fig. 1A) and belongs to a three-member family. To determine the specific isoform expressed in B cells, we used non-overlapping primer sets to amplify and quantify the transcripts that encode Iqgap1, Iqgap2, and Iqgap3 by quantitative PCR in sorted and purified bone marrow-derived total B cells. We found that only the transcripts expressing Iqgap1 were the predominant isoform, and the Iqgap2 and Iqgap3 are undetectable in total B cells (Fig. 1B). Therefore, to establish the role of IQGAP1 in B cell development and function, we used Iqgap1 gene knockout mice in C57BL/6 strain background (Iqgap1−/−) [24]. Earlier genomic and biochemical studies confirmed that this insertion-deletion mutation targeting exons 26 and 27 of the Iqgap1 gene resulted in a null allele [24]. Therefore, we analyzed the expression of the IQGAP1 protein in the wild-type (WT, C57BL/6) and Iqgap1−/− mice. WT B cells contained a 190 kDa corresponding to the size of the full-length IQGAP1 protein, which was absent in Iqgap1−/− B cells (Fig. 1C).
B cell development in the BM of Iqgap1 −/− mice is impairedTo investigate the role of IQGAP1 in B cell development, we analyzed the absolute numbers of B cells in the BM using the pan B cell lineage marker B220 in flow cytometry. The absolute numbers of total lymphocytes in the BM of Iqgap1−/− mice were comparable to that of WT (Fig. 1D, E). Irrespective of this, the absolute numbers of B220+ B cells were significantly increased in the BM of Iqgap1−/− mice compared to that of WT (Iqgap1−/−: 11.74 ± 4.72 × 104; WT: 7.46 ± 2.62 × 104; n = 17, 17; and p = 0.002) (Fig. 1D, E). B220+ B cells can be further divided into B220High and B220Low subsets. Iqgap1−/− mice exhibited significant increases in both B220High (Iqgap1−/−: 10.2 ± 4.5 × 105; WT: 6.4 ± 2.3 × 105; n = 17, 17; and p = 0.003) and B220Low subsets (Iqgap1−/−: 1.55 ± 0.44 × 104; WT: 1.02 ± 0.43 × 104; n = 17, 17; and p = 0.001). Based on B220 and IgM expression, BM B cells are classified into pro/pre-B (B220LowIgM−), immature B cells (B220+IgMLow), and mature/recirculating B (B220HighIgMHigh). The absolute numbers of B220LowIgM− pro/pre (Iqgap1−/−: 6.45 ± 3.0 × 104; WT: 3.94 ± 1.3 × 104; n = 14, 14; and p = 0.008) and recirculating immature B220+IgMLow population (Iqgap1−/−: 3.4 ± 1.5 × 104; WT: 2.17 ± 1.11 × 104; n = 14, 14; and p = 0.022) were significantly increased in Iqgap1−/− mice compared to WT (Fig. 1F, G). In contrast, the number of B220HighIgMHigh mature B cells was comparable between Iqgap1−/− and WT mice (Iqgap1−/−:0.89 ± 0.37 × 104; WT: 0.88 ± 0.52 × 104; n = 14, 14; and p = NS) (Fig. 1G). We further analyzed the expression of CD43 in B220+ B cells, distinguishing B1 B cells from follicular, marginal zone, and immature B cells in the unstimulated adult murine spleen. The absolute B220+CD43+ B cell numbers did not differ significantly between Iqgap1−/− and WT (Iqgap1−/−: 1.6 ± 0.89 × 106 and WT: 1.3 ± 0.52 × 106; n = 9, 9; p = 0.36) (Fig. 1H, I). This indicates the number of B1 B cells does not differ between the Iqgap1−/− and WT mice.
Lack of IQGAP1 forms a blockade at early B cell development, and it is cell-autonomousUsing flow cytometry, we found B220MedCD43− pre-B cells were significantly higher in Iqgap1−/− (Iqgap1−/−: 7.6 ± 3.2 × 106 and WT: 5.0 ± 1.7 × 106; n = 8, 8; p = 0.041), which is consistent with the increased B220LowIgM− pro/pre-B cells populations. As pro/pre-B-I cells mature into pre-B-II (large and small), c-kit expression decreases, and CD25 levels increase. In this context, we also observed a significantly increased B220+CD25+ pre-B-II population (Iqgap1−/−: 4.10 ± 2.16 × 106 and WT: 2.35 ± 1.31 × 104; n = 9, 9; p = 0.05) in Iqgap1−/− mice (Fig. 2A, B). BM-derived cells were also stained with anti-B220, anti-IgM, and anti-IgD Abs and analyzed by flow cytometry. The absolute numbers of IgM+IgD+ (Iqgap1−/−: 0.57 ± 0.25 × 106 and WT: 0.42 ± 0.19 × 106; n = 13, 13; p = NS) and IgM−IgD+ (Iqgap1−/−: 0.48 ± 0.15 × 106 and WT: 0.52 ± 0.25 × 106; n = 13, 13; p = NS) B cells were comparable between Iqgap1−/− and WT mice (Fig. 2C, D). However, the pre-B IgM−IgD− (Iqgap1−/−: 0.68 ± 0.30 × 107 and WT: 0.40 ± 0.13 × 107; n = 13, 13; p = 0.008) and the immature IgM+IgD− (Iqgap1−/−: 3.38 ± 1.68 × 106 and WT: 1.71 ± 0.62 × 106; n = 13, 13; p = 0.003) B cells were increased in Iqgap1−/− mice compared to that of WT (Fig. 2C, D), indicating a loss of developmental regulation during the early stages of B cell maturation.
Fig. 2
Lack of IQGAP1 blocks B cell development at pre-B cell stage in the BM. A, B Absolute numbers of B220+CD25+ large and small pre-B-II stage by flow cytometry. C, D Absolute numbers of B220+IgM−IgD− Pre-B, B220+IgM+IgD− immature B and B220+IgMHighIgD+ or B220+IgMLowIgD+ mature B cells. BM cells were stained with anti-B220, anti-IgM, and anti-IgD antibodies, and the absolute numbers were calculated per million total gated lymphocytes. Data shown in A, C, and E are one representative panel from at least three independent experiments. Open and filled circles in B and D are values obtained from an individual mouse from four independent experiments. Data are shown with the mean ± SEM. Statistical significance was calculated using the Student’s t-test, and p-values are shown below each graph. E Indicated host mice were lethally irradiated and reconstituted with donor-derived 5 × 105 BM cells. Eight weeks later, cells from the host BM were harvested and analyzed. The absolute numbers of reconstituted pro-B cells were identified with anti-B220 and anti-CD43 antibody staining. B220MedCD43+ and B220+CD43− cells were quantified from six host mice, and the average absolute numbers are presented as bar graphs. F Absolute numbers of reconstituted B220+IgM−IgD− Pre-B, B220+IgM+IgD− immature B, and B220+IgMHighIgD+ or B220+IgMLowIgD+ mature B cells. BM cells were stained with anti-B220, anti-IgM, and anti-IgD antibodies, and the absolute numbers were calculated per million total gated lymphocytes. Data shown on the left are representative panels. The average absolute numbers are presented as bar graphs. Data shown are from two independent experiments with three mice each. Data are shown with the mean ± SEM. Statistical significance was calculated using the Student’s t-test, and p-values are shown below each graph
IQGAP1 is expressed in most of the hematopoietic cell lineages [25]. Since IQGAP1 has been globally deleted [24] in the mice we used, the defects we observed in absolute B cell numbers can be cell-intrinsic or due to an abnormality in the stromal microenvironment. To distinguish between these two possibilities, we performed bone marrow (BM) transplantation (WT → WT; Iqgap1−/− → WT; WT → Iqgap1−/−; Iqgap1−/− → Iqgap1−/−). After eight weeks, we analyzed the B cell development in the reconstituted host mice using flow cytometry and found lymphocyte reconstitution was successful in all hosts. Compared to WT, transferring Iqgap1−/− BM cells into WT or Iqgap1−/− hosts resulted in an increased absolute number of lymphocytes in the BM (Supplementary Fig. 1A). Absolute numbers of B220+ B, B220High, and B220Low cell numbers were significantly increased in host mice that received IQGAP1−/− BM cells (Supplementary Fig. 1B). Also, although B220+IgMNeg cells did not show any considerable change, B220+IgMHigh and B220+IgMLow cells were significantly increased in the recipient mice that were transplanted with Iqgap1−/− BM cells compared to that of WT (Supplementary Fig. 1C). Analyzing Pre/Pro-B and Pro-B cells (B220+CD43+) and immature-B and Pre-B cells (B220MedCD43−) in reconstituted mice confirmed the accumulation of immature and Pre-B cells (Fig. 2E). Staining for immature (IgM+IgD−) and mature (IgM+IgD+ and IgM−IgD+) B cell populations in the reconstituted mice indicated a significant increase in the IgM+IgD− and IgM+IgD+ but not the IgM−IgD+ B cells (Fig. 2F). In the spleens from the BM reconstituted mice, we did not observe an increase in the absolute number of lymphocytes or B220+ B cells (Supplementary Fig. 2A). However, the transfer of Iqgap1−/− BM cells into the WT or Iqgap1−/− mice led to a significant increase in the absolute numbers of immature IgM+IgD− and mature IgM+IgD+ but not mature IgM−IgD+ B cell populations (Supplementary Fig. 2B). These results demonstrate that the lack of IQGAP1 in the hematopoietic compartment but not the stromal environment is primarily responsible for the B cell developmental defects.
Lack of IQGAP1 significantly increases IgMHighCD23− T1 B cells in the spleensNext, we analyzed the B cell maturation in the spleen. Single-cell suspensions of unstimulated splenocytes were stained with appropriate markers and were analyzed by flow cytometry. Lack of IQGAP1 did not alter the total lymphocyte numbers in the Iqgap1−/− mice (Iqgap1−/−: 52.11.0 ± 6.7 × 106 and WT: 53.63 ± 8.7 × 106; n = 16, 16; p = NS) (Fig. 3A). Moreover, similar to the BM, the absolute numbers of B220+ B cells in the spleen were moderately but significantly increased in Iqgap1−/− compared to that of the WT mice (Iqgap1−/−: 16.58 ± 6.26 × 106 and WT: 13.17 ± 5.16 × 106; n = 31, 31; p = 0.022) (Fig. 3B). To further analyze the role of IQGAP1 in splenic B cell development, we stained them with anti-IgM and anti-IgD Abs. Although there were moderate increases in IgM+IgD− B cells that contain immature T1 B and mature MZB (Iqgap1−/−: 2.91 ± 2.17 × 106 and WT: 1.85 ± 1.37 × 106; n = 12, 12; p = NS) and mature IgM+IgD+ (Iqgap1−/−: 5.3 ± 2. 95 × 106 and WT: 3.46 ± 2.01 × 106; n = 12, 12; p = NS) or IgM−IgD+ (Iqgap1−/−: 5.08 ± 2.49 × 106 and WT: 3.88 ± 1.82 × 106; n = 12, 12; p = NS) B cells in the spleen of Iqgap1−/− mice, they did not differ significantly (Fig. 3C).
Fig. 3
Lack of IQGAP1 leads to increased T1 B cells in the spleen. A The total number of lymphocytes within the spleens of WT and Iqgap1−/− mice was comparable. Absolute numbers were calculated from live cell gating of the lymphocyte population using flow cytometry. Data shown represent the total lymphocytes per spleen from both strains. B Absolute total splenic B cells were comparable between the WT and Iqgap1−/− mice. Viable cells within the lymphocyte gate were analyzed for the absolute number of total B cells using anti-B220 antibody using flow cytometry. Data shown represent the B220+ B cells per spleen from both strains. C Absolute numbers of immature and mature B cells do not differ between the WT and Iqgap1−/− mice. Total splenic B cells were stained with anti-B220, anti-IgM, and anti-IgD antibodies. Data shown represent the B220+IgM+ immature B/mature MZ B and B220+IgM+IgD+ or B220+IgM−IgD+ mature B cells. Data represent absolute numbers of immature or mature B cells per spleen from both strains. D Absolute numbers of T1-B but not T2-B or T3-B are altered in Iqgap1−/− mice. Total splenocytes were stained with anti-B220 and anti-CD93 antibodies, and the double-positive cells were gated and quantified. E B220+CD93+ B cells were further sub-divided into B220+CD93+IgM+CD23− T1-B, B220+CD93+IgM+CD23+ T2-B, and B220+CD93+IgM−CD23+ T3-B cells. Open and filled circles in A, B, C, and E are values obtained from an individual mouse from four independent experiments. Data are shown with the mean ± SEM. Statistical significance was calculated using the student’s t-test, and p-values are shown below in each graph
Next, we identified the transitional B cells among the B220+ splenocytes based on their CD93 positivity. Transitional T1, T2, and T3 B cells are defined as IgMHighCD23−, IgMHighCD23+, and IgMLowCD23+, respectively. We gated the B220+ splenocytes into CD93+ and CD93− B cells (Fig. 3D). We subdivided the CD93+ B cells based on IgM and CD23 positivity. Total percentages of B220+CD93+ B cells did not vary between the WT and the Iqgap1−/− splenocytes. However, the absolute number of IgMHighCD23− T1 B cells was significantly increased among the splenocytes from Iqgap1−/− mice (Fig. 3E). Absolute T2 and T3 B cell numbers were comparable between the WT and the Iqgap1−/− mice.
Absence of IQGAP1 differentially affects FO B and MZ B cells in the spleensB220+ splenic B cells can also be separated into newly-formed (NF B) (CD21Low/−CD23Low/−), follicular (FO B) (CD23HighCD21Low/−) and marginal zone (MZ B) (CD21HighCD23Low/−) cells. Therefore, we next analyzed the numbers FO, NF, and MZ B cells among splenocytes using flow cytometry (Fig. 4A). The absolute numbers of B220-gated CD21Low/−CD23High FO B (Iqgap1−/−: 11.7 ± 3.5 × 106 and WT: 8.92 ± 3.82 × 106; n = 14, 14; p = 0.026) and the CD21Low/−CD2 3Low/− NF B cells (Iqgap1−/−: 6.37 ± 2.71 × 106 and WT: 4.33 ± 1.18 × 106; n = 14, 14; p = 0.018) were significantly increased in Iqgap1−/− mice. However, the absolute number of MZ B was significantly reduced in the spleens of IQGAP1−/− mice (Iqgap1−/−: 1.18 ± 0.6 × 106 and WT: 1.59 ± 0.87 × 106; n = 14, 14; p = 0.05). In addition to these changes, the B cell follicles in the spleens of Iqgap1−/− mice were extended, and each T cell area (Blue) was surrounded by multiple B cell regions (Red). Compared to this, in the spleens from WT mice, most of the T cell area was primarily encircled by a single B cell follicle. MZ B cells can also be defined by their exclusive high IgM, CD21, and CD1d expression [26]. Staining for IgM and CD21 (Iqgap1−/−: 0.93 ± 0.44 × 106 and WT: 1.56 ± 0.0.53 × 106; n = 11, 11; p = 0.0049) or IgM and CD1d (Iqgap1−/−: 0.75 ± 0.32 × 106 and WT: 1.18 ± 0.31 × 106; n = 9, 9; p = 0.0364) confirmed the significant decrease in the absolute number of MZ B cells (Fig. 4B, C). We performed immunohistochemical staining of spleen cryosections to establish the exclusive reduction in MZ B cells. Iqgap1−/− mice displayed normal B cell follicle architecture (Fig. 4D), as revealed by the anti-CD3ε and anti-B220 mAb staining. Next, we used MOMA-1 mAb to determine the changes in the marginal zone region. The spleen sections from the WT mice revealed a well-defined band of B220+ MZ area outside of the MOMA-1+ metallophilic macrophages. However, the marginal zone regions were greatly reduced and less contiguous in Iqgap1−/− mice, further confirming a reduction in MZ B cells (Fig. 4E).
Fig. 4
Lack of IQGAP1 leads to increased NF B and FO B but decreased MZ B cells. A Absolute numbers of CD21Low/−CD23Low/− NF B, CD21LowCD23High FO B, and CD21HighCD23Low/− MZ B cells are shown. Total splenic B cells were stained with anti-B220, anti-CD21, and anti-CD23 antibodies, and live cells were gated and compared between the WT and Iqgap1−/− mice by flow cytometry. Data shown represent absolute numbers per million live B220+ B cells. B Reduction in MZ B cells was validated using anti-IgM and anti-CD21 antibody staining of B220+ splenic B cells. Data shown represent absolute numbers per million live B220+ B cells. C Reduction in MZ B cells was validated using anti-IgM and anti-CD1d antibody staining of B220+ splenic B cells. Data shown represent absolute numbers per million live B220+ B cells. Open and filled circles in A, B, and C are values obtained from an individual mouse from four independent experiments. Data are shown with the mean ± SEM. Statistical significance was calculated using the student’s t-test, and p-values are shown below in each graph. D, E Immunohistochemical analyses of splenic sections from WT and Iqgap1−/− mice. Spleens were isolated and embedded in paraffin, and 7-micron thick sections were cut and stained for total B cells (anti-B220), T cells (anti-CD3ε), and metallophilic macrophages (anti-MOMA antibody). Panels show the locations of T and B cells as part of the B cell follicles and metalophillic antigen-1+ macrophages surrounding the follicles. F) Indicated host mice were lethally irradiated and reconstituted with donor-derived 5 × 105 BM cells. Eight weeks later, cells from the host BM were harvested and analyzed. Absolute numbers of reconstituted CD21Low/−CD23Low/− NF B, CD21LowCD23High FO B, and CD21HighCD23Low/− MZ B cells are shown. Total splenic B cells were stained with anti-B220, anti-CD21, and anti-CD23 antibodies, and live cells were gated and compared between the WT and Iqgap1−/− mice. Data shown represent absolute numbers per million live B220+ B cells
These reductions were B cell-intrinsic as reconstitution with Iqgap1−/− BM cells either into WT or Iqgap1−/− mice confirmed these defects. In the spleen of the reconstituted mice, we did not observe an increase in the absolute number of lymphocytes or B220+ B cells. However, the transfer of Iqgap1−/− BM cells into the WT or Iqgap1−/− mice led to a significant increase in the absolute numbers of immature (IgM+IgD−) but not mature (IgM+IgD+ and IgM−IgD+) B cell populations. Reconstitution with Iqgap1−/− BM cells either into WT or Iqgap1−/− mice showed a decrease in CD21HiCD23Low/− MZ B cells (Fig. 4F). A decrease in the MZ B cells was further validated by a reduction in IgM+CD21High B cells in the reconstituted mice (Supplementary Fig. 3). Our results demonstrate that the B cell defects in the BM and spleen of Iqgap1−/− mice were cell-intrinsic.
Lack of IQGAP1 impairs T-independent and T-dependent antibody productionThe significant impairment in B cell response in Iqgap1−/− mice could be due to either a developmental or a functional defect. To narrow it down, we first analyzed the basal immunoglobulin levels in the pre-immune sera of WT and Iqgap1−/− mice using enzyme-linked immunosorbent assay (ELISA). Then, we quantified each isotype in the pre-immune sera using an ELISA-based clonotyping assay. Figure 5A shows that IgG1, IgG2a, IgG2b, and IgA levels were comparable between WT and Iqgap1−/− mice. However, the levels of IgM were significantly reduced. (Fig. 5A). Secreted natural IgM is central to neutralizing pathogens and self-antigens. Thus, the presence of multiple Ig isotypes in the sera of Iqgap1−/− mice indicated that the lack of IQGAP1 did not result in a global hypo-responsiveness of B cells. In contrast to IgM, IgG3 was significantly increased in the sera of Iqgap1−/− mice (Fig. 5A). The IgG3 subtype possesses an extended hinge region, superior molecular flexibility, broad polymorphisms, and unique glycosylation sites compared to other IgG subclasses. However, the relevance of increased IgG3 in Iqgap1−/− mice sera is unknown.
Fig. 5
Lack of IQGAP1 impairs T-independent and T-dependent antibody production. A Pre-immune sera were collected from 20 unchallenged WT or Iqgap1−/− mice each and were analyzed for the levels of soluble IgM, IgA, IgG1, IgG2a, IgG2b, and IgG3 using isotype-specific capture and detection antibodies-based ELISA. Data presented were an average of duplicates for each isotype of each serum from multiple experiments. A total of 20 sera were used for each strain. Data are shown with the mean ± SEM. Statistical significance was calculated using the student’s t-test, and p-values are shown below in each graph. B T-independent B cell responses were measured following the TNP-AECM-Ficoll antigen challenge. Four WT or four Iqgap1−/− mice were used to collect pre-immune or immune sera. ELISA was performed with titrated concentrations of sera (1:400, 1:800, and 1:1600). Antigen-specific IgM, IgG1, IgG2a, and IgG3 isotypes were tested and shown. C T-dependent B cell responses were measured following the TNP-KLH antigen challenge. Four WT or four Iqgap1−/− mice were used to collect pre-immune or immune sera. ELISA was performed with sera collected at different time points before (pre-immune) and after the antigen challenge (post 7d, 21d, and following 7d after booster challenge). Antigen-specific IgM, IgG1, IgG2a, and IgG2b isotypes were tested and shown. Open and filled circles in A and B are values obtained from an individual mouse. Black and grey bars represent, respectively. Statistical significance was calculated using student’s t-test, and p-values are shown below each graph
To test whether IQGAP1 plays a role in humoral immune responses, we challenged age and sex-matched WT and Iqgap1−/− mice with T-independent (TI) antigen TNP-AECM-Ficoll (TNP (2,4,6-Trinitrophenyl), where the hapten is conjugated to Amino Ethyl Carboxy Methyl-Ficoll and collected sera to analyze the presence of antigen-specific IgM, IgG1, IgG2a, or IgG3. Our phenotypic analyses indicated increased naïve B cells in Iqgap1−/− mice expressed membrane-bound IgM and IgD in both the BM and spleen. Irrespective of this and an increase in the overall FO B cell number, Iqgap1−/− mice could not produce similar amounts of TNP-specific antibodies during TI responses as WT mice (Fig. 5B). Next, we examined the T-dependent (TD) immune responses by intraperitoneal injection of TNP-KLH. In the primary response, the generation of TNP-specific IgM, IgG1, IgG2a, or IgG2b was significantly reduced in Iqgap1−/− mice) compared to WT mice (Fig. 5C). Further, after the antigen boost, the KLH-specific IgG1 did not recover and remained significantly low in the Iqgap1−/− mice compared to WT (Fig. 5C). These results demonstrate that IQGAP1 plays a crucial role in the activation and functions of B cells.
Receptor editing at κ the locus is moderately defective in B cells lacking IQGAP1The diversity in the length of hypervariable complementarity-determining region 3 (CDR3) of the heavy chain (CDRH3) indicates a repertoire of rearranged VH genes and polyclonality [27]. Using PCR-based spectrotyping, we analyzed the fragment size and peak height for IgHV3, IgHV5, IgHV6, and IgHV7 in B cells from the BM. Our results show that the length of the variable regions and the relative number of sequence-occurrences of the heavy chains in B cells from the WT and Iqgap1−/− mice followed an expected normal Gaussian distribution indicative of diverse and comparable repertoire (Fig. 6A). Thus, the lack of IQGAP1 did not affect the B cell repertoire diversity at the heavy chain rearrangement. Furthermore, the preserved heavy chain rearrangement in B cells from Iqgap1−/− mice indicated that the absence of IQGAP1 did not affect the diversity of B cell repertoire at the heavy chain rearrangement.
Fig. 6
Light chain but not heavy recombination is affected by the lack of IQGAP1. A PCR-based spectrotyping was employed to quantify the diversity in the length of hypervariable CDR3 of the heavy chain (CDRH3) in B cells from the BM of WT and Iqgap1−/− mice. The fragment size and peak height for IgHV3, IgHV5, IgHV6, and IgHV7 indicated that the repertoire of rearrangement VH genes and polyclonality between these two strains are comparable and followed the expected normal Gaussian distribution indicative of diverse and comparable repertoire. B) Igκ and Igλ light chains in the sera are reduced in Iqgap1−/− mice. C RS rearrangement at the κ-locus is reduced in the absence of IQGAP1. Statistical significance was calculated using the student’s t-test, and p-values are shown below in each graph
CD43+ B cell progenitors in mice rearrange immunoglobulin Igκ genes before the assembly of a productive VHDHJH joint. Thus, μ chain rearrangement and expression are not prerequisites to Igκ light chain gene rearrangements in normal development [28]. Therefore, we next analyzed the quantity of κ and λ light chains in the sera of non-challenged healthy WT and Iqgap1−/− mice using an ELISA-based assay. We found a significant reduction in both the κ and λ light chains in Iqgap1−/− mice compared to the WT (Fig. 6B). This demonstrates that the defect we see in the antibody production in challenged mice could be related to the production of the light but not the heavy chains. Indeed, small GTPase, Ras, plays an essential role via Mek1/2 and Erk1/2 in silencing Ccnd3 transcription, exiting from the cell cycle, and initiating Igκ recombination and light chain editing [29]. Receptor editing in the light chain loci forms a critical basis for high-affinity maturation that is a continuous process in an ongoing immune response [30, 31], and impairment in κ chain rearrangement could account for the functional defects in T-dependent and independent antibody responses. Therefore, we next analyzed the light chain rearrangement. The orderly recombination of genes at the κ locus is the first step in the light chain rearrangement. The successful recombination of genes at the κ locus is proceeded by rearranging non-coding gene segments defined as ‘recombining sequence’ (RS) [32]. RS rearrangements occur higher in the most highly edited B cells. Thus, quantifying RS provides a readout for successful κ chain rearrangement [33]. Our results show that the levels of amplified RS products were moderately lower in the B cells derived from Iqgap1−/− mice (Fig. 6C). Thus, the functional defects observed in the B cells from Iqgap1−/− mice are due to an inability to rearrange the light chains appropriately.
B cell activation requires IQGAP1-Mek1/2-Erk1/2 signalosomeIQGAP1 is an essential scaffold in organizing Erk1/2 activation and phosphorylation [21]. Four tandem Isoleucine-Glutamine (IQ) domains are located between 745 and 864 amino acids. These IQ domains primarily mediate interactions with Rap1 [34], B-Raf or C-Raf [2], and Mek1/2 [21]. Therefore, we hypothesized that IQGAP1 regulates the sequential activation of the Raf/Mek1/2/Erk1/2 cascade. To test this, we activated purified total splenic B cells with soluble anti-IgM mAb and recombinant IL-4. Rac1/Cdc42 activates p21-activated kinase (Pak) that links the upstream BCR signaling to Raf by phosphorylating S445 in B-Raf and S338 in C-Raf [35]. By western blot analyses, we found increasing amounts of phosphorylated B-Raf (Serine 445) and phosphorylated C-Raf (Serine 338) in both Iqgap1−/− and WT B cells following anti-IgM-mediated activation (Fig. 7A). However, the level of B-Raf phosphorylation was much higher in the B cells from Iqgap1−/− but not the WT. Total B-Raf and total C-Raf were comparable between the Iqgap1−/− and WT B cells (Fig. 7A).
Fig. 7
IQGAP1 orchestrates ERK1/2 activation in B cells. A Sorted splenic B cells were activated with soluble anti-IgM antibody and recombinant IL-4 for indicated time points and lysed. Protein quantities were estimated and analyzed by western blots. Lysates were analyzed for the total and phosphorylated forms of Raf1, B-Raf, Mek1, Erk1/2, JNK1/2, and p38. Numbers below each blot represent fold change calculated following normalized values against their respective total and based on the phosphorylated protein levels of the WT. B PCR array results of splenic B cells following activation with anti-IgM antibody and recombinant IL-4. Data shown is one representative panel of three independent experiments. Data shown are obtained by repeated probing of the same membrane for indicated proteins
B-Raf or C-Raf catalyzes the phosphorylation of Mek1/2, which in turn phosphorylates Erk1/2 [36]. Since IQ domains recruit Mek1/2, we analyzed their activation status. and found that the phosphorylation of Mek1/2 was considerably reduced in B cells lacking IQGAP1. The WW domain of IQGAP1 (685–710 amino acids) contains two conserved tryptophan, positioned about 20–22 amino acids apart (Wx20–22W), that function as an interaction module for proline-rich ligands [37]. This domain is also responsible for recruiting Erk1/2 via a polyproline (APPPxxPY) motif [20]. Therefore, we next analyzed the phosphorylation status of Erk1/2 along with the two other MAPKs, Jnk1/2 and p38. All three kinases were active in the WT B cells. However, Erk1/2 and Jnk1/2 phosphorylation levels in Iqgap1−/− B cells were considerably reduced (Fig. 7A). p38 phosphorylation was not affected in the Iqgap1−/− B cells and was comparable to the WT.
We performed a PCR array analysis to define further the status of specific transcription factors altered in the early B cells from Iqgap1−/− mice. We stimulated the sorted splenic B cells with anti-IgM mAb and compared the expression of transcription factors between unstimulated and stimulated conditions from both strains. Averages from triplicates were used to calculate the fold change (Fc) between Iqgap1−/− B cells over WT under both experimental conditions (Fig. 7B). Similar to the total B cells from the BM, the levels of Foxo3, Foxo4, MafK, Stat5a, and Tcf4 have increased two or more folds in non-stimulated Iqgap1−/− B cells compared to WT. The expression levels of all these factors were considerably reduced in Iqgap1−/− B cells following anti-IgM mAb-mediated activation. However, the levels of c-Fos, Jun, Sox4, and Tbx21 were reduced under unstimulated and stimulated conditions. Interestingly, levels of Id2, c-Kit, Myc, Nfatc1, and Rel were increased in Iqgap1−/− B cells following anti-IgM mAb-mediated activation (Fig. 7B).
IQGAP1 is essential for suppressing Stat5a and Stat5bEarlier studies have shown that pre-BCR-mediated activation counters the IL-7R pathway [29]. Signaling via IL-7R sustains the survival and proliferation of pro-B cells via the activation of cell-cycle-related genes, including Ccnd3 [38]. In addition, phosphorylated Stat5A and Stat5B, downstream of IL-7R, translocated into the nucleus and formed a complex with PRC2/EZH2 to bind to 5′ intronic enhancer of the κ gene (iEκ) to actively suppress its transcription by limiting the access to E2A, which is induced by the pre-BCR signaling. In addition, the pre-BCR-mediated Ras-Mek1/2-Erk1/2 pathway also silences Ccnd3 to orchestrate the exit from the cell cycle and initiate κ chain recombination [29, 39]. Next, we investigated the status of E2A, Ezh2, Stat3, Stat5A, and Stat5B in WT and Iqgap1−/− B cells. First, we analyzed the protein levels of these transcription factors in the total B cells derived from the BM of both strains by western blot analyses (Fig. 8A). The limited numbers of pro-and pre-B cells excluded our ability to use them in these assays. E2A, Stat5A, Stat5B, and Ezh2 were comparable between the WT, Iqgap1−/− B cells.
Fig. 8
IQGAP1 is required to suppress Stat5a and Stat5b and to initiate the transcription of IRF4 and IRF8. A Total sorted BM B cells were used to analyze E2A, Stat3, Stat5A, Stat5B, IRF4, and β-actin protein levels by western blot analyses. Data shown are representative of three experiments. B Total sorted BM B cells were activated with anti-IgM mAb, and the cytoplasmic and nuclear protein fractions were separated and analyzed for E2A, Stat3, Stat5A, Stat5B, and β-actin. Data shown are representative of three experiments. C mRNA from sorted BM B cells was used to quantify the expression of Irf4 and Irf8 transcripts. Data shown are the average relative expression of these transcripts from four mice. D mRNA from sorted splenic B cells was used to quantify the expression of Irf4 and Irf8 transcripts. Data shown are the average relative expression of these transcripts from four mice. Statistical significance was calculated using the student’s t-test, and p-values are shown below in each graph. E Cartoon summarizing the role of IQGAP1 during early B cell development. IL-7R transduces its signal via Stat3, Stat5A, and Stat5B. Signals via pre-BCR initiate two distinct pathways, One via SLP65-mediated IRF4 and IRF8 activation and the second via the Ras-RafB/C-Mek1/2-Erk1/2/Jnk1/2 pathway mediated by IQGAP1. Four tandemly organized IQ domains can recruit C-Raf or B-Raf. IQ domains also have the specificity to Mek1/2. Rafs mediate the activation of Mek1/2 on the IQGAP1 scaffold. The WW domain of IQGAP1 recruits Erk1/2, which Mek1/2 phosphorylates. Phosphorylated Erk1/2 activates several transcription factors, including E2A, Elk1, and CREB. E2A competes with Stat5 dimers for the 5′ intronic Eκ enhancer accessibility. Successful displacement of Stat5 dimers initiates the recombination process of Igκ gene. E2A also relieves the Stat5-mediated repression of Rag1 and Rag2 genes. Stat5-mediated transcription of Ccnd3 is relieved by the phosphorylated Ras-Mek1/2-Erk1/2 pathway. The potential candidates that repress the transcription of Ccnd3 are Cdkn2a or p21. The mechanism by which IQGAP1 regulates the expression of Irf4 and Irf8 genes is not known. However, the role of IRF4 in B cell development has been well-established. IRF4 binds to the 3′ Eκ, further promoting light chain recombination. IRF4 also binds to the 5′ Eλ to initiate the transcription of the Igλ gene. IRF4 and Ikaros3 (Ailos) repress the surrogate light chain component λ5-encoding gene. IRF4 also plays an important role in initiating the transcription of Cxcr4. It is essential for the pre-B cells to traffic from their IL-7-producing stromal niche to the IL-7-deficient niche to transition into immature B cells. Repressing Id2 and Id3 is also needed for the pre-B cells that are regulated by Ebf1, a potential target downstream of Erk1/2
In contrast, no protein band was found for IRF4 in Iqgap1−/− B cells (Fig. 8A). Next, we evaluated the levels of these transcription factors translocated into the n
留言 (0)