The significance of lymphangiogenesis in various diseases has been extensively examined in recent literature. These studies have provided significant insights into the contrasting effects of lymphangiogenesis on disease pathophysiology. Lymphangiogenesis relies on the proliferation, migration, and differentiation of lymphatic endothelial cells (LECs). The cellular processes lead to the biosynthesis of lymphatic vessels, which transport excess fluid and regulate immune responses in the lymphatic system [1]. Recent studies utilizing genetic lineage tracing and single-cell RNA sequencing have demonstrated that stem/progenitor cells also play a crucial role in lymphangiogenesis [2]. Additionally, M1 macrophages have been shown to polarize and transdifferentiate into new LECs through activation of the vascular endothelial growth factor (VEGF-C)/vascular endothelial growth factor receptor 3 (VEGFR3) pathway [3]. Lymphatic vessel proliferation comprises healthy lymphangiogenesis (during wound healing and corpus luteum development) and pathological lymphangiogenesis. The latter is caused by pathological situations such as inflammation, tumors, and transplant rejection, among others [4,5,6]. Physiological and pathological lymphangiogenesis often entail the enlargement and sprouting of preexisting lymphatic vessels (LVs) rather than neolymphangiogenesis, which is more closely related to lymph node transfer [7]. The interaction between lymphangiogenesis and various clinical conditions has a complex effect on the organism. Advanced imaging and genetic approaches have made it possible to investigate specific structures and functions within the lymphatic systems in various diseases.
Lymphangiogenesis plays complex immune regulatory roles via various mechanisms, differing from the nuanced variations of microenvironments in tissues and organs. The newly formed lymphatic vessels can either enhance or inhibit the immune response [8, 9]. The lymphatic system maintains homeostasis and supports immune responses throughout various tissues and organs [10]. In both health and disease, the lymphatic system also plays a crucial role in regulating immune responses by directly influencing immune cells and coordinating their movement from tissues to draining lymph nodes (dLNs) [11]. The unique characteristics of lymphatic vessels in both health and disease demonstrate specificity related to tissue and organ types. The characteristics influence the varied outcomes of lymphangiogenesis in different disease contexts.
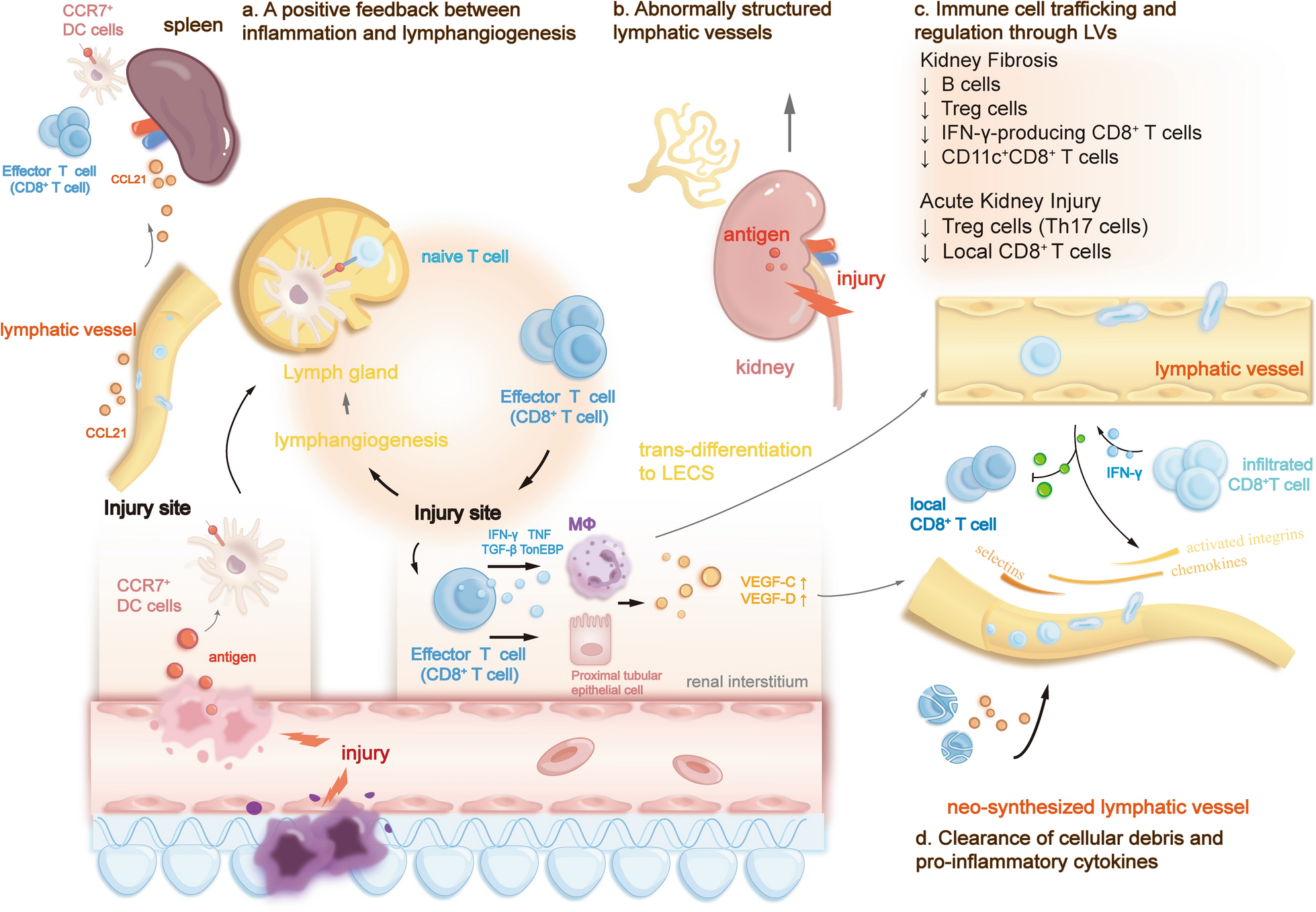
Within the kidney, lymphangiogenesis is closely linked to kidney tissue inflammation, fibrosis progression, and transplant rejection [12]. Evidence unveils that it can elicit dual-sided effects in various kidney pathologies [12, 13]. Studies have illuminated that kidney lymphangiogenesis exhibits an intricate immune regulatory mechanism capable of promoting or alleviating immune responses [14], depending on the specific kidney pathology under consideration. Emerging evidence suggests that, within kidney diseases, the distinct trafficking patterns of diverse immune cells and varying durations of different pathological conditions significantly contribute to the dual-sided effect of lymphangiogenesis [15, 16].
The kidney lymphatic system selectively transports renal interstitial fluid and immune cells. It actively contributes to the maintenance of kidney homeostasis and the orchestration of kidney immune response. Notably, preexisting lymphatic vessels within the kidney are predominantly distributed in the renal cortex and rare in the medulla. However, neo-synthesized lymphatic vessels can proliferate extensively throughout the kidney [17].
Lymphatic migrations of immune cells are regulated by Various chemokines, including chemokine (C–C motif) ligand 19 (CCL19), CCL21, and chemokine (C-X-C motif) ligand 12 (CXCL12). Additionally, several inflammatory and anti-inflammatory mediators, including Interleukin-1β (IL-1β), Tumor Necrosis Factor-α (TNF-α), Interleukin-10 (IL-10), and Transforming Growth Factor β (TGF-β), also involves in the regulation of lymphatic immune cell migrations [1].
To be specific, recent findings have uncovered that lymphatic vessels can suppress the expansion of CD8+ T cells [18, 19]. The interaction between Mac-1 on DCs and ICAM-1 on LECs mediates the adhesive interactions between DCs and LECs, thereby inhibiting the ability of DCs to induce T cell proliferation [20]. Moreover, chemokine receptor chemokine (C–C motif) receptor 7 (CCR7) expressed on DCs and its CCL21 produced by LEC are the main molecules involved in DC migration [21]. At the same time, reducing DCs is beneficial for slowing the progression of inflammation [22]. The reasons and mechanisms underlying this remarkable discrepancy in diverse kidney diseases require further investigation.
LECs and their immune regulatory role
LECs are crucial in immune responses during inflammation, tumour, and other pathological conditions. Different subsets of LECs, including peripheral capillary LECs and lymph node LECs, have distinct functions. The primary functions of peripheral capillary LECs include fluid drainage, leucocyte transport, and participation in lipid metabolism. They also actively regulate the endocytosis of antigens, mediating by clathrin and caveolin. Interestingly, capillary LECs exhibit phenotypic adaptations in varying microenvironments [23]. Therefore, they can dynamically orchestrate the trafficking and activities of various immune cells. Among the intricate process of immune cell trafficking, lymphatic vessel endothelial receptor-1 (LYVE-1) makes the first adhesive contact between migrating immune cells and lymphatic endothelium, initiating the entry and trafficking of immune cells within afferent lymphatic vessels [24]. Moreover, capillary LECs secrete various chemokines to drive immune cell intravasation through a complicated process of actomyosin-mediated immune motility and β2 integrin activation during inflammatory status [24]. Among these cytokines, CCL21 is one of the most important and well-studied regulators. By binding to heparan sulfate within the extracellular matrix, CCL21 generates a hypotactic concentration gradient to promote the migration of diverse leucocytes, such as DCs, neutrophils, and monocytes, through interacting with CCR7 expressed on these immune cells [25,26,27]. Furthermore, accumulating data suggests numerous cytokines and chemokine/receptor combinations are involved in lymphatic migration. Immunosuppressive substances like IL-10 and TGF-β may prevent immune cells from migrating through the lymphatic system [28,29,30]. LN LECs exert varied functions after transporting molecules and cells to dLNs. These cells are pivotal contributors to immune surveillance in both health and disease. LN LECs and specifically distributed rapidly classify molecules [31,32,33,34,35]. These two types of LECs subtly regulate innate and adaptive immune responses [21, 22, 36,37,38,39,40,41].
Cytokines and chemokines involved in lymphangiogenesis
Lymphangiogenesis is predominantly regulated by VEGF-C and VEGF-D, both of which directly bind with VEGFR-3 and the co-receptor neuropilin 2 (NRP2), expressed on the surface of LECs, subsequently inducing lymphangiogenesis [42]. Recent studies have revealed that several types of macrophages can promote lymphangiogenesis by secreting VEGF-C in various pathological conditions, including kidney damage and cardiac injury [43,44,45,46,47]. Cortical and medullary kidney tubules can secrete VEGF-C and VEGF-D within the kidney [48]. Studies suggest that VEGF-C has an essential role in the development of lymphangiogenesis, but its impact on the maintenance of lymphatic vessels might be limited [49]. Conversely, unlike VEGF-C, VEGF-D dominates the maintenance of lymphangiogenesis, which indicates a modulatory function of VEGF-D in its developmental stage [49]. Furthermore, TGF-β and connective tissue growth factor (CTGF) also contribute to the induction of lymphangiogenesis in kidney diseases, particularly in kidney inflammation and fibrosis [50]. Additionally, angiopoietins (Angs) are involved in the lymphangiogenesis mechanism. In fact, the Ang2/Tie/PI3K signaling pathway plays a crucial role in lymphangiogenesis; blocking this pathway leads to a decrease in VEGFR3 and inhibits lymphatic vessel formation [51]. Similarly, the transcription factors FOXC1 and FOXC2, which are part of the Forkhead box (FOX) family, positively regulate lymphangiogenesis. Studies have shown that FOXC1 and FOXC2 are essential for regulating the Ras/ERK signaling pathway during lymphangiogenesis, and the loss of FOXC1 and FOXC2 promotes excessive activation of ERK, leading to abnormal lymphangiogenesis [52].
The immune regulatory functions of lymphangiogenesis in kidney diseasesAcute kidney injury
In many AKI animal models and AKI patient biopsies, VEGF-C and VEGF-D expression increased, as did robust lymphangiogenesis. Following kidney damage, inflammatory mediators such as Interferon-gamma (IFN-γ), TNF-α, and TGF-β promote lymphangiogenesis via several mechanisms [48, 53,54,55,56]. Functional neo-lymphatic vessels can manifest the same role as preexisting renal lymphatic vessels, promoting inflammation resolution through drainage of retained fluid, clearance of cellular debris, removal of pro-inflammatory cytokines and cells, and mobilizing immune cells [11, 48]. Macrophages are highly adaptable to transfer into various distinct phenotypes within the local microenvironment. Among these macrophages, M1 macrophages are predominantly infiltrated during the AKI process, primarily promoting inflammation response and inducing kidney injury [57]. They enhance the synthesis of new lymphatic vessels in renal inflammation and fibrosis microenvironment due to elevated expression levels of VEGF-C induced by TGF-β [49]. Recent evidence has demonstrated that M1 macrophages can directly contribute to the synthesis of new lymphatic vessels through transdifferentiating into LECs [3, 58]. Increased levels of VEGF-C directly suppress macrophage autophagy, which prompts M1 macrophage polarization into LECs [3].
Lymphangiogenesis adversely affects the AKI process, exacerbating renal inflammation [56, 59, 60]. In general, AKI-induced lymphangiogenesis can exert a dual-sided impact on the kidney. In AKI, kidney lymphangiogenesis also acts as an immunological regulator to balance immunity and immune pathology despite clearing excessive fluids, noxious stimuli, and inflammatory cells. Neo-synthesized lymphatic vessels can either induce or suppress the immune response in AKI models, regulating local and systematic immune systems through diverse mechanisms [61]. Firstly, it significantly enhances lymphatic flow, actively participating in the early immune regulation process after AKI. During AKI’s initiation, maintenance, and regression process, abundant immune cells, including inflammatory monocyte, neutrophil, lymphocyte and natural killer cells, orchestrate the overall immune response [62]. Therefore, lymphangiogenesis can mitigate kidney tissue inflammation by properly removing infiltrated immune cells at the injury site (Tables 1, 2).
Table 1 The role of important immune cells that involve in kidney diseaseTable 2 Two-sided consequences of immune regulation through lymphangiogenesis in kidney diseasesDespite lymphatic flow, some immune cells, including T cells, B cells, and DCs, can also directly regulate LECs-related signal pathways [63, 64], contributing to the inflammatory progress. Entry of naïve T cells to afferent lymphatics is regulated by the S1P (sphingosine-1-phosphate) receptor pathway [65], while memory T cells also possess CCR7, which binds with CCL21. Immune cells with CCR7 can also be regulated by the CCL21 gradient expressed by LECs, actively migrating to dLNs through afferent lymphatics [66]. Of note, current evidence has demonstrated that enhancing antigen-specific T helper cell 1 (Th1 cell) cell migration from tissues to dLNs accelerates the resolution of inflammation. In the setting of AKI, upgraded infiltration of Th cells (T helper cells), particularly T helper cell 17 (Th17 cell), has been observed [67]. Th17 cell, which aggravates tissue injury by recruiting neutrophils and other inflammatory cells, is the most abundant lymphocyte infiltrated at the injury site following AKI in mice [68, 69]. Additionally, intestinal flora-derived Th17 cells have been proved to migrate to the kidney in kidney disease. They enter peripheral blood circulation through lymphatic vessels regulated by the S1P-R1 pathway [16]. Subsequently, they return to the renal inflammation site through blood circulation, further exacerbating the inflammatory response [16]. Increased reduction of Th17 cells through kidney lymphangiogenesis may significantly mitigate kidney damage, alleviating AKI and the following progression to chronic kidney disease (CKD). However, further studies are required to demonstrate whether lymphangiogenesis can aggravate kidney damage by regulating these Th cells.
Moreover, due to the adaptiveness of LECs, lymphangiogenesis can also directly suppress the local CD8+ T cells during inflammation. This intricate mechanism has been well-studied in the setting of skin lymphangiogenesis. Lymphatic endothelial cells (LECs) largely express nonhematopoietic programmed death-ligand 1 (PD-L1) to limit local CD8+ T cell effectors to functioning in inflamed skin and melanoma [70]. Despite PD-L1-dependent inhibition of T cell antigen receptor (TCR) signaling, evidence supports that PD-L1 can regulate lymphocyte migration through endothelial and epithelial barrier tissues [70], which indicates that PD-L1 may directly regulate T cell transendothelial migration without antigen presentation mechanism. Similarly, the activated PD-1 signalling pathway in the kidney protects the ischemia–reperfusion-induced AKI mouse model [71]. At the beginning of inflammation, infiltrated antigen-specific CD8+ T cells produce IFN-γ, which directly induces PD-L1 expression in adjacent inflammation-induced lymphatic vessels. Abundant PD-L1 expressed by LECs limits the further accumulation of CD8+ T cells at the injury site, alleviating kidney inflammation.
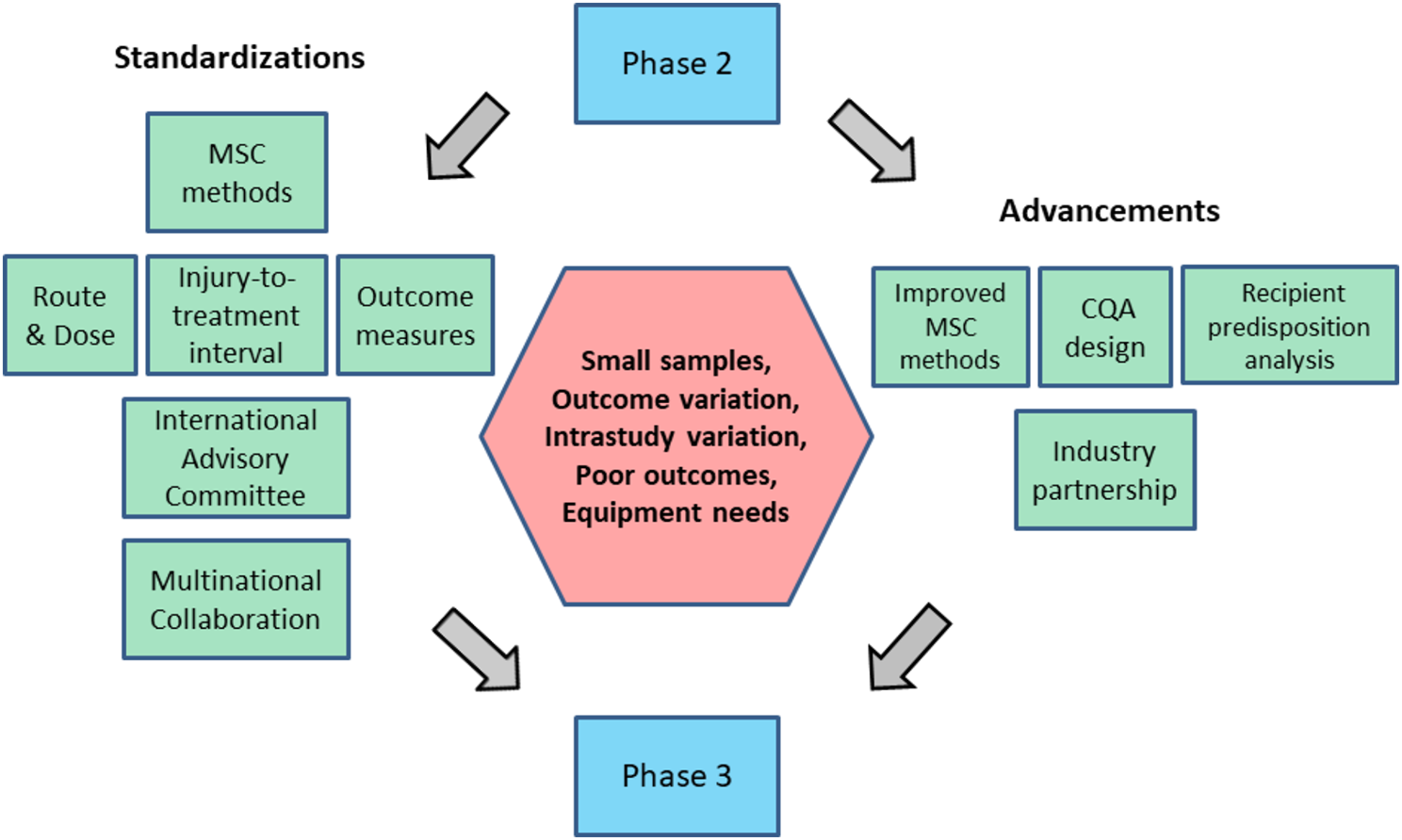
In the context of AKI, lymphatic migration of immune cells affects local immunity bidirectionally. Current evidence has demonstrated that lymphangiogenesis can also be detrimental to AKI [11, 72]. This detrimental impact closely correlates with positive immune feedback (Fig. 1) that enhances immune cells’ constant migration and activation at the injury site. During kidney inflammation, the increased level of CCL21 that is overexpressed by preexisting LECs, along with other chemokine and integrin pathways, promotes kidney dLNs and spleen to recruit more CCR7+ immune cells through afferent lymphatic vessels [73, 74]. The significantly elevated recruitment leads to systemic expansion of lymphocytes [56]. Within kidney dLNs, CCR7+ DCs present antigens of injury sites to CD8+ T cells, promoting T cell proliferation in dLNs. After which, the activated CD8+ T cells retu








留言 (0)