
記住我

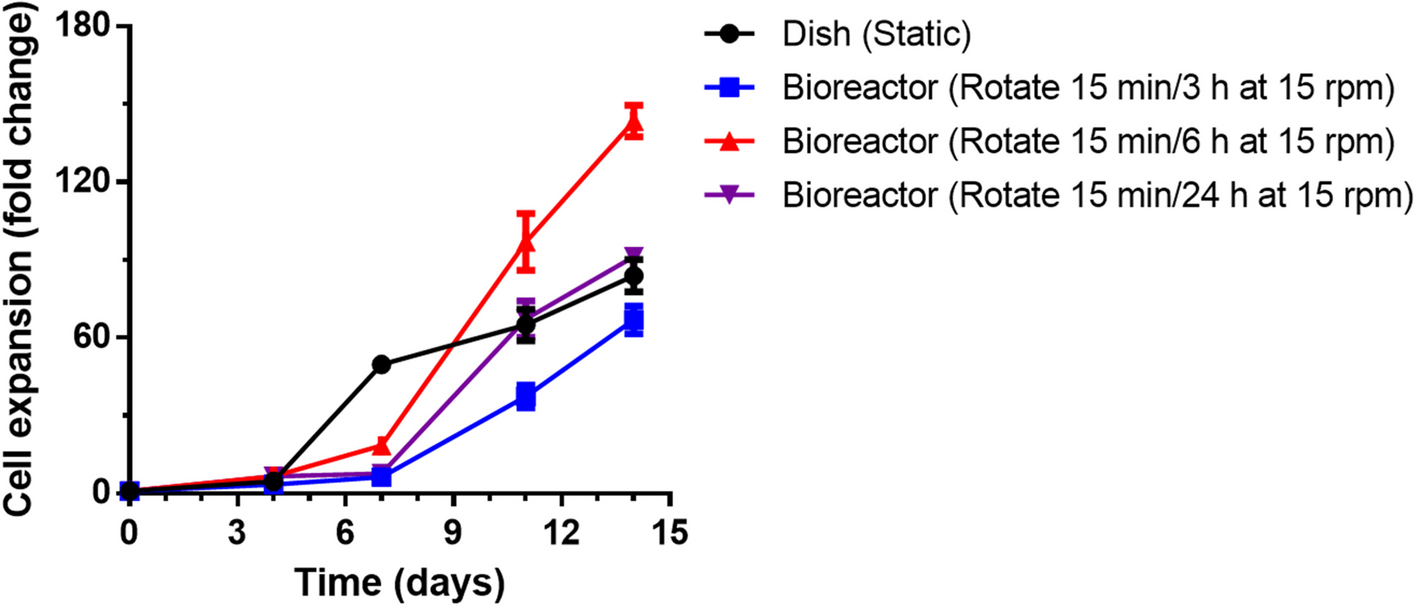





To assess the impact of different rotation parameters on overall cell proliferation, various rotation cycles were initially measured (Fig. 1). The cells cultured via traditional methods in static dishes exhibited rapid growth during the initial 7 d. The cells then stabilized and exhibited slow growth, reaching an approximately 84-fold increase in cell number by day 14. This deceleration resulted from the formation of larger cell aggregates, which, in turn, reduced the cell proliferation rate in the later stages of static culture. In contrast, the pseudostatic culture showed relatively slower growth in the first 7 d but grew faster in the subsequent period. This is due to an adaptation phase in the pseudostatic bioreactor during the initial 7 d, during which period rotation assists in optimizing cell aggregation and dispersion. The optimal cell aggregate size significantly affects cell‒cell interactions, increasing the availability of crucial soluble factors, such as growth factors, cytokines, nutrients, and bioactive molecules, which are important for mediating cellular proliferation [23]. In the pseudostatic culture system, an inadequate rotation frequency (15 min/24 h) resulted in suboptimal dispersion, whereas an excessively high frequency (15 min/3 h) affected cell aggregation, consequently impeding proliferation. At an appropriate rotation frequency (15 min/6 h), the cells showed a remarkable 144-fold increase in proliferation by day 14, significantly increasing their potential for cell expansion compared with that of conventional static dish cultures.
Fig. 1
The proliferation of NK-92MI cells cultured in a static dish control system and a pseudostatic bioreactor system on days 4, 7, 11, and 14
Shear stress has also been identified as a critical factor in maintaining cell cultures in bioreactors [24]. The gentle agitation generated by the pseudostatic system's rotation not only promotes uniform distribution of nutrients and oxygen but also applies controlled shear stress to the cells. This helps regulate aggregate size and structure, preventing the formation of overly large clusters that could impede mass transfer. By fine-tuning the rotation parameters, the bioreactor achieves a balance between promoting cell growth and avoiding excessive stress, which could otherwise lead to cell damage or death.
Viability of NK-92MI cellsThe viability of NK-92MI cells under various culture conditions was assessed by flow cytometry. Although the total cell count was greater after 7 d of static culture (Fig. 1), the increase in cell aggregation and local accumulation of waste slightly reduced cell viability compared with that of the other pseudostatic groups, resulting in approximately 65% viability after 14 d (Fig. 2). In the bioreactor group employing periodic rotation, where the size of the cell aggregates affects the proliferation efficiency, the cell viability consistently surpassed 85% after 14 d. These findings underscore the need for suitable dynamic culture conditions to maintain NK-92MI cell survival. To optimize cell expansion and viability, the parameters were standardized to a rotation of 15 min/6 h at 15 rpm to ensure the most effective cell amplification ratio and sustained cell viability.
Fig. 2
The viability of NK-92MI cells cultured in a static dish control system and a pseudostatic bioreactor system was evaluated via flow cytometry on days 4, 7, 11, and 14
Live/dead cell staining and NK-92MI cell aggregate sizeAfter 1, 7, and 14 d of cell culture in both static dishes and bioreactors, meticulously isolated cell aggregates were observed via confocal microscopy to perform live/dead cell staining (Fig. 3). The average aggregate sizes are summarized in Table 1. In static culture, the sizes of the cell aggregates on days 7 and 14 were 140.71 ± 48.97 µm and 395.70 ± 86.15 µm, respectively. Conversely, the periodic shear stress of the bioreactor’s pseudostatic culture notably decreased the cell aggregate size, which averaged approximately 118.44 ± 32.13 µm by day 14. Such variations in aggregate size directly affect survival rates. Although NK-92MI cells cultured in traditional static culture initially grew rapidly (Fig. 1), after 14 days, the inner part of the larger cell mass underwent apoptosis owing to limited mass transfer (Fig. 3). In contrast, the bioreactor group with an appropriate cell aggregate size exhibited fewer instances of apoptosis after 14 days, as depicted in Fig. 2.
Fig. 3
Evaluation of NK-92MI cells via a live/dead cell staining assay after 1–7 and 14 days of cultivation. Living cells in green were stained with calcein AM, and dead cells in red were stained with propidium iodide. Scale bar = 100 μm
Table 1 The Average ± standard deviation of cell aggregate size in static dish and pseudo-static bioreactor systemsWhen the cell aggregate size exceeds 300 µm, nutrient and oxygen transport to and waste transport from the center of the aggregates become suboptimal, leading to apoptosis in the central cells of the aggregates [23,24,25,26]. This finding was corroborated by live/dead cell staining after analysis via flow cytometry (Fig. 4). While a certain level of cell death is expected in culture, the data clearly show that the pseudostatic culture method significantly reduces the proportion of dead cells compared to static culture conditions. And Fig. 5 illustrates the general size distribution, indicating that the bioreactor system effectively maintained size control.
These results suggest that the homemade bioreactor system developed in this study effectively generated cell aggregates of appropriate size, promoted cell proliferation and mass transfer, and ultimately improved cell survival rates.
Fig. 4
Flow cytometry analysis of the live/dead stained NK-92 MI cells after 14 d of cultivation. a Cells were cultured via a traditional static culture method in a culture dish. b Cells were cultured via a pseudostatic culture method in a custom pseudostatic bioreactor
Fig. 5
Observation of spheroid morphology and size distribution under static and pseudostatic culture methods after 14 days of cultivation. Bright field images in (a) show the morphological characteristics of cell spheroids in static dish cultures, while (c) illustrates those in a bioreactor system. The size distributions for the static and pseudostatic culture methods are presented in (b) and (d), respectively. Scale bar = 100 μm
Phenotypic changes in activated NK-92MI cells in the self-designed bioreactorAfter optimizing the pseudostatic culture of NK-92MI cells, they were further introduced into a self-designed, one-step, homemade bioreactor system, allowing the cells to proliferate within a closed system and undergo IL-15- or IL-18-induced activation. To explore potential alterations in the NK-92MI cell phenotype induced by this culture system, qPCR analysis was performed to measure several NK-92MI cell markers (Fig. 6). The mRNA levels of the positive markers CD2 and CD56 were detected in NK-92MI cells, whereas the native negative marker CD16 was not detected [15, 27]. These findings suggested that neither the bioreactor culture system nor activation with IL-15 or IL-18 significantly altered the specific phenotype of NK-92MI cells.
Fig. 6
Positive NK-92MI markers, including CD2 and CD56, and the negative NK-92MI marker CD16 were used to identify the NK-92MI phenotype. The mRNA levels are expressed as relative levels normalized to the GAPDH level (defined as 1)
Cytotoxicity-related gene expression in NK-92MI cells after homemade bioreactor cultivationWe further investigated the genes associated with immunotherapy and cancer cell killing in NK-92MI cells and compared their expression levels with those of cells cultured conventionally in cell dishes (Fig. 7). NKG2A is an inhibitory NK receptor that recognizes specific ligands on target cells and transmits inhibitory signals [28]. Moreover, NK cells possess the dual capacity to produce proinflammatory (IFN-γ) and anti-inflammatory (IL-10) cytokines, highlighting their essential role both as inflammatory agents for clearing infections and as regulators to manage inflammation and mitigate immune-mediated damage to the host [29, 30]. Higher expression levels of IFN-γ in NK-92MI cells correlate with a stronger ability to kill cancer cells, while IL-10 acts as an antagonist of NK cell activation [31, 32]. Compared with those of normal NK-92MI cells, upon entry into the reactor, the expression levels of genes associated with the inhibition of NK-92MI cell activation, such as NKG2A and IL-10, were slightly decreased. Conversely, the expression levels of genes related to activated NK-92MI cells, particularly IFN-γ, were significantly increased, especially in the IL-15 and IL-18 activation groups. Compared with those in normal NK-92MI cells, the gene expression levels of IFN-γ in the IL-15 and IL-18 activation groups were upregulated 2.8- and 3.2-fold, respectively. These findings are consistent with those of previous studies that utilized IL-15 or IL-18 for NK cell activation [32, 33].
Fig. 7
Cytotoxicity-related genes, including NKG2A, IFN-γ, and IL-10, were used to evaluate the increase in gene expression in activated NK-92MI cells. Relative gene expression levels were normalized to GAPDH gene expression levels and expressed as the fold change compared with the levels in normal NK-92MI cells
NK-92MI cell-mediated cellular cytotoxicityTo determine the activity of NK-92MI cells against cancer cells via the developed homemade one-step proliferation and activation closed bioreactor culture system, the human erythroleukemic cell line K562 was used as the target cell line (Fig. 8).
Fig. 8
NK-92MI cell-mediated cytotoxicity against K562 cells. After the proliferation and activation of NK-92MI cells induced by IL-18 through a homemade bioreactor, the cells showed a cytotoxic effect on K562 cells after coculture for 0 h (a), 2 h (b), 3 h (c), and 4 h (d). e Quantification of NK-92MI cell-mediated cytotoxicity was determined by measuring the fluorescence intensity in the supernatant via a plate reader
After labeling with calcein AM, K562 cells exhibited green fluorescence under a fluorescence microscope. After 14 days of proliferation and activation, the NK-92MI cells were cultured. As shown in Fig. 8b–d, within 4 h, the amount of fluorescent dye released from the killed K562 cells gradually decreased, reducing the fluorescence intensity of the cells. The real-time behavior of NK-92MI cells in response to the target K562 cancer cells is shown in Supplementary Video S1. These outcomes enabled further collection of supernatants and the quantitative assessment of cytotoxicity via a specific lysis formula [2, 34]. The results confirmed that the activated cells exhibited enhanced cytotoxicity against K562 cells (Fig. 8e). Specifically, the IL-18-activated group exhibited a 1.62-fold increase in cytotoxicity compared with the nonactivated group. Previous studies have indicated that the cytotoxicity of NK-92MI cells to cancer cells can be increased through IL-18-induced activation [33, 35]. However, IL-15-induced NK-92MI cell-mediated cytotoxicity was not notably pronounced, with a 1.17-fold increase (Fig. 8). It has been reported that IL-15 has some influence on cell survival and can enhance cellular cytotoxicity within a short 24 h period, but this effect diminishes after 96 h [35]. In this study, the relatively modest impact of IL-15 activation on cytotoxicity may be attributed to its suitability for the short-term activation of NK-92 cells.
It has been reported that cytokines captured by NK cells can indirectly trigger signal transduction in surrounding NK cells through cell‒cell interactions within the cell aggregate, maximizing the utilization of cytokines [18, 36]. Therefore, maintenance of aggregate characteristics may be directly related to cell functionality. Additionally, providing adequate mechanical stimulation, including turbulence and stress to NK-92MI cells, would activate NK-92MI cells and generate functional extracellular vesicles of high quality and quantity for cancer treatment [22].
In this study, we designed a custom one-step proliferation and activation closed bioreactor culture system. The NK-92MI cells rapidly proliferated in the upper chamber, forming the adequate cell aggregate size necessary for the desired cell count and survival. Through simple valve control, the 3D NK-92MI cell aggregates were subsequently directed to the lower chamber for specific activation in a single step, avoiding cell loss and cultivation interference during the 14-d culture period. While their distinct cell surface phenotypes are maintained, activated cells enhance the expression of genes associated with cell cytotoxicity, thereby increasing NK-92MI cell-mediated cytotoxicity against tumor cells. This closed system ensures high efficiency, is eco-friendly, is easy to operate, and mitigates the overall experimental contamination risk. Moreover, this design has potential for broader applications in the proliferation and activation of other suspended immune cells, potentially opening new avenues for immunocellular therapy.
留言 (0)