
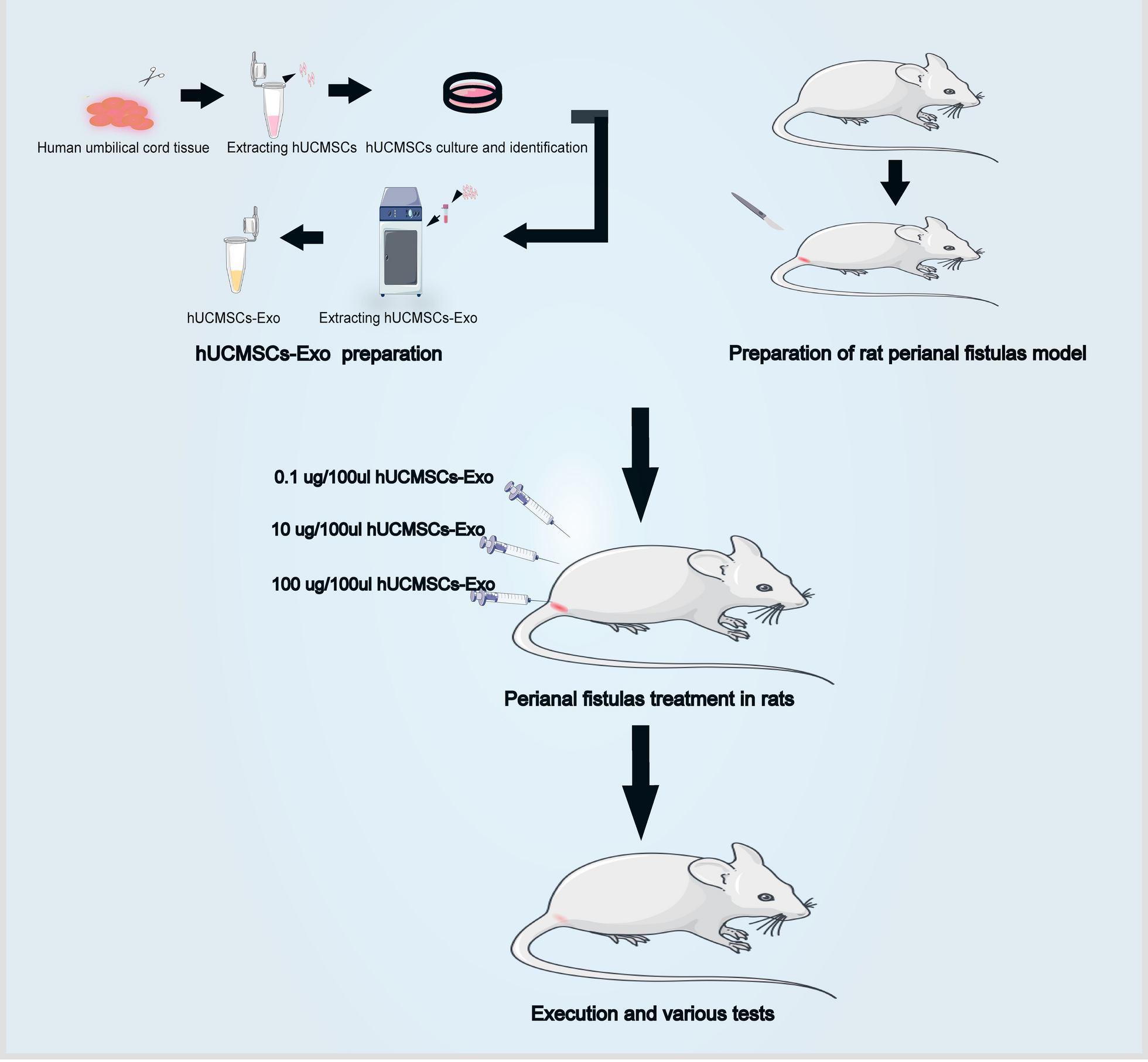
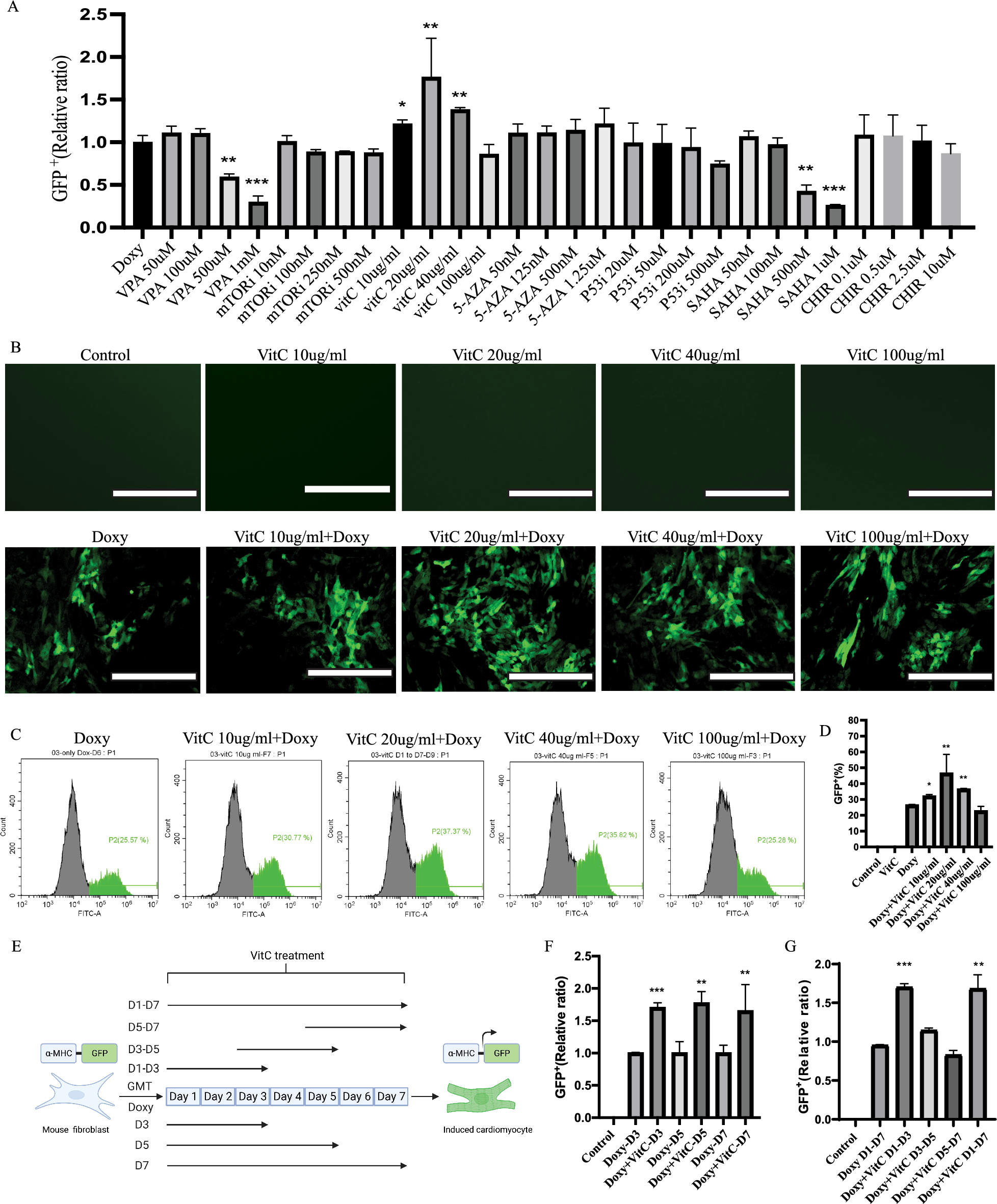
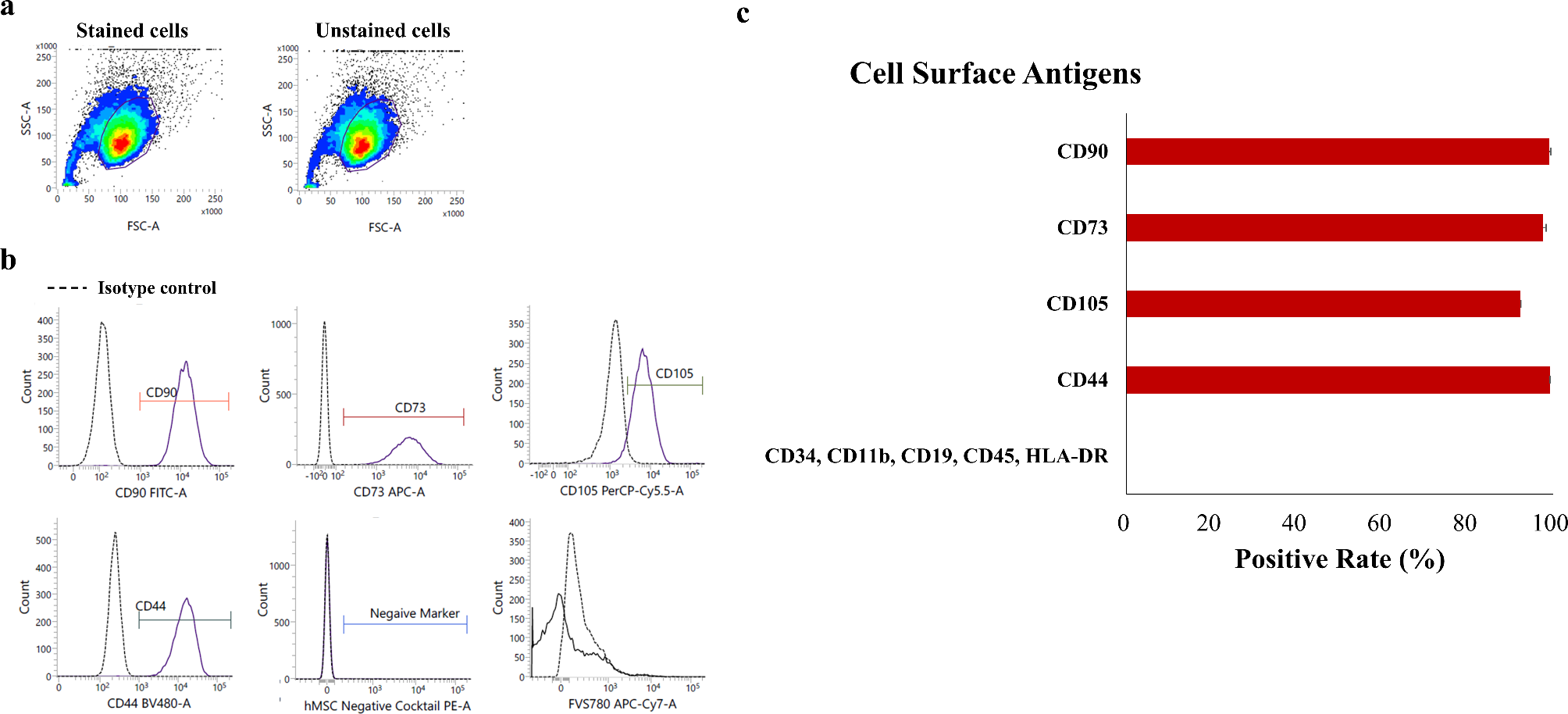
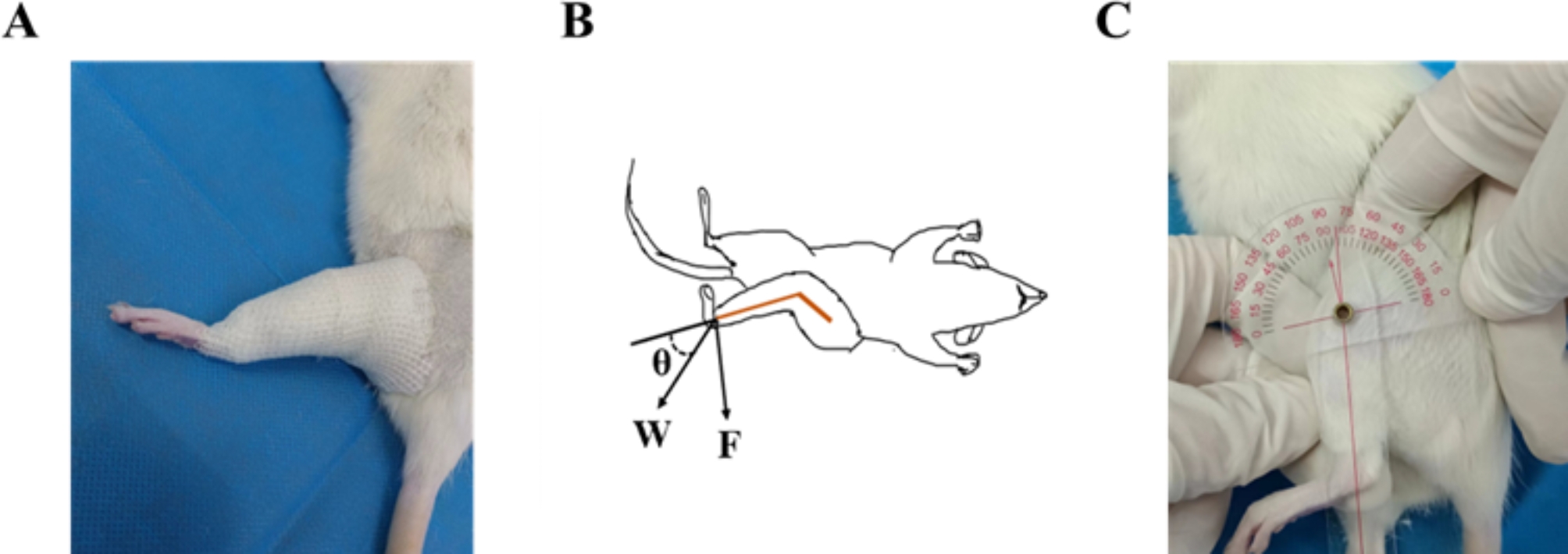
記住我






The protocol was originally developed using adherent cultures and derived from design of experiments [18]. Suspension protocol was initiated in TB10 media (Table 1) to support aggregate formation [36]. This media was designed to stabilize aggregate formation while limiting aggregate growth. CDM2 medium [37, 38] poorly sustained aggregate growth during iPSC differentiation in bioreactors (Data not shown), so a descendent media was developed (TB2) specifically for sustaining iPSC differentiation in bioreactor suspension conditions (Table 1). Initial evaluation of the protocol transfer was performed in two culture conditions: adherent and in suspension bioreactors using the NCRM1 iPSC-line. Stage 3 cells from a suspension environment and adherent culture were compared and evaluated through RNA sequencing (Figure S1). RNA sequencing data in heatmap (Figure S1-A) and volcano plot (Figure S1-B-C) evaluation of core endocrine genes showed that expression levels in adherent cells were lower than those expressed in 3D environment (Figure S1-A). Several genes that are essential for insulin transcription such as NEUROD1, MAFA, PPARGC1A and NKX2.2 [19] along with INS were expressed with higher transcript levels in bioreactor samples. It was observed that genes usually expressed in earlier stages such as FOXA2 and HNF1β had higher expression levels in adherent culture indicating that 2D environment differentiation might be lagging. Since the cell architecture of human islets is in 3D and not 2D, dissociating and reaggregating the cells can more closely mimic conditions during embryonic islet development [22]. Thus, endocrine stage adherent cells were dissociated and aggregated in flasks (Figure S1-D) as a means of additional comparison of the final material as aggregates. ELISA assay was used for quantifying C-peptide content per aggregate in this comparison. The results indicated that the C-peptide per aggregate content observed in suspension environment was significantly higher compared to pseudoiselets generated from the adherent cultures (Figure S1-E). These results were consistent with the data from RNA sequencing despite reaggregation. Thus, there are discrepancies between culture characteristics requiring custom differentiation optimization. Aggregate size and density might be potential factors impacting the differentiation process and cell fate [36].
Expansion and differentiation of iPSCs into insulin-producing cellsTo develop and solidify the protocol transferred from adherent to suspension environment, an iterative strategy of stage-wise optimization was used to determine and control critical process parameters using two cell lines. Prior to initiating the differentiation protocol, 90 million adherent cells were collected from flasks of two different cell lines, RCRP5005N and NCRM-1. Each batch was then reseeded into a 500 ml vertical wheel bioreactor at a density of approximately 1.8 × 10^5 cells/ml in TB10 media which consists of Essential 8 (E8) medium supplemented with polyethylene glycol (PEG) and Heparin Sodium Salt (HS). On the 3rd day, differentiation was initiated as previously described [18]. The protocol, originally optimized for adherent culture using a HD-DoE approach, aimed to induce dorsal foregut endoderm from pluripotent stem cells. This protocol (TB-beta) was modified as depicted in (Fig. 1A) with the basal media TB2 (Table 1). Modifications in the basal media supplements were made to enhance aggregate stability and limit aggregate fusion events, thus minimizing cluster diameter.
Fig. 1
Bioreactor-based differentiation protocol timeline. A Schematic of the 3-stage differentiation pancreatic protocol used in bioreactors. Figure is created with BioRender.com B Images of bioreactor aggregates sampled throughout the different protocol stages. C Average aggregate size measurements on two different cell lines used throughout the protocol
During the initial 3 days of differentiation toward definitive endoderm, cells were optimized for FOXA2 expression. The morphology and growth of the clusters were characterized at all stages of differentiation, and the aggregate diameter measured in the range of (300 ± 100) µm for both cell lines (Fig. 1B–C). Retinoic acid, LDN3189, A8301, and PD0325901 were identified as differentiation factors for stage 1 (DFE) based on previous optimization efforts for HNF1β and FOXA2 [18]. By the end of this stage, bioreactors contained approximately 325 million cells of NCRM-1 and 420 million cells of RCRP5005N, representing ~ 4 × fold expansion throughout the process. Following the DFE stage, aggregates from each bioreactor were split into two additional 500 ml bioreactors to control cellular density. One half of the NCRM-1 cells remained in their original spent medium in a continuous bioreactor run with additional Pancreatic Progenitor (PP) stage factors being spiked in (Fig. 1A). It was also noted that aggregate diameter remained consistent for both cell lines throughout PP induction (Fig. 1B–C).
Subsequently, cells were cultured in endocrine inducing media for 10–30 days, consisting of TB2 media supplemented with gamma secretase and A8301. Media changes occurred for all bioreactors except the NCRM-1 cells that had never received a media exchange. NCRM-1 kept the same medium with spiking in additional factors needed for endocrine induction. In this study, several parameters and changes were identified and deemed essential for optimizing efficient large-scale manufacturing of islet-like clusters. Some of these parameters include controlling aggregate size without dissociation, altering basal media to support aggregation, conducting a 100% suspension protocol, extending the PP induction and endocrine induction stages differentiation periods, and limiting basal media changes throughout the process.
Optimization of time through a stage-wise approachCell evaluation occurred at the end of each differentiation stage. Prior to bioreactor seeding, cells were assessed for pluripotency markers SSEA4 and TRA-1–60 (Figure S2A). Populations exceeded 90% for both markers and over 85% co-expression. At the conclusion of DFE stage, FOXA2 expression ranged from 50 to 85%, with no detectable OCT4 expression, signifying the absence of undifferentiated cells (Figure S2B). Immunostaining on bioreactor clusters, which were plated on a vitronectin coated 24 well plate, revealed co-expression of HNF1β and FOXA2 (Figure S2C). HNF1β was originally optimized as a marker in the adherent protocol for directing dorsal foregut endoderm differentiation and subsequent endocrine cell fates [18].
To optimize PDX1 expression, cells remained as clusters in PP stage media for 4–8 days. This time study on PP stage cells determined the optimal duration for differentiation in a suspension environment. Bioreactor PP stage cells were sampled at various time points. While the control sample remained in stage 2 (PP) for only 4 days, two additional samples were kept in PP induction stage media for 6 and 8 days, respectively. Subsequently, the cells were subjected to endocrine induction medium for 10 days and then analyzed for the expression of several endocrine specific markers including GCG, INS, SST, NKX6.1, NKX2.2, and FOXA2 (Fig. 2A). Prolonging the exposure of the PP inducing media led to increased expression of all the afore-mentioned markers except for SST. Notably, the extra two days in the PP inducing media resulted in increased expression of GCG, SST, NKX6.1, and NKX2.2 as compared to both the 4- and 8-day induction periods. It was noted that the expression of INS and FOXA2 continued to increase over time. During the endocrine induction period, marker expression analysis was performed weekly. Prolonging the exposure to the endocrine inducing medium resulted in increased expression of INS, MAFA, MFN1, NKX6.1, and PDX1.Only SST and NEUROD1 expression decreased over time (Fig. 2B). Additionally, daily sampling after 10 days in endocrine induction media revealed that extending this time by at least an additional five days resulted in a 20-fold increase in C-PEP expression. Glucose stimulated insulin release assays conducted over this 5-day time-period showed a minimal increase in C-PEP on day 10, and the largest increase was observed on day 13. Further increases following depolarization with KCL on day 15 showed a significantly higher C-PEP content (~ tenfold) as compared to the control samples performed on day 10 of endocrine inducing stage 3 media (Fig. 2D).
Fig. 2
Time study evaluating differentiation and function. A Gene expression profile from the end of stage 3 of harvested cells that had different prolonged PP induction timing of 4 days, 6 days, and 8 days B Gene expression profile of harvested endocrine cells with increasing 1-week intervals. C C-Peptide concentration per aggregate count as a function of time in endocrine induction media. D Glucose-Stimulated Insulin Secretion of endocrine aggregates as a function of time. The stimulation index for day 11, 12, 13, 14, 15 after the end of endocrine induction (10 days), was 1.4, 1.2, 1.8, 1.2 and 1.4 respectively. All bar charts show individual points with mean ± SD. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001
Characterization of stage-specific markers throughout the protocolThe transition of β-cells during maturation involves notable alterations in the expression levels of HK2 (Hexokinase 2), LDHA (Lactate Dehydrogenase A), and SLC16A1 (Monocarboxylate Transporter 1) [19, 39, 40]. These genes play crucial roles in glucose metabolism and lactate production within cells. As β-cells mature, LDHA expression declines, leading to decreased lactate production. Additionally, SLC16A1 expression decreases, limiting the capacity for lactate transport further contributing to the metabolic shift from lactate generation to pyruvate utilization. This causes a β-cell maturation event by facilitating functional adaptation to full glucose utilization [19].
Comparing the original adherent protocol to the bioreactor production of β-cells over time focusing on these three genes (Fig. 3A) showed that these ‘functional disallowed genes’ are repressed throughout differentiation process within bioreactors. The difference between adherent and bioreactor culture becomes especially significant during the PP-induction stage and throughout the endocrine induction stage, where suspension cultures show significant repression of these glycolytic genes. These results match the findings shown in our RNA sequencing analysis presented in (Figure S1) which suggested that adherent culture lags in differentiation. To determine the shift in gene expression occurring throughout the differentiation protocol, key insulin transcription factors such as NEUROD1, NEUROG3, NKX6.1, NKX2.2, and MAFA, which play crucial roles in β-cell maturation [19, 39, 41,42,43], were monitored throughout the entire process (Fig. 3B). NKX6.1 expression increased after the media changed into the endocrine induction media. Conversely, NEUROD1, NEUROG3, NKX2.2, and MAFA expression began to significantly increase as the duration of the endocrine induction media was lengthened, suggesting a maturation benefit for extended differentiation time. Throughout the culture, FOXA2 and HNF1β were also tracked. FOXA2 regulates gene expression crucial for β-cell development during embryonic pancreatic development [18], and HNF1β dives exocrine development [18]. HNF1β exhibited a significant increase in expression levels between the differentiation from DFE to a PP but had a diminishing expression after endocrine induction. This was expected since its continued expression leads to exocrine development. In contrast, FOXA2 continued to rise post endocrine induction since it governs the expression of key genes like PDX1 and maintains the differentiated state and functionality of mature β-cells. Additionally, the expression levels of INS and GCG were mapped throughout the process, both showing significant increases during endocrine induction, in agreement with the discussed results.
Fig. 3
Characterization of glycolytic and stage specific markers throughout the differentiation protocol. A Glycolytic genes expression comparison between adherent and suspension culture over time. Comparisons are made at the end of each differentiation stage. B Gene expression profile of cells at different stages of the protocol. The stage being assayed is indicated below the graph. All bar charts show individual points with mean ± SD. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001
The iPSC-derived islet-like clusters generated from the bioreactors were assessed for endocrine marker expression through immunostaining of the clusters. It was revealed that the aggregates expressed several known pancreatic markers, including PDX1, C-PEP, CGA, GCG, and SST, indicating the induction of a genuine pancreatic endocrine state (Figure S3A). Some co-expression of endocrine products was noted with the presence of CPEP + /SST + and CPEP + /GCG + cells being observed.
Continuous culture dramatically impacts the differentiation towards pancreatic progenitorsMaintaining stable cellular homeostasis, characterized by minimal fluctuations, is a prerequisite for optimal cellular function and environment understanding which leads to better system control [44]. This stability ensures that processes such as metabolism, signaling, and gene expression remain finely regulated within narrow ranges conducive to cellular function. Multiple factors contribute to the fluctuation and homeostasis and not just a small number of regulatory enzymes [45,46,47]. Factors include growth conditions, glucose concentration, cellular signaling pathways like AKT1, enzymatic regulation by LDH, PFK, and PEP, oxygen availability, cellular metabolism, tissue-specific functions, metabolic shifts, all collectively shaping the balance between lactate production and consumption [44]. To better understand the full bioprocess and nutrient limitations in maintaining a cellular homeostatic environment within the culture, a continuous media study was conducted throughout the differentiation protocol (Figure S3B). The stage-specific utilization of glucose and lactate generation throughout the culture time was assessed. A comparison was made between the Control media change bioreactor and the continuous bioreactor at the end of each stage and every week after endocrine induction. Spent media samples collected throughout the differentiation protocol were evaluated (Fig. 4A). Initially, the glucose concentration in the Control media change bioreactor decreased by approximately 30%. By around day 17 of the protocol, this decrease became less than approximately 16%, eventually reaching around 11% at harvest. With each media change, a steady increase in lactate concentration in the basal medium was observed. Toward the end of the control protocol which received full media changes throughout the process, an accumulation of (6 mM) lactate was measured. Comparing the profile of glucose and lactate change over time between the Control media change bioreactor and the continuous condition revealed significant differences. The glucose consumption rate decreased from approximately 30% before the initiation of differentiation to negligible levels toward the end of the protocol, stabilizing at around 7 mM after approximately 5 days from the start of differentiation (Fig. 4A). Similarly, lactate accumulation increased only during the first 3–5 days of the protocol, reaching a steady state after 5 days in culture at around 17 mM.
Fig. 4
Impact of glucose consumption and lactate accumulation and profile on the differentiation and growth of cells A. Glucose and lactate concentration profile throughout the differentiation protocol on two bioreactors: the control which has frequent media changes at the different stages and the continuous which has no media changes but only spike in of differentiation factors at each stage. B Gene expression profile of cells for stage specific markers on both bioreactors after 20 days in stage 3 endocrine induction media. C Cell growth profile throughout the differentiation period on both control and continuous bioreactors. D The change of glucose over lactate concentration over time for the continuous bioreactor. All bar charts show individual points with mean ± SD. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001
The impact of not changing the basal medium and spiking differentiation factors in a continuous culture of cells was assessed alongside the Control media change bioreactors that underwent the process previously described (Fig. 1A). A comparison of expression levels at late endocrine induction stage revealed significant differences in the levels of PDX1, NEUROD1, and SST between the control cells and the cells that remained in the same medium with only spiking in stage-specific differentiation factors (Fig. 4B). The cells that were not subjected to basal media change exhibited higher expressions of PDX1, NEUROD1, INS, and NKX2.2 but lower expression of SST and GCG. This suggests an increased preference for β-cells throughout the differentiation process.
To explore the metabolic phenotype of the islet aggregates generated by our suspension protocol, we examined the growth rate of cells in both cultures (Fig. 4C) along with the consumption rate of glucose relative to lactate production over the culture period (Fig. 4D). The results further suggest that the cells are potentially primarily relying on glycolysis during the early stages of differentiation (S1-2) and transition to oxidative phosphorylation towards endocrine induction during the protocol (Fig. 4D). This transition occurs earlier in the continuous bioreactor because the glucose consumption rate reached a steady state faster. Endocrine cells produced from a continuous bioreactor run were assessed for INS expression using a HD-DoE assay that included multiple secretagogues (Figure S4A). This demonstrated that a fivefold increase in insulin levels can be attained within a 3-h incubation using the combinatorial influences of high glucose Rapamycin, DCA, oxytocin, and arginine (Figure S4B). The baseline medium on the cells was at low glucose whereas high glucose (17.5 mM) was incorporated as an additive in the design.
Nutrient consumption and metabolite measurementsExamining potential alternative fuel and nutrients sources for the cell, the amino acid profile throughout the culture of both the control and the continuous bioreactors was analyzed (Figure S5A-B). Amino acid concentrations were measured using the REBEL Cell Culture Analyzer (908 Devices). Essential amino acids such as histidine, isoleucine, leucine, lysine, methionine, phenylalanine, threonine, tryptophan, and valine were maintained throughout the culture period in both bioreactors. However, some amino acids were completely depleted in both culture mediums, including L-aspartic acid after 5 days and L-glutamic acid after 16 days. Amino acid metabolism is crucial for normal pancreatic β-cell function, and alanine and glutamine are known for their role in regulating β-cell function and insulin secretion [48]. At the end of culture, glutamine and alanine concentrations were at a higher concentration than they occur in fresh media, suggesting they were not growth-limiting (Figure S5A-B). However, the source of their increase remains unknown, unlike previous observations attributing their increase to the GlutaMAX™ additive [49]. This increased level of alanine and glutamine compared to the starting medium was not observed in the Control bioreactor that had frequent media changes between stages and throughout the extended endocrine induction stage. No other significant differences were observed between the two bioreactors. As previously noted, the bioreactor with limited media replenishment showed better differentiation than the Control media change bioreactor. Dietary manipulations of amino acids and serum deprivation have been linked to promoting adult-like traits in pancreas β-cells derived from human stem cells [34, 50,51,52].
In addition, both culture bio-profiles were assessed using FLEX2 (Nova Biomedical), analyzing Gln, Glu, NH4 +, Na +, K +, Ca + +, pH, PCO2, and PO2 throughout the culture period for both reactors (Figure S6A-B). The osmolarity of the culture medium steadily increased in the continuous bioreactor but remained within the range of 280–320 mOsm/kg. This increase can be attributed to the accumulation of solutes from nutrient metabolism and other waste products. In contrast, the osmolarity of the Control media change bioreactor fluctuated as the media was replenished at different stages of the differentiation process. Glutamine and glutamate levels were also assessed, and both showed depletion over time. This was consistent with measurements taken using the REBEL analyzer. Both bioreactors were comparable in their bio-profile, except for major differences observed in the continuous decrease of pH in the continuous bioreactor, as expected, and the rate of oxygen consumption. The gases measured in the media may have been impacted by the time between collection and measurement, however, the overall impact is the same for all samples. The overall data profile showed that the PO2 level began steadily decreasing after 10 days of culture or PP induction stage of differentiation. Although the interface of the media with the gas in the headspace of both bioreactors is the same since they are both the same size (500 ml), the near-equivalent flux of oxygen into the media may not be sufficient to replenish increased oxygen consumption in the 0.5L continuous vessel as compared to the Control media change bioreactor that simply has all the media replaced at regular intervals.
Validation and scalingAfter resuming iterative process optimization and improvement efforts and implementing them as needed to achieve the desired culture outcomes, the process was validated using multiple bioreactors at different densities. Three different seeding densities were used to seed 0.5L bioreactors (75 M, 90 M and 120 M). The aggregates generated were monitored throughout the experiment and harvested at the end of the process (Fig. 5A). The cell clusters were evaluated for staining with dithizone (DTZ, which binds zinc within insulin granules), flow cytometry and protein levels of selected markers, viability, and oxygen consumption rate. There was a variable retention of DTZ throughout the different runs with the strongest intensity observed in Endo75A’ while little if any was observed in Endo75B’ (Fig. 5B). The aggregates were then analyzed for viability (Fig. 5C) and size (Fig. 5D). The cells were viable after the induction of endocrine cells, and the culture aggregate diameter average was below 500 µm. The growth rate of the bioreactors followed a profile of rapid proliferation during the early stages of differentiation (Generation of PP) and plateauing after endocrine induction (Fig. 5E). A sample of the clusters was digested to be further tested for counts (Fig. 5F) and flow cytometry (Figure S7–10). As expected, the largest yield was observed in bioreactors with the largest seeding density. These bioreactors also had the largest aggregate diameter; however, this didn’t impact viability (Fig. 5E).
Fig. 5
Scale-up validation on multiple 0.5L bioreactors with different seeding densities. A An image of all bioreactors runs for this validation. B Dithizone staining of endocrine aggregates products. C Live dead staining on endocrine stage aggregates generated from all bioreactors using fluorescein diacetate for live staining and propidium iodide for dead staining. D Violin plot of the aggregate diameter variance on endocrine stage aggregate from all bioreactors runs including previous control. E The growth rate of cells over time for the Control media change bioreactor Endo75A. F Table summary of the viability of digested endocrine aggregates and their respective total cell count from each 500 ml bioreactor
The continuous and Control media change bioreactors were tested for OCR and ECAR (Fig. 6A). Consistent with our findings, iPSCs-derived pancreatic progenitors produced with constant medium changes have lower OCR compared to those produced in a continuous bioreactor. iPSC-islets generated with our continuous bioreactor protocol have a higher aerobic respiration capacity. Mature function is dependent on obligatory aerobic metabolism and an increase in aerobic capacity of iPSC-derived islets is suggestive of an increased functionality, though full function has not been obtained. To assess the robustness of the continuous bioreactor process we next performed a continuous run using three different cell iPSC lines (Figure S11). It was shown that the process itself worked for all cell lines, however the efficacy of the pancreatic induction protocol varied widely. The early endodermal genes FOXA2 and HNF1β were all induced to similar levels, however genes representative of a pancreatic progenitor state showed variance between the cell-lines (Figure S11A). PDX1 induction was detected in all three cell lines, but was significantly reduced in the RCRP5005N lines, whereas NKX6.1 was significantly reduced in the NCRM-4 line. Consequently, endocrine induction was severely reduced with only SST reaching similar induction levels in the NCRM-4 cell line. Immunofluorescent staining was performed on the three continuous bioreactor runs to confirm the reduced induction levels of PDX1 and CPEP in the different iPSC lines (Figure S11B).
Fig. 6
Energy map profile and cryopreservation recovery on endocrine cells. A Seahorse data on OCR and their respective ECAR profiles on a control run with regular media changes versus a continuous bioreactor run. B Live dead staining using fluorescein diacetate for live staining and propidium iodide for dead staining. On endocrine aggregates that were recovered from cryopreservation. C-peptide concentration profile normalized to the number of aggregates on cryopreserved and recovered cells from bioreactors. OCR and ECAR charts show individual points with mean ± SD. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001
Cryopreservation and recoveryThe harvested aggregates were cryopreserved in CS10 solution with 10% DMSO in liquid nitrogen. Some vials were then recovered in TB2 medium + 10 µM Y-27632 ROCK inhibitor as aggregates. The clusters were checked for viability using FDA/PI stain immediately, and after 5 days of recovery, (Fig. 6B) no change was observed. The cells were > 85% viable after recovery. In addition, the cells were evaluated for C-PEP content in high glucose medium (17.5 mM) over time using a C-PEP Elisa kit. The concentration was then normalized per aggregate number (Fig. 6C). The C-PEP content was ~ 22 pmol per aggregate before cryopreservation. It decreased immediately after recovery but then reached a steady state after 5 days and stayed at 5 pmol per aggregate for the next 10 days.
留言 (0)