
記住我

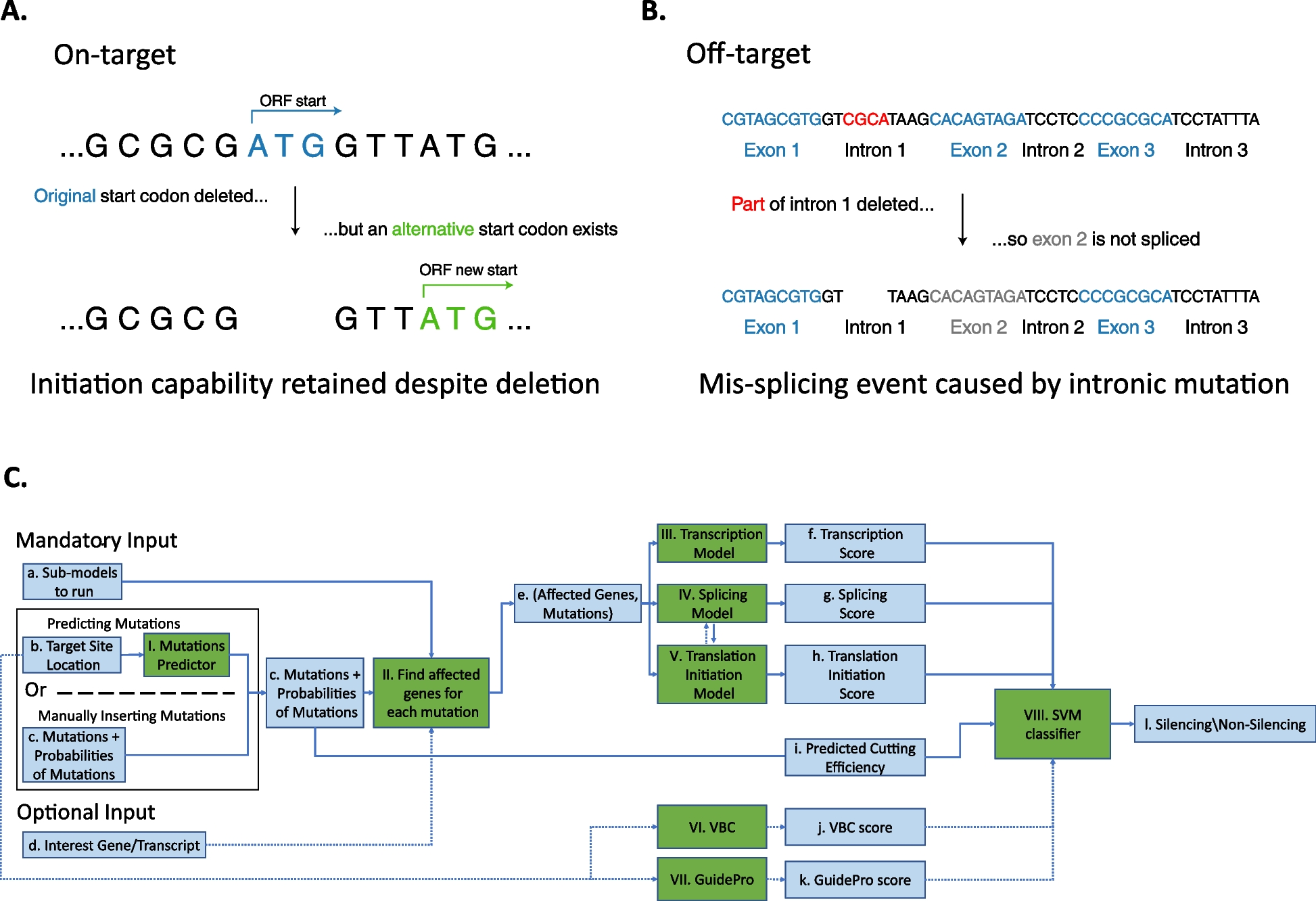


Auditory brainstem response (ABR) recordings showed that Mir96+13G>A and Mir96+14C>A homozygotes exhibit profound deafness at all ages tested, showing no response at the highest sound level tested (95 dB sound pressure level (SPL)) at any of the ages studied (14 days to 6 months old) (Fig. 2). Mir96+14C>A heterozygous mice have mild progressive hearing loss most pronounced at high frequencies and progressing with age to lower frequencies (Fig. 2B), which correlates with the human phenotypes [5, 16]. However, Mir96+13G>A heterozygous mice have normal hearing up to 6 months old (Fig. 2A), which does not mimic the phenotype of humans with the equivalent mutation in heterozygosis [5, 16].
Fig. 2
ABR thresholds of Mir96+13G>A (A) and Mir96+14C>A (B) mice at ages from postnatal day (P)14 to P180. ABR measurements in response to click stimuli and tone pips ranging from 3 to 42 kHz were recorded from anaesthetised mice at different ages from P14 to P180. Homozygous mice for either mutation are shown as red triangles, heterozygotes as blue squares, and wildtypes as black circles. Each point in the plot shows the mean of the lowest stimulus level (threshold) at which a response is observed, ± SD. Points at 95 dB sound pressure level (SPL) indicate that there is no response up to the maximum sound level used. Both males and females were tested and are plotted together; n numbers are shown on each plot. Asterisks indicate significant differences (Bonferroni-corrected p < 0.05, mixed linear model pairwise comparison) between heterozygous mice and wildtypes (blue) or homozygous mice and wildtypes (red)
The mice were generated and maintained on the C57BL/6N genetic background. C57BL/6N mice are known to have age-related hearing loss, which is caused in part by the Cdh23ahl allele [39]. At 4 weeks of age, the high frequencies are affected, whereas the lower frequencies remain unaffected up to 6 months of age [40]. We observed a similar pattern in wildtype mice from the Mir96+13G>A and Mir96+14C>A lines, which showed mild hearing loss at 24–42 kHz from 8 weeks old, but retained good low-frequency hearing sensitivity up to 6 months old (Fig. 2).
Mir96 +13G>A heterozygotes have normal waveforms and retain normal hearing on a different genetic background and when subjected to noiseIn order to further investigate the hearing of Mir96+13G>A heterozygotes, we first plotted their waveforms at 6 months old and found no obvious difference when compared to the wildtype waveforms (Fig. 3A). Mice were aged to 1 year old, but there was still no difference in the thresholds (Fig. 3B).
Fig. 3
Further electrophysiological investigation of the Mir96+13G>A heterozygous mice. A Mean ABR waveforms at 12 kHz, shown at 20 dB (top) and 50 dB (bottom) above threshold (sensation level, SL) ± standard deviation, at 6 months old. There is no obvious difference in waveform between Mir96+13G>A heterozygous (blue, n = 20) and wildtype mice (black, n = 14). B Mean ABR thresholds from Mir96+13G>A homozygous (red triangles), heterozygous (blue squares) and wildtype (black circles) mice at 9 months and 1 year old. Males and females are plotted together; n numbers are shown on each plot. C Mean ABR thresholds from Mir96+13G>A homozygous (red triangles), heterozygous (blue squares) and wildtype (black circles) mice on a mixed C3HeB/FeJ, C57BL/6N background at ages from 3 weeks to 6 months old. Males and females are plotted together; n numbers are shown on each plot. For B and C, asterisks indicate significant differences (Bonferroni-corrected p < 0.05, mixed linear model pairwise comparison) between heterozygous mice and wildtypes (blue) or homozygous mice and wildtypes (red). D Mean ABR thresholds from Mir96+13G>A heterozygous and wildtype mice before and at multiple time points after noise exposure, showing similar recovery of thresholds in wildtype (brown circles, n = 5) and heterozygous (purple squares, n = 5) mice. Control wildtype (black circles, n = 5) and heterozygous (blue squares, n = 5) littermates went through the same set of ABR measurements and spent the same time in the noise exposure chamber, but without the noise. Males and females are plotted together. Asterisks indicate significant differences (Bonferroni-corrected p < 0.05, mixed linear model pairwise comparison) between unexposed wildtype and unexposed heterozygous mice (blue) or noise-exposed wildtype and noise-exposed heterozygous mice (purple). Error bars in all panels are standard deviation
Next the hearing of wildtype, heterozygous and homozygous Mir96+13G>A mice on a mixed C57BL/6N, C3HeB/FeJ background was tested. The C3HeB/FeJ strain was selected for this outcross because it is known to show good hearing into old age and was the genetic background of the Mir96Dmdo mutant line previously studied [4]. However, on the mixed background no difference was found between wildtypes and heterozygotes up to 6 months old, while homozygotes were still profoundly deaf at all ages tested (Fig. 3C).
Finally, Mir96+13G>A heterozygote and wildtype mice on the original C57BL/6N background were subjected to 100 dB SPL, 8–16 kHz noise for 1 h to ask if the heterozygotes were more sensitive to noise damage than wildtypes. The noise-exposed mice exhibited a threshold shift which gradually recovered over the subsequent 4 weeks, but there was no obvious difference in thresholds or in threshold recovery rate between the wildtype and heterozygous mice (Fig. 3D). The difference in the mean thresholds of noise-exposed heterozygotes and wildtypes at 30 kHz 14 and 28 days after noise is statistically significant, but there is a lot of variability between individual mice, so we would not conclude that this is a biologically relevant difference. While the Mir96+13G>A heterozygous non-exposed mice do appear to have worse thresholds than the non-exposed wildtypes, this is because these five wildtype mice have better high-frequency thresholds than the mice which went through the initial ABR tests (Fig. 2).
Mir96 +13G>A and Mir96 +14C>A homozygous mice have severely affected stereocilia bundlesScanning electron microscopy (SEM) showed that homozygous mice of either mutation have severely affected stereocilia bundles at postnatal day 28 (P28). Stereocilia defects were seen in both outer hair cells (OHCs) and IHCs and were more severe in Mir96+14C>A than in the Mir96+13G>A mutants. We observe disorganised stereocilia that had lost their normal staircase arrangement, a fusion of stereocilia, and giant stereocilia, particularly in the IHCs (Fig. 4I, N, O), as well as missing stereocilia bundles (Fig. 4H, K, N). That phenotype becomes more striking at high frequency regions (Additional file 3: Fig. S1).
Fig. 4
Scanning electron micrographs of Mir96+13G>A (D–J) and Mir96+14C>A (K–P) mice at 4 weeks old. Representative examples of wildtype (A–C), heterozygous (D–F, K–M) and homozygous (H–J, N–P) mice are shown. All images correspond to the 12 kHz best frequency region. For each panel, the left column (A, D, H, K, N) shows a zoomed-out image with inner and outer hair cell rows. The middle column (B, E, I, L, O) shows an inner hair cell close up, and the right column (C, F, J, M, P) shows an outer hair cell close up. Mir96+13G>A mice: wildtype (n = 4), heterozygote (n = 3), homozygote (n = 3). Mir96+14C>A mice: wildtype (n = 2), heterozygote (n = 6), homozygote (n = 3). Arrowheads in F point to the loss of stereocilia in the shortest row of the OHC bundles in Mir96+13G>A heterozygotes. Scale bar on left hand panels = 10 µm; scale bar for single hair cells = 1 µm
In the heterozygotes, stereocilia damage is less severe than in the homozygotes. Mir96+13G>A heterozygous mice, despite having normal ABR thresholds, show loss of some stereocilia in the shortest row of the OHC bundles, which worsens at higher frequency regions (Fig. 4F). In Mir96+14C>A heterozygotes, which have mild hearing loss by P28, some OHC stereocilia bundles have a U-shape, instead of the typical V-shape observed in wildtypes; this rounding is more often seen towards the apical turn of the cochlea (Fig. 4M). In inner hair cells of Mir96+14C>A heterozygotes, some of the stereocilia appeared to taper towards their tips (Fig. 4L).
Mir96 +13G>A homozygous mice show a reduction in the number of IHC synapsesConfocal microscopy was used following immunolabelling of the pre- and postsynaptic markers CTBP2 and GRIA2 (GluR2), respectively, to study the synapses at 4 weeks old (Fig. 5A). Mir96+13G>A homozygotes were found to have a significant reduction in the number of ribbon synapses (defined as colocalised pre- and postsynaptic labelling) per inner hair cell compared to wildtypes at 4 weeks old (Fig. 5B). There were no differences in heterozygotes compared to wildtypes. Furthermore, no significant differences were found in the number of synapses in Mir96+14C>A wildtypes, heterozygotes and homozygotes (Fig. 5C).
Fig. 5
Analysis of synapses in Mir96+13G>A and Mir96+14C>A mutant mice at 4 weeks old. A Confocal images of the whole-mount organ of Corti. Synapses were examined using an anti-CtBP2 antibody to mark pre-synaptic ribbons (pink) and an anti-GluR2 antibody to mark postsynaptic densities (green). Nuclei are shown in blue (DAPI). The images correspond to the cochlear region of 12 kHz best frequency. Scale bar = 5 µm. Insets in the Mir96+13A>G wildtype and homozygote panels show enlarged examples of a colocalised synapse (wildtype) and orphan pre- and postsynaptic labels (homozygote). Quantification of ribbon synapses per IHC in Mir96+13G>A (B) and Mir96+14C>A (C). Confocal z-stacks were obtained with a z-step size of 0.25 µm and maximum intensity projection images were used for synapse counting. Colocalised pre- and postsynaptic components were defined as a synapse. Synapses were counted and divided by the number of IHCs, determined by Myo7a staining (not shown in the images, only used for quantification purposes). All data are shown as mean ± SD and statistically analysed by one-way ANOVA with Tukey’s multiple comparisons test (** = p < 0.01). Mir96+13G>A mice: wildtype (n = 4), heterozygote (n = 5), homozygote (n = 3); p = 0.0016 (wildtype vs heterozygote adj. p = 0.65; heterozygote vs homozygote adj. p = 0.0041; wildtype vs homozygote adj. p = 0.0018). Mir96+14C>A mice: wildtype (n = 5), heterozygote (n = 3), homozygote (n = 2); p = 0.18
Ocm and Slc26a5 are downregulated in Mir96 +13G>A and Mir96 +14C>A homozygotesOcm and Slc26a5 are two genes that are strongly expressed in normal outer hair cells and both were strongly downregulated in Mir96Dmdo mice [4] and in Mir183/96dko mice [12]; neither are predicted targets of miR-96, nor of any of the mutant miR-96 forms. Therefore, we measured expression levels of these two genes in the two new mutants studied here using qPCR on RNA from the organ of Corti at P4, and both genes were found to be significantly downregulated in Mir96+13G>A and Mir96+14C>A homozygotes (Fig. 6).
Fig. 6
Ocm and Slc26a5 are downregulated in Mir96+13G>A and Mir96+14C>A mutants. RT-qPCR was carried out on cDNA from the organ of Corti of P4 wildtype (grey) and homozygous (red) littermates to test gene expression changes. Mean expression levels were calculated from individual expression levels from each mouse and normalised to expression in a wildtype littermate. Relative expression levels were determined using the 2−ΔΔct equation [23], using Prox1 as an internal control for the amount of organ of Corti tissue present because Ocm and Slc26a5 are specifically expressed in hair cells whereas Prox1 is expressed in supporting cells of the organ of Corti [24]. Error bars represent the standard deviation (Wilcoxon test; * = p < 0.05). Mir96+13G>A Ocm = 6 wildtypes, 6 homozygotes, p = 0.0022; Slc26a5 n = 4 wildtypes, 4 homozygotes, p = 0.029. Mir96+14C>A Ocm = 3 wildtypes, 7 homozygotes, p = 0.017; Slc26a5 n = 3 wildtypes, 7 homozygotes, p = 0.017. At least three technical replicates were used for each experiment
Mir96 +14C>A homozygous mice have a larger number of differentially expressed genes than Mir96 +13G>A homozygotesIn order to further explore the transcriptome of these mice, we carried out RNA-seq analysis of the organ of Corti of homozygotes and wildtype sex-matched littermates at P4. This age was chosen because earlier analyses had shown that all hair cells were still present at this early age, and we wanted to detect expression changes reflecting the different genotypes of the mice rather than due simply to reduced numbers of the cell type of interest (hair cells). Our results revealed that many genes are significantly dysregulated (FDR < 0.05) in the organ of Corti of Mir96+13G>A and Mir96+14C>A homozygous mice, confirming that miR-96 controls a complex network of genes in the inner ear (Fig. 7, Additional file 4: Table S3). Mir96+13G>A homozygotes have 328 significantly differentially expressed genes (DEGs), 203 of which are upregulated and 125 downregulated. In Mir96+14C>A mutants, a total of 693 genes are significantly differentially expressed, with 369 upregulated and 324 downregulated. The much larger number of DEGs in Mir96+14C>A homozygous mice compared to Mir96+13G>A homozygotes correlates with the more severe structural phenotype observed in these mutants (Fig. 4). The two mutants share only 124 DEGs misregulated in the same direction, indicating that transcriptomic changes due to each mutation are very different, even though both mutations are only one base apart in the Mir96 sequence (Fig. 7, Additional file 4: Table S3).
Fig. 7
Comparison of up- and downregulated genes in Mir96+13G>A and Mir96+14C>A homozygotes. The majority of differentially expressed genes (DEGs) are not shared. Three genes are upregulated in Mir96+14C>A and downregulated in Mir96+13G>A homozygotes
Sylamer analysis identifies both loss of wildtype targets and gain of novel targets in each mutantWe used Sylamer [30] to assess the impact of the two mutations on the mRNA profile of both mutants. Sylamer counts the occurrences of all possible heptamers in the 3′UTRs of the genes in the RNA-seq datasets, considered in order from most upregulated to most downregulated, and measures the enrichment of each heptamer throughout the ranked gene list. We found that the complementary heptamer to the miR-96 seed region (GTGCCAA, red line) was greatly enriched in the 3′UTRs of hundreds of genes upregulated in Mir96+13G>A and Mir96+14C>A homozygotes (Fig. 8). This indicates that miR-96 normally represses a wide range of target genes, and when it is mutated, it is not able to repress its targets, which then become upregulated.
Fig. 8
Sylamer enrichment landscape plots for Mir96+13G>A and Mir96+14C>A homozygous mice. The plots show enrichment and depletion of heptamers in the 3′UTRs of the differentially expressed genes (DEGs). The x-axis shows the list of DEGs for each mutant ranked from most upregulated (left) to most downregulated (right), based on the fold change (logFC). The Y axis shows the hypergeometric significance for the enrichment or depletion of heptamers in 3′UTRs in the leading parts of the gene list. Positive values indicate enrichment and negative values indicate depletion. The grey lines show the profiles of unenriched heptamers, while the coloured lines represent heptamers that are complementary to the seed regions of wildtype and mutant miR-96. Sylamer measures the enrichment of every possible heptamer in the 3′UTRs of the genes, in cumulative bins of 500 (x-axis), and generates a landscape plot. In this case, the main peaks in both mutants are from the wildtype seed (red) and show that it is enriched in the upregulated genes. The negative peaks on the right hand of each plot indicate that the downregulated genes are enriched in heptamers corresponding to the mutant miR-96. In the Mir96+13G>A graph, the purple line corresponds to the mutant seed region, and in the Mir96+14C>A graph, the main peak of the mutant seed region is indicated in cyan. These negative peaks suggest that the mutant miR-96 is acquiring new targets. Neither of the mutant seed regions is enriched in the other mutant (cyan line in Mir96+13G>A, purple line in Mir96+14C>A)
We then asked whether mutations led to the silencing of potential acquired targets, that is, those genes containing in their 3′UTR binding sites complementary to the mutant seed region of miR-96. We observed that the heptamer complementary to the mutant miR-96 is enriched among the most downregulated genes, indicating that mutant miR-96 influences the expression of newly acquired target genes. The complementary heptamer to the Mir96+13G>A mutant seed region is GTGTCAA (Fig. 8, purple line), and the one complementary to the Mir96+14C>A mutant seed region is GTTCCAA (Fig. 8, cyan line).
The differentially expressed genes in Mir96 +13G>A and Mir96 +14C>A mutants are enriched in specific processesWe carried out gene set enrichment analysis (GSEA) [31, 32] on the transcriptomes from each mutant (Additional file 5: Table S4) and visualised the results in Cytoscape. An enrichment map was generated for each mutant (Additional file 3: Fig. S2) [34]. From the Mir96+13G>A enrichment map (Additional file 3: Fig. S2A), we observed that a large number of DEGs were involved in synaptic activity, with terms like “presynaptic”, “dopamine transport” and “NMDA activation” being enriched, which correlates with the synaptic phenotype in this mutant (Fig. 5). In the Mir96+14C>A network (Additional file 3: Fig. S2B), we observed gene sets involved in functions related to the cytoskeleton, extracellular matrix and adhesion molecules, which may be connected to the severe stereocilia degeneration observed in these mice (Fig. 4).
We also analysed the significantly misregulated genes using Ingenuity Pathway Analysis (Qiagen, Germany), which identified canonical pathways with a significant overlap with the misregulated genes. There were 10 pathways with significant overlaps in the Mir96+13G>A data, including sensory processing of sound by outer and inner hair cells of the cochlea, and several pathways involved in synaptic activity, correlating with the GSEA analysis (Additional file 6: Table S5). From the Mir96+14C>A data, we obtained 45 significant pathways, which may reflect the higher number of significantly differentially expressed genes in this mutant dataset. The pathways included sensory processing of sound, degradation of the extracellular matrix, assembly of collagen fibrils and integrin cell surface interactions, which again correlates with the GSEA analysis (Additional file 6: Table S5).
Comparison of the misregulated genes with Mir96 Dmdo and Mir183/96 dko mutantsOur aim in investigating the transcriptome of these Mir96 mutant mice was to determine candidate proteins with therapeutic potential. However, comparison of DEGs in Mir96+13G>A and Mir96+14C>A with previous data from Mir96Dmdo ([15], Additional file 5: Table S4) and Mir183/96dko [12] (Fig. 9) revealed only six genes shared between the four mutants. Myo3a, Hspa2 and St8sia3 are upregulated, while Tmc1, Slc26a5 and Ocm are downregulated. Some other genes are differentially expressed in just two or three of the mutants, but most of them are differentially expressed in only a specific mutant, highlighting the transcriptomic differences obtained as a consequence of the different mutations and the different approaches. Furthermore, some genes are misregulated in two or more mutants but in different directions. For example, Otof is downregulated in Mir96+13G>A and Mir96+14C>A mutants, but it is upregulated in Mir96Dmdo mice.
Fig. 9
Comparison of the differentially expressed genes between four miR-96 mutants: Mir96+13G>A, Mir96+14C>A, Mir96Dmdo and Mir183/96dko. Downregulated genes are indicated in dark teal, upregulated genes in red, and genes that are upregulated in one mutant and downregulated in another are shown in orange
Identification of candidate miR-96 targetsIdentifying targets of a master regulator is an important step in building a network which can then be assessed for therapeutic potential. We took three approaches to investigate potential direct targets of miR-96 in all four mutant mice.
First, we used Sylamer with different word lengths (6, 7 (as above) and 8) and identified the peak enrichment of the wildtype seed region closest to the start of the ranked genes (ranked from most upregulated to most downregulated). Genes ranking higher than this threshold (that is, to the left of this peak, shown by a vertical line in Additional file 3: Fig. S3) whose 3′UTR sequences contain matches to the miR-96 seed region are candidate targets [30] (Additional file 7: Table S6).
Secondly, we used the gene set enrichment analysis to identify gene sets defined by a common regulator, usually identified by the presence of a binding motif close to their start sites. We performed the same GSEA analysis on the Mir183/96dko and Mir96Dmdo transcriptomes [4, 12] (Additional file 8: Table S7) and tested the identified transcription factors from all four analyses for the presence of wildtype miR-96 seed region matches in their 3′UTRs (Additional file 7: Table S6).
Thirdly, we used Ingenuity Pathway Analysis (IPA) to predict upstream regulators [41] based on the significantly misregulated genes in each of the four mutant miR-96 transcriptomes. From the list of potential upstream regulators, we selected those genes which were predicted to have higher activity in homozygotes and had wildtype miR-96 seed region matches in their 3′ UTRs, and excluded genes known to be downregulated (Additional file 7: Table S6).
Only two proteins were identified as candidates by more than one approach: Pax7 and Atf3, found in both the GSEA and IPA analyses (Additional file 7: Table S6). There were no overlaps between either the GSEA or the IPA target lists and the Sylamer targets.
Identifying candidate therapeutics from whole transcriptome dataBecause we were unable to identify a shortlist of candidate target genes or proteins to assess for therapeutic potential, we chose instead to use the whole transcriptome, comparing it to known drug profiles from DrugMatrix, which contains the results from thousands of experiments treating rats or rat cell lines with drugs [42]. This was carried out using the SPIED (Searchable Platform-Independent Expression Database) platform [43,44,45]. In order to focus on the most important differentially expressed genes and to exclude, as much as possible, genes misregulated because they are novel targets of a specific mutant miR-96 seed, we looked at genes with consistent differential expression between the Mir96+13G>A and the Mir96+14C>A transcriptomes (7944 genes in total, Additional file 9: Table S8). We did not include the other two transcriptomes because they were carried out on different platforms and with different methods, which could result in exclusion of genes for reasons unrelated to their biological relevance (for example, Ccer2, which is downregulated in the Mir183/96dko and the Mir96+14C>A transcriptomes, was not present on the microarray used for the Mir96Dmdo study).
SPIED outputs 100 profiles by default, ranked from the most similar to the input profile to the most dissimilar, and the drugs with the most dissimilar profile are potential therapeutics because they lead to a complementary change in transcription patterns across the 7944 genes tested. Among the anti-correlated drug profiles compared with the profiles from Mir96+13G>A and Mir96+14C>A (Additional file 9: Table S8) is amitriptyline, a tricyclic antidepressant which has previously been reported to improve recovery after noise-induced hearing loss in guinea pigs [46]. Amitriptyline was also identified in the SPIED analysis of the Mir183/96dko transcriptome, which should not include novel targets since the mutant allele is a deletion of Mir96 (Additional file 9: Table S8). It is known to cross the blood–brain barrier [47], which suggests it may also be able to cross the blood-labyrinth barrier. We therefore considered it a good candidate for testing in Mir96 mutant mice.
Amitriptyline delays hearing loss in Mir96 +14C>A heterozygotesWe first tested amitriptyline in wildtype C57BL/6N mice to verify that it did not affect hearing and found that at 4 weeks old there was no difference between wildtype mice drinking water with saccharin and wildtype mice drinking water with saccharin and amitriptyline (Additional file 3: Fig. S4). We then tested Mir96+14C>A heterozygotes and homozygotes, and wildtype littermates, given either saccharin alone, saccharin and 200 µg/ml amitriptyline, or saccharin and 400 µg/ml amitriptyline. We observed progressive hearing loss in the heterozygotes drinking amitriptyline, but it was significantly delayed at high frequencies between 24 and 36 kHz compared to the heterozygotes drinking only saccharin, most visibly at 30 kHz at 4 weeks old (Fig. 10). No improvement was seen in the homozygotes drinking amitriptyline, and increasing the dose of amitriptyline to 400 µg/ml made no difference to the hearing impairment in either heterozygotes or homozygotes (Fig. 10).
Fig. 10
Amitriptyline delays the progression of hearing loss in Mir96+14C>A heterozygotes. Mean ABR thresholds from Mir96+14C>A heterozygous mice and wildtype littermates drinking either saccharin (open squares), 200 µg/ml amitriptyline plus saccharin (circles) or 400 µg/ml amitriptyline plus saccharin (diamonds) at ages from 21 days to 6 months. Asterisks indicate significant differences (Bonferroni-corrected p < 0.05, mixed linear model pairwise comparison) between mice drinking either amitriptyline or double-dose amitriptyline (2 × Ami) compared to mice of the same genotype drinking saccharin. Significant differences between heterozygotes on differing dosages are marked in blue, and significant differences between wildtypes on different dosages are marked in black. The final plot shows mean thresholds at 30 kHz plotted against time. Error bars are standard deviation
留言 (0)