
記住我






Sepsis represents a significant global health challenge, impacting nearly 50 million people and accounting for approximately 20% of all deaths worldwide (1). Despite advances in understanding its pathophysiology, septic shock often results in mortalities of 30% or higher. Management of sepsis is generally supportive. While fluids and antibiotics improve outcome, no pharmacologic therapy has been shown to effectively target the dysregulated host response seen in sepsis (2, 3). While numerous preclinical studies have shown promise in improving outcomes based upon sound mechanistic insights, effective translation of specific treatments for sepsis into clinical practice at the bedside has proven to be an elusive goal (4). Consequently, there is a clear need to develop targeted and effective therapeutics to augment current strategies in sepsis management.
AMP-activated protein kinase (AMPK) is a central integrator of cellular energy and metabolic homeostasis (5). Once activated, AMPK participates in triggering downstream effector proteins involved in a range of biological responses, from glucose metabolism and lipid oxidation to autophagy (6, 7). Although AMPK activation is best known for its role in long-term treatment of diabetes through drugs such as metformin, growing evidence suggests that AMPK activation can exert anti-inflammatory effects through several mechanisms. As such, AMPK is increasingly viewed as a potential therapeutic target for a number of human conditions, including cardiovascular diseases, ischemia-reperfusion injuries, and various inflammatory diseases. Numerous pharmacological agents have been identified that can activate AMPK, ranging from widely used medications to innovative substrates to natural compounds.
In critically ill patients, multiple meta-analyses have demonstrated an association between preadmission use of metformin, an indirect AMPK activator, and lower mortality in adult septic patients who have diabetes mellitus (8, 9). Further, a propensity score matched cohort of 2,691 septic ICU patients with type 2 diabetes mellitus demonstrated decreased 90 day mortality, reduced severe kidney injury and increased renal recovery in patients exposed to metformin during their hospitalization (10). Additionally, metformin exposure during Staphylococcus aureus bacteremia (regardless of prior use) was shown to be an independent predictor of survival in a study of 452 patients with diabetes (11). Understanding the limitations of associative studies and that causation should not be inferred from these studies, activation of AMPK may represent a therapeutic strategy for the treatment of sepsis that is worthy of further study. In fact, a protocol has recently been published for a planned randomized clinical trial examining the safety and feasibility of metformin as a treatment for sepsis-associated acute kidney injury (12). While metformin is well known as a treatment for diabetes mellitus, AMPK activation has numerous effects above and beyond this, and the full spectrum of effects of AMPK activation during sepsis remains to be fully elucidated. This review explores the current understanding of the role of AMPK in sepsis, emphasizing the therapeutic implications of AMPK activators.
Overview of AMPKAMPK functions as an energy sensor that is activated in response to cellular energy depletion. Activation is triggered by increased AMP/ATP or ADP/ATP ratios during conditions such as starvation, hypoxia, ischemic stress, or exercise. Structurally, AMPK is a heterotrimeric complex comprised of three subunits: the catalytic α-subunit, the scaffolding β-subunit, and the regulatory γ-subunit. There are two isoforms of the α and β subunits, while the γ-subunit has three isoforms (13, 14). The α1, β1, and γ1 subunits are ubiquitously expressed, whereas other combinations display tissue-specific expression. For instance, the α2 and β2 subunits are predominantly found in heart and skeletal muscle (15). Primary physiological AMPK activation is achieved by phosphorylation at Thr172 of the AMPK α-subunit predominantly by upstream kinases including liver kinase B1 (LKB1) and calcium/calmodulin-dependent protein kinase kinase-beta (CaMKKβ) (5, 14). Activation of the LKB1/AMPK signaling pathway reduces lung vascular permeability and the systemic inflammatory response following lipopolysaccharide (LPS) treatment (16). Macrophage LKB1 also helps control local Klebsiella pneumoniae growth during pneumonia by maintaining adequate alveolar macrophages in the lung (17). In contrast, CaMKKβ activates AMPK via an increase in intracellular Ca2+ concentration, independent of the AMP/ATP or ADP/ATP ratios. The CaMKKβ/AMPK pathway is pivotal in providing protection against LPS-induced neuroinflammation and cerebral ischemia/reperfusion injury (18, 19). Activation of AMPK then influences a number of downstream pathways related to carbohydrate, amino acid, and lipid metabolism, as well as mitochondrial function, autophagy, and cell growth (14, 16).
AMPK activatorsAMPK can be activated directly or indirectly by a number of different pharmacological compounds. 5-aminoimidazole-4-carboxamide ribonucleoside (AICAR) is commonly utilized as an experimental direct AMPK activator. AICAR is transformed intracellularly into AICAR monophosphate, which acts as an AMP mimetic, consequently activating AMPK. However, it is worth noting that AICAR also activates other AMP-regulated enzymes, such as fructose-1,6-bisphosphatase through which it could play a role in regulating glycolysis and gluconeogenesis unrelated to AMPK (13). Over the past decade, more specific direct small molecule AMPK activators have been developed. For instance, A-769662 specifically activates AMPK by directly binding to the AMPK β-subunit, resulting in allosteric activation. In contrast, metformin, a frequently prescribed medication for type 2 diabetes mellitus, along with other biguanides, indirectly activates AMPK by inhibiting the mitochondrial respiratory chain complex I, leading to an increase in AMP levels. Additionally, IM156 is a novel biguanide more potent than metformin in activating AMPK. Several natural products derived from plants, such as berberine from traditional Chinese medicine and the polyphenol resveratrol, have also been identified as AMPK activators (20, 21).
Kinetics of AMPK activation during sepsisAMPK activation changes during sepsis differ depending on both tissue and species. AMPK activation is increased at 12 hours and declines at 24 hours following CLP compared with sham control in mouse liver (22). In contrast, AMPK activation is elevated at 6 hours in rat heart and remains elevated at 24 and 72 hours following CLP compared with sham control (23). A trend towards lower AMPK activation is observed in mouse lung 24 hours after CLP (24) while AMPK activation trends higher by 6 hours in mouse kidney after CLP, before returning to baseline (25). AMPK activation in diaphragm and tibialis anterior is also increased 48 and 96 hours after CLP in mice (26). In contrast, human biopsy data demonstrated that in vastus lateral muscle, AMPK activity is reduced in critically ill ICU patients (not restricted to sepsis), with an interquartile range of 4 to 6 days following admission (27).
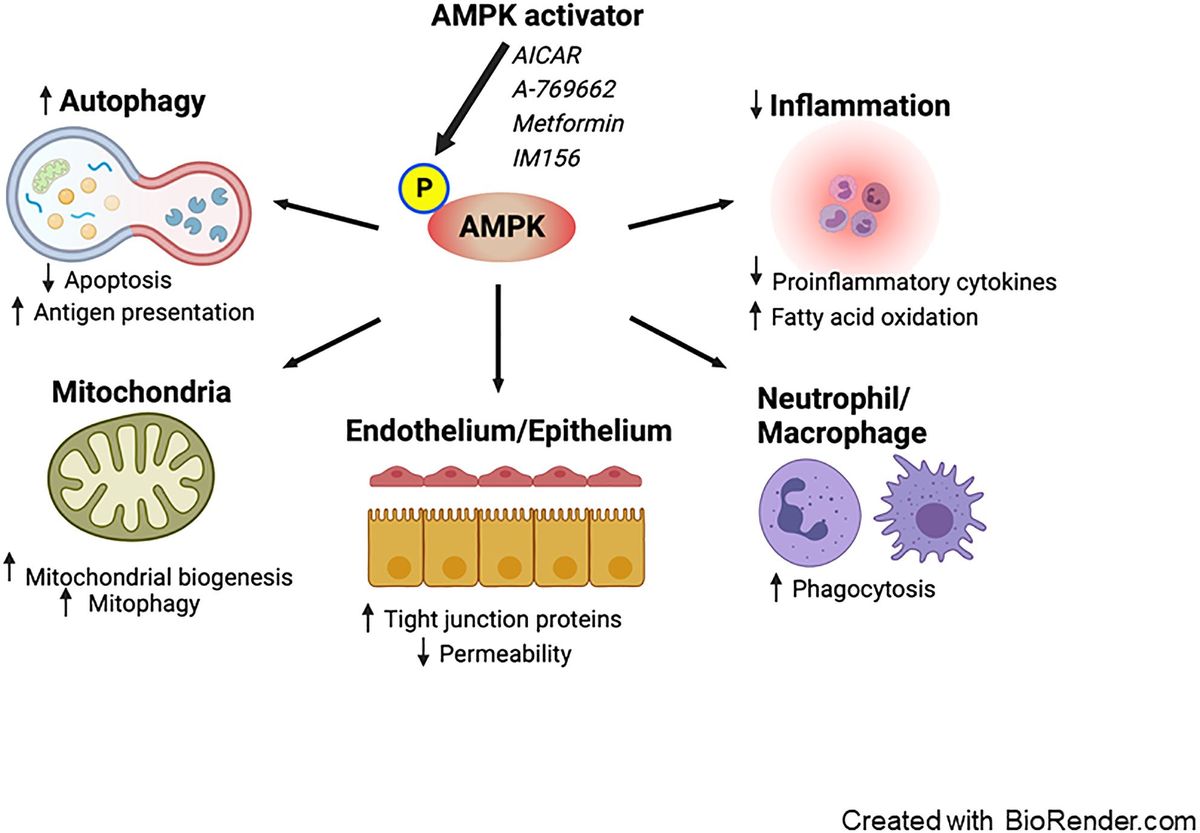
Potential beneficial effects of AMPK activation in treating sepsisExperimental studies (described in further detail below) have demonstrated that metformin can potentially mitigate sepsis-induced organ failure via AMPK activation (28) via a number of physiological benefits (Figure 1).
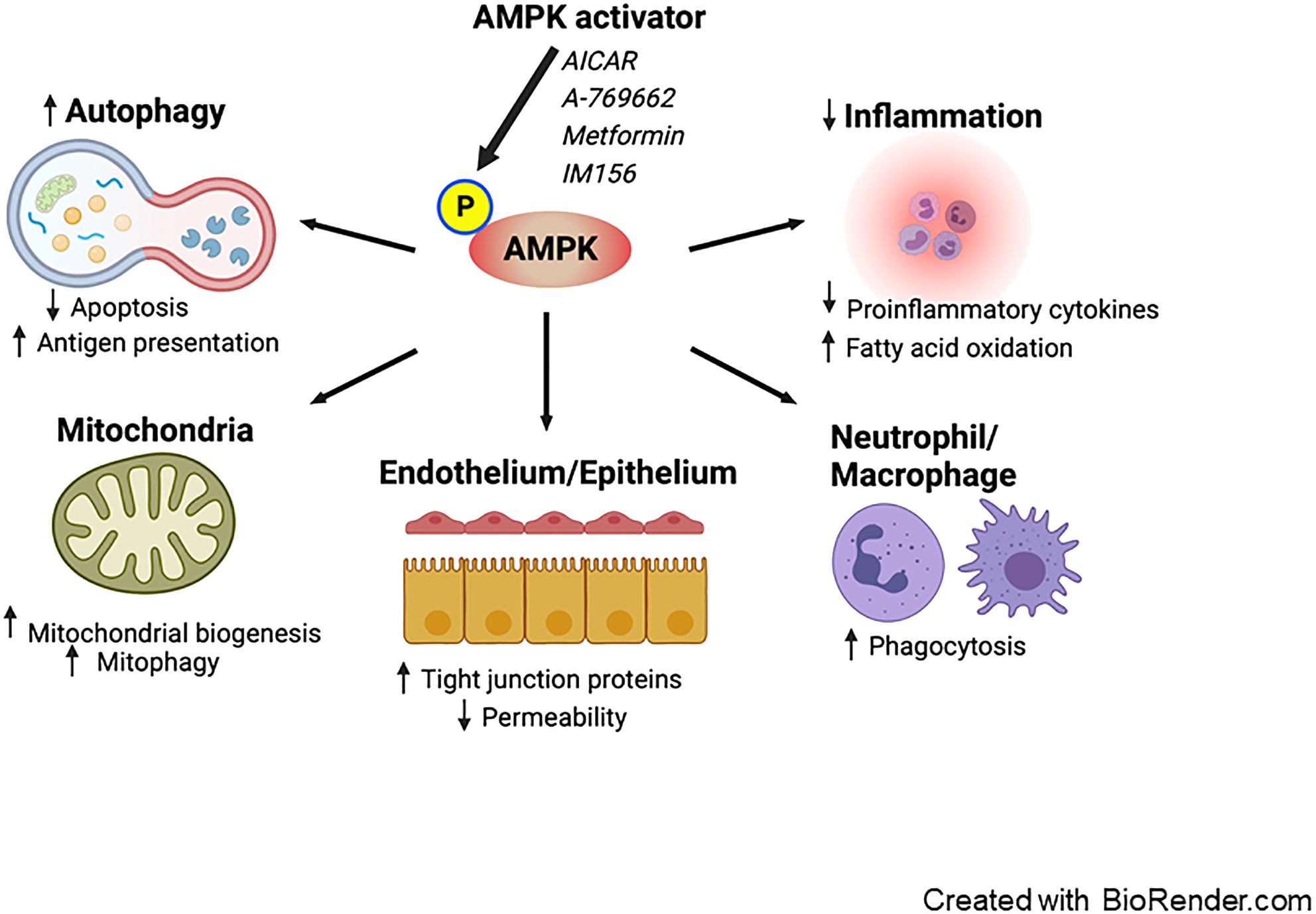
Figure 1. Proposed effects of AMPK activation on sepsis. Pharmacological activation of AMPK enhances autophagy, leading to a decrease in apoptosis and an increase in antigen presentation. AMPK activation promotes mitochondrial biogenesis and mitophagy. It exerts anti-inflammatory effects by reducing the production of pro-inflammatory cytokines and facilitating fatty acid oxidation. Furthermore, AMPK boosts the phagocytic activity of neutrophils and macrophages. Additionally, AMPK reduces vascular and intestinal permeability by restoring the integrity of tight junctions in the endothelium and epithelium. AMPK; AMP-activated protein kinase.
Anti-inflammatory effectSepsis is characterized as having dysregulated inflammation. While the balance between inflammation and immunosuppression is complex, modulating excessive inflammation – especially in the early stages of sepsis – and the resulting organ damage stands out as a potential therapeutic target (29). The role of AMPK in inflammation regulation has been extensively characterized. AMPK activation indirectly suppresses nuclear factor-κB, a pivotal transcriptional factor that induces the inflammatory response, through various mediators including SIRT1, FOXO, and PGC1α (30). Diminished AMPK activity in macrophages leads to a marked increase in the expression of pro-inflammatory cytokines IL-6 and TNF following LPS treatment with a simultaneous downregulation of the anti-inflammatory cytokine IL-10 (31). In contrast, AMPK activation causes a decrease in LPS-induced IL-6 and TNF production, coupled with a surge in IL-10 production. In addition, AMPK exhibits anti-inflammatory properties by modulating lipid metabolism. It is recognized that sepsis impacts fatty acid metabolism, with systemic inflammation and lipotoxicity stemming from reduced fatty acid oxidation and an accumulation of free fatty acids (32, 33). AMPK aids in enhancing fatty acid oxidation and mitigating excessive lipid accumulation via the phosphorylation of acetyl-CoA carboxylase (5).
Induction of autophagyAutophagy is a dynamic process through which intracellular materials are degraded and subsequently recycled in lysosomes. In mouse cecal ligation and puncture (CLP), a model of polymicrobial intraabdominal sepsis, autophagy is accelerated 6 hours after the induction of sepsis, but this is followed by a decline in autophagy in the liver, heart, and spleen (34).
Some preclinical studies have indicated that autophagy offers protective effects against sepsis in multiple vital organs including lung, heart, kidneys and brain although it may be potentially harmful in skeletal muscle (35). The beneficial aspects of autophagy can be attributed, in part, to the preservation of mitochondrial integrity, the prevention of apoptosis, and the enhancement of MHC II antigen presentation (36). Interestingly, suppressing autophagy in the liver heightens mortality rates after experimental sepsis, primarily due to the induction of apoptosis and mitochondrial damage (37). Thus, autophagy modulation might be a promising therapeutic target for sepsis treatment. The Unc-51-like autophagy-activating kinase 1 (ULK1) protein kinase is pivotal in the initiation of autophagy. AMPK acts directly as an upstream signal facilitating ULK1 activation. Moreover, AMPK suppresses the mechanistic target of rapamycin (mTOR), another main regulator that exerts negative control over autophagy (38).
Mitochondrial homeostasisMitochondria are responsible for numerous physiological processes, including ATP generation through oxidative phosphorylation, reactive oxygen species (ROS) production, calcium homeostasis, and the initiation of apoptosis. Mitochondrial homeostasis hinges on two opposing yet harmoniously coordinated processes: mitochondrial biogenesis and mitochondrial selective autophagy (often referred to as mitophagy) (39). Mitochondrial biogenesis is the process through which mitochondria adapt by increasing both in number and size, ensuring their robust heath and functionality. In contrast, mitophagy is a form of specialized autophagy targeting the selective degradation of damaged mitochondria via lysosomes. Beyond general autophagy, ULK1 is believed to be integral to mitophagy under various conditions (6).
Sepsis frequently leads to mitochondrial dysfunction, which has been mechanistically linked to poor patient outcomes (40, 41). This dysfunction is mainly marked by disturbance in electron transport chain function, causing excessive ROS production. The surge in ROS is further exacerbated by pro-inflammatory cytokines, such as IL-1, IL-6, and TNF. This cascade eventually triggers apoptosis, induced by the release of cytochrome c and other pro-apoptotic proteins (42, 43). Emerging evidence highlights the protective roles of both mitochondrial biogenesis and mitophagy during sepsis. In response to mitochondrial dysfunction, AMPK activation serves as a key compensatory mechanism by stimulating both mitochondrial biogenesis and mitophagy, restoring function and reducing damage (41, 44–46). Notably A-769662, a potent AMPK activator, bolsters mitochondrial biogenesis, as evidenced by an increase in nuclear PGC-1α activity within the lungs (47). Additionally, pre-treatment with lycorine, a benzyl isoquinoline alkaloid, decreases mortality and mitigates cardiac injury following CLP, by activating AMPK and suppressing ROS production and oxidative stress (48). In contrast, inhibiting mitophagy in macrophages leads to enhanced survival following sepsis. This phenomenon is attributed to ROS production in mitochondria, facilitating bacterial clearance and host defense (49).
Role of AMPK in immune cellsAMPK has been demonstrated to play a crucial role in the immune response, particularly within myeloid and T cells. Activating AMPK with either metformin or AICAR leads to reduced bacterial loads in peritoneal lavage following peritonitis-induced sepsis, associated with the amplification of neutrophil chemotaxis, phagocytosis, and bacterial neutralization (50). Additionally, metformin-induced AMPK activation inhibits the proliferation of Listeria pneumophilia within macrophages through enhanced mitochondrial ROS production (51). Moreover, AMPK is an important regulator of high mobility group box 1 (a consequential late mediator in lethal sepsis) release within stimulated macrophages and monocytes (52). AMPK is also crucial for T cell-mediated immunity as mice with a T cell-specific deletion of AMPKα1 have reduced counts of CD4+ and CD8+ T cells within bronchoalveolar lavage and lung tissues following pulmonary infections caused by either Influenza A or Listeria monocytogenes (53). While viral load in the lung remains consistent in mice with T cell-specific AMPKα1 deletion following Influenza A, knockout mice exhibit elevated bacterial loads within the liver (with a trend towards an increase in the spleen) following Listeria monocytogenes.
Antimicrobial effectAMPK plays a complex role in infectious diseases, especially viral infections, with its impact varying depending on the specific pathogen. AMPK activation also plays a key role in defending the host against Mycobacterium tuberculosis. This bacterium bypasses the host’s autophagic defense system in part by activating the mTOR pathway. AMPK therefore has been highlighted as a potential therapeutic target in the fight against mycobacterium infection (54). Within severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), AMPK activation leads to the phosphorylation and stabilization of angiotensin-converting enzyme 2 (ACE2) (55). While ACE2 facilitates SARS-CoV-2 entry into host cells, it also offers protection against lung injury by reducing inflammation, fibrosis, and pulmonary arterial hypertension (56). Metformin was extensively studied for its potential anti-COVID-19 effects. Based on lack of efficacy in the TOGETHER and COVID-OUT trials, metformin is not recommended for treatment of patients with COVID-19 in international guidelines; however, interest remains in identifying subsets of patients in whom it may be beneficial (57–60). IM156, a novel biguanide more potent than metformin in activating AMPK, offers protection following CLP by enhancing bacterial clearance, regulating cytokine release, and preventing lymphocyte apoptosis (61). IM156 also inhibits ROS generation and formation of neutrophil extracellular traps in response to LPS, so the mechanism underlying its survival benefit following sepsis is likely to be multifactorial.
Cytomegalovirus (CMV) reactivation is frequently seen in critically ill patients without primary immunodeficiency diseases (62). CMV relies on AMPK for glycolytic activation during its replication and suppressing AMPK activity can hinder CMV replication (63). Additionally, the effects of AMPK activation on bacterial infections vary based on the specific host cells infected and the nature of the pathogen. A comprehensive review of the impact of host AMPK activation on various microbial species has been conducted and can be found elsewhere (64).
Regulation of epithelial/endothelial tight junction assemblyAMPK activation exerts beneficial impacts on tight junction assembly. For instance, AMPK activation using metformin and AICAR has been shown to counteract airway epithelial barrier disruption caused by Pseudomonas aeruginosa, a significant microorganism linked to hospital-acquired infections in critically ill patients (65).
The intestinal epithelium serves as a dynamic protective barrier, separating the host from its external environment. This barrier’s integrity is regulated by both tight junction-dependent and independent mechanisms (66, 67). Gut barrier dysfunction characterized by intestinal hyperpermeability is associated with various diseases and systemic inflammation (68). AICAR bolsters ZO-1 formation, leading to a reduction in intestinal paracellular permeability following heat stress in rats (69). In a model of experimental colitis, metformin-induced AMPK activation upregulates the expression of occludin, ZO-1, and claudin-1, mitigating colonic damage (70). Current evidence suggests that sepsis impairs tight junctions in two different pathways (pore and leak), associated with intestinal hyperpermeability and unfavorable outcomes (67, 71). To our knowledge, there have been no studies investigating the impact of AMPK activation on intestinal barrier function in sepsis.
Disruption of endothelial tight junction proteins also worsens microvascular permeability and exacerbates multiple organ dysfunction (72). A number of investigations have centered on the interplay between AMPK activity and endothelial permeability in the context of LPS stimulation. AMPKα1-deficient mice display elevated cardiac vascular permeability, reduced endothelial ZO-1 expression, and myocardial edema post-LPS exposure (73). Conversely, in wild-type mice, these adverse effects are counteracted by AICAR. Additionally, both metformin and AICAR are effective in alleviating LPS-induced pulmonary endothelial permeability in rats (74).
AMPK activation as a therapeutic target in sepsisRegardless of the degree of AMPK activation in tissues following sepsis, a large number of pre-clinical studies have demonstrated beneficial effects of AMPK activation following either CLP-induced sepsis or sterile inflammation in LPS-induced endotoxemia. Table 1 summarizes the effects of AMPK activators or drugs/agents that can positively mediate AMPK pathway in animal studies (22, 23, 25, 47, 61, 74–104). Notably, a single dose of metformin administered 1 hour after CLP improves survival associated with preservation of metabolic fitness (25). Additionally, Dexmedetomidine, a sedative commonly used for critically ill patients, enhances hepatic autophagy through AMPK activation and leads to improved survival compared to vehicle in mice subjected to CLP (22).
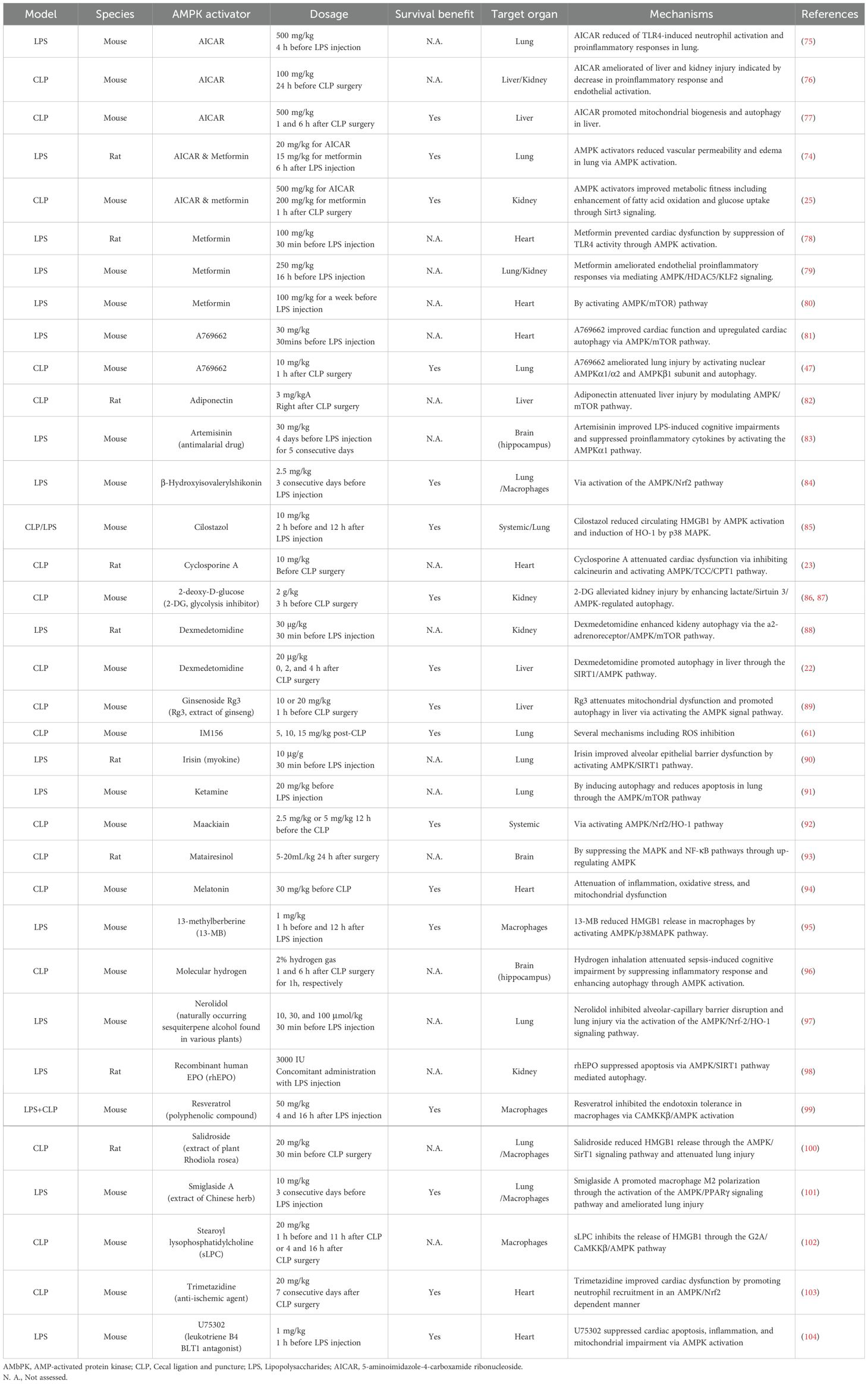
Table 1. Effects of AMPK activation on CLP-induced sepsis or LPS-induced inflammation.
Drawbacks of AMPK activationWhile a substantial body of evidence highlights the beneficial effects of AMPK activation, data also suggest that inhibiting AMPK can reduce liver injury. The AMPK inhibitor compound C reduces hepatocyte apoptosis and mitigates liver damage by inhibiting the pro-apoptotic protein JNK in a model of LPS/D-galactosamine-induced fulminant hepatitis (105). Additionally, inhibiting AMPK improves LPS-induced liver damage by suppressing the ROS/NF-κB signaling pathway (106). The role of AMPK in myocardial ischemic injury remains debated, with conflicting views on whether it is beneficial or harmful (107). Furthermore, in certain tumor types or under conditions of limited nutrients or hypoxia, AMPK has been observed to promote tumor growth (13).
DiscussionAlthough AMPK activation shows promise in the treatment of sepsis, several barriers remain prior to translation to bedside use above and beyond the absence of clinical trials supporting the efficacy of this approach. First, sepsis is a highly heterogeneous condition characterized by the presence of multiple suphenotypes, each with distinct pathophysiological mechanisms and clinical outcomes (108–116). Additionally, subtype strategies using clinical, biomarker and transcriptomic data do not identify comparable patient populations with sepsis (117), suggesting precision medicine approaches to identifying which patients might respond to AMPK-targeted therapies would be challenging. Next, the diversity of cell types involved in the septic response further complicates the potential usage of AMPK activation strategies due to potential of unwanted off-target effects. Additionally, immunometabolic paralysis – a state of metabolic dysfunction in immune cells – is another potential barrier to effective sepsis treatment with AMPK activation. While AMPK is a central regulator of cellular metabolism, its role in reversing immunometabolic paralysis in sepsis is not yet fully understood. Emerging evidence suggests that AMPK activation may influence immune cell function, but the effects appear to be context-dependent, requiring more precise targeting strategies. Future research should focus on elucidating the specific pathways through which AMPK modulates immune metabolism during sepsis and developing biomarkers to guide the use of AMPK-targeted therapies in overcoming immunometabolic paralysis (44).
Recent studies have identified itaconate, a metabolite derived from the TCA cycle, as a significant modulator of immune responses, contributing to disease tolerance in sepsis (118). Itaconate is mainly produced by activated macrophages through the enzyme immune-responsive gene 1 (IRG1) and inhibits ferroptosis of macrophages via Nrf2 pathways in sepsis-induced acute lung injury (119). As a central regulator of cellular energy homeostasis, AMPK interacts with multiple metabolic pathways, including those linked to the TCA cycle. This interaction suggests that AMPK and itaconate may work together to modulate immune responses and metabolic reprogramming in sepsis given the role of mitochondrial TCA cycle metabolites in physiology and disease (120). Additionally, AMPK may influence itaconate production by regulating IRG1 expression, as AMPK can modulate inflammatory responses through altering macrophage polarization (121). Given that itaconate has been demonstrated to regulate AMPK signaling in hepatocytes (122, 123), this suggests a complementary approach to direct AMPK activation in modulating metabolic and inflammatory responses in sepsis.
Ultimately, AMPK plays a crucial role in numerous biological processes that are relevant to sepsis pathogenesis, and numerous studies suggest that either AMPK activation or deficiency can influence susceptibility to sepsis in ways that vary by age or sex (124, 125). It is exciting to consider the possibility of AMPK as a potential therapeutic target in human sepsis given the numerous pre-clinical studies demonstrating a benefit of AMPK activation and associative studies suggesting a potential benefit of metformin in critical illness. However, there is currently insufficient human evidence to support using metformin or other AMPK activators to treat septic patients in the ICU. Ongoing and future clinical trials may clarify the role (if any) in activating AMPK in human sepsis while future research focused on tissue-specific and condition-specific AMPK activators will hopefully clarify mechanisms underlying potential efficacy.
Author contributionsTY: Writing – original draft, Writing – review & editing. CC: Writing – original draft, Writing – review & editing.
FundingThe author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was supported by the National Institutes of Health (GM148217).
Conflict of interestThe authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statementThe authors declare that no Generative AI was used in the creation of this manuscript.
Publisher’s noteAll claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References1. Rudd KE, Johnson SC, Agesa KM, Shackelford KA, Tsoi D, Kievlan DR, et al. Global, regional, and national sepsis incidence and mortality, 1990-2017: analysis for the Global Burden of Disease Study. Lancet. (2020) 395:200–11. doi: 10.1016/S0140-6736(19)32989-7
PubMed Abstract | Crossref Full Text | Google Scholar
2. Evans L, Rhodes A, Alhazzani W, Antonelli M, Coopersmith CM, French C, et al. Surviving sepsis campaign: international guidelines for management of sepsis and septic shock 2021. Crit Care Med. (2021) 49:e1063–e143. doi: 10.1097CCM.0000000000005337
PubMed Abstract | Google Scholar
3. Singer M, Deutschman CS, Seymour CW, Shankar-Hari M, Annane D, Bauer M, et al. The third international consensus definitions for sepsis and septic shock (Sepsis-3). JAMA. (2016) 315:801–10. doi: 10.1001/jama.2016.0287
PubMed Abstract | Crossref Full Text | Google Scholar
4. De Backer D, Deutschman CS, Hellman J, Myatra SN, Ostermann M, Prescott HC, et al. Surviving sepsis campaign research priorities 2023. Crit Care Med. (2024) 52:268–96. doi: 10.1097/CCM.0000000000006135
PubMed Abstract | Crossref Full Text | Google Scholar
5. Steinberg GR, Carling D. AMP-activated protein kinase: the current landscape for drug development. Nat Rev Drug Discovery. (2019) 18:527–51. doi: 10.1038/s41573-019-0019-2
Crossref Full Text | Google Scholar
7. Sharma A, Anand SK, Singh N, Dwivedi UN, Kakkar P. AMP-activated protein kinase: An energy sensor and survival mechanism in the reinstatement of metabolic homeostasis. Exp Cell Res. (2023) 428:113614. doi: 10.1016/j.yexcr.2023.113614
PubMed Abstract | Crossref Full Text | Google Scholar
8. Liang H, Ding X, Li L, Wang T, Kan Q, Wang L, et al. Association of preadmission metformin use and mortality in patients with sepsis and diabetes mellitus: a systematic review and meta-analysis of cohort studies. Crit Care. (2019) 23:50. doi: 10.1186/s13054-019-2346-4
PubMed Abstract | Crossref Full Text | Google Scholar
9. Li Y, Zhao H, Guo Y, Duan Y, Guo Y, Ding X. Association of preadmission metformin use and prognosis in patients with sepsis and diabetes mellitus: A systematic review and meta-analysis. Front Endocrinol (Lausanne). (2021) 12:811776. doi: 10.3389/fendo.2021.811776
PubMed Abstract | Crossref Full Text | Google Scholar
10. Gómez H, Del Rio-Pertuz G, Priyanka P, Manrique-Caballero CL, Chang CH, Wang S, et al. Association of metformin use during hospitalization and mortality in critically ill adults with type 2 diabetes mellitus and sepsis. Crit Care Med. (2022) 50:935–44. doi: 10.1097/CCM.0000000000005468
PubMed Abstract | Crossref Full Text | Google Scholar
11. Lee JY, Kim ES, Chang E, Bae S, Jung J, Kim MJ, et al. Clinical impact of metformin exposure during Staphylococcus aureus bacteremia in patients with diabetes mellitus. Eur J Clin Microbiol Infect Dis. (2023) 42:1439–47. doi: 10.1007/s10096-023-04679-6
PubMed Abstract | Crossref Full Text | Google Scholar
12. Saraiva IE, Hamahata N, Huang DT, Kane-Gill SL, Rivosecchi RM, Shiva S, et al. Metformin for sepsis-associated AKI: a protocol for the Randomized Clinical Trial of the Safety and FeasibiLity of Metformin as a Treatment for sepsis-associated AKI (LiMiT AKI). BMJ Open. (2024) 14:e081120. doi: 10.1136/bmjopen-2023-081120
PubMed Abstract | Crossref Full Text | Google Scholar
16. Yang Y, Dong R, Hu D, Chen Z, Fu M, Wang DW, et al. Liver kinase B1/AMP-activated protein kinase pathway activation attenuated the progression of endotoxemia in the diabetic mice. Cell Physiol Biochem. (2017) 42:761–79. doi: 10.1159/000478068
PubMed Abstract | Crossref Full Text | Google Scholar
17. Otto NA, de Vos AF, van Heijst JWJ, Roelofs J, van der Poll T. Association of myeloid liver kinase B1 depletion with a reduction in alveolar macrophage numbers and an impaired host defense during gram-negative pneumonia. J Infect diseases. (2022) 225:1284–95. doi: 10.1093/infdis/jiaa416
Crossref Full Text | Google Scholar
18. Li C, Zhang C, Zhou H, Feng Y, Tang F, Hoi MPM, et al. Inhibitory effects of betulinic acid on LPS-induced neuroinflammation involve M2 microglial polarization via caMKKβ-dependent AMPK activation. Front Mol Neurosci. (2018) 11:98. doi: 10.3389/fnmol.2018.00098
PubMed Abstract | Crossref Full Text | Google Scholar
19. Yao Y, Ji Y, Ren J, Liu H, Khanna R, Sun L. Inhibition of autophagy by CRMP2-derived peptide ST2-104 (R9-CBD3) via a CaMKKβ/AMPK/mTOR pathway contributes to ischemic postconditioning-induced neuroprotection against cerebral ischemia-reperfusion injury. Mol Brain. (2021) 14:123. doi: 10.1186/s13041-021-00836-0
PubMed Abstract | Crossref Full Text | Google Scholar
20. Bernard GR, Wheeler AP, Russell JA, Schein R, Summer WR, Steinberg KP, et al. The effects of ibuprofen on the physiology and survival of patients with sepsis. The Ibuprofen in Sepsis Study Group. N Engl J Med. (1997) 336:912–8. doi: 10.1056/NEJM199703273361303
PubMed Abstract | Crossref Full Text | Google Scholar
21. Heidary Moghaddam R, Samimi Z, Asgary S, Mohammadi P, Hozeifi S, Hoseinzadeh-Chahkandak F, et al. Natural AMPK activators in cardiovascular disease prevention. Front Pharmacol. (2021) 12:738420. doi: 10.3389/fphar.2021.738420
PubMed Abstract | Crossref Full Text | Google Scholar
22. Yu Q, Zou L, Yuan X, Fang F, Xu F. Dexmedetomidine protects against septic liver injury by enhancing autophagy through activation of the AMPK/SIRT1 signaling pathway. Front Pharmacol. (2021) 12:658677. doi: 10.3389/fphar.2021.658677
PubMed Abstract | Crossref Full Text | Google Scholar
23. Liu J, Chen D, Liu X, Liu Z. Cyclosporine A attenuates cardiac dysfunction induced by sepsis via inhibiting calcineurin and activating AMPK signaling. Mol Med Rep. (2017) 15:3739–46. doi: 10.3892/mmr.2017.6421
PubMed Abstract | Crossref Full Text | Google Scholar
24. Liu Z, Bone N, Jiang S, Park DW, Tadie JM, Deshane J, et al. AMP-activated protein kinase and glycogen synthase kinase 3β Modulate the severity of sepsis-induced lung injury. Mol Med (Cambridge Mass). (2016) 21:937–50. doi: 10.2119/molmed.2015.00198
Crossref Full Text | Google Scholar
25. Jin K, Ma Y, Manrique-Caballero CL, Li H, Emlet DR, Li S, et al. Activation of AMP-activated protein kinase during sepsis/inflammation improves survival by preserving cellular metabolic fitness. FASEB J. (2020) 34:7036–57. doi: 10.1096/fj.201901900R
PubMed Abstract | Crossref Full Text | Google Scholar
26. Stana F, Vujovic M, Mayaki D, Leduc-Gaudet JP, Leblanc P, Huck L, et al. Differential regulation of the autophagy and proteasome pathways in skeletal muscles in sepsis. Crit Care Med. (2017) 45:e971–e9. doi: 10.1097/CCM.0000000000002520
PubMed Abstract | Crossref Full Text | Google Scholar
27. Weber-Carstens S, Schneider J, Wollersheim T, Assmann A, Bierbrauer J, Marg A, et al. Critical illness myopathy and GLUT4: significance of insulin and muscle contraction. Am J Respir Crit Care Med. (2013) 187:387–96. doi: 10.1164/rccm.201209-1649OC
PubMed Abstract | Crossref Full Text | Google Scholar
28. Ismail Hassan F, Didari T, Khan F, Niaz K, Mojtahedzadeh M, Abdollahi M. A review on the protective effects of metformin in sepsis-induced organ failure. Cell J. (2020) 21:363–70. doi: 10.22074/cellj.2020.6286
PubMed Abstract | Crossref Full Text | Google Scholar
31. Sag D, Carling D, Stout RD, Suttles J. Adenosine 5'-monophosphate-activated protein kinase promotes macrophage polarization to an anti-inflammatory functional phenotype. J Immunol. (2008) 181:8633–41. doi: 10.4049/jimmunol.181.12.8633
PubMed Abstract | Crossref Full Text | Google Scholar
32. Idrovo JP, Yang WL, Jacob A, Corbo L, Nicastro J, Coppa GF, et al. Inhibition of lipogenesis reduces inflammation and organ injury in sepsis. J Surg Res. (2016) 200:242–9. doi: 10.1016/j.jss.2015.06.059
PubMed Abstract | Crossref Full Text | Google Scholar
33. Van Wyngene L, Vandewalle J, Libert C. Reprogramming of basic metabolic pathways in microbial sepsis: therapeutic targets at last? EMBO. (2018) 10:e8712. doi: 10.15252/emmm.201708712
Crossref Full Text | Google Scholar
34. Takahashi W, Watanabe E, Fujimura L, Watanabe-Takano H, Yoshidome H, Swanson PE, et al. Kinetics and protective role of autophagy in a mouse cecal ligation and puncture-induced sepsis. Crit Care. (2013) 17:R160. doi: 10.1186/cc12839
PubMed Abstract | Crossref Full Text | Google Scholar
37. Oami T, Watanabe E, Hatano M, Teratake Y, Fujimura L, Sakamoto A, et al. Blocking liver autophagy accelerates apoptosis and mitochondrial injury in hepatocytes and reduces time to mortality in a murine sepsis model. Shock. (2018) 50:427–34. doi: 10.1097/SHK.0000000000001040
PubMed Abstract | Crossref Full Text | Google Scholar
40. Kumar S, Srivastava VK, Kaushik S, Saxena J, Jyoti A. Free radicals, mitochondrial dysfunction and sepsis-induced organ dysfunction: A mechanistic insight. Curr Pharm Des. (2024) 30:161–8. doi: 10.2174/0113816128279655231228055842
PubMed Abstract | Crossref Full Text | Google Scholar
41. Lira Chavez FM, Gartzke LP, van Beuningen FE, Wink SE, Henning RH, Krenning G, et al. Restoring the infected powerhouse: Mitochondrial quality control in sepsis. Redox Biol. (2023) 68:102968. doi: 10.1016/j.redox.2023.102968
PubMed Abstract | Crossref Full Text | Google Scholar
43. Preau S, Vodovar D, Jung B, Lancel S, Zafrani L, Flatres A, et al. Energetic dysfunction in sepsis: a narrative review. Ann Intensive Care. (2021) 11:104. doi: 10.1186/s13613-021-00893-7
PubMed Abstract | Crossref Full Text | Google Scholar
44. McBride MA, Owen AM, Stothers CL, Hernandez A, Luan L, Burelbach KR, et al. The metabolic basis of immune dysfunction following sepsis and trauma. Front Immunol. (2020) 11:1043. doi: 10.3389/fimmu.2020.01043
PubMed Abstract | Crossref Full Text | Google Scholar
45. Zhu CL, Yao RQ, Li LX, Li P, Xie J, Wang JF, et al. Mechanism of mitophagy and its role in sepsis induced organ dysfunction: A review. Front Cell Dev Biol. (2021) 9:664896. doi: 10.3389/fcell.2021.664896
PubMed Abstract | Crossref Full Text | Google Scholar
47. Kitzmiller L, Ledford JR, Hake PW, O'Connor M, Piraino G, Zingarelli B. Activation of AMP-activated protein kinase by A769662 ameliorates sepsis-induced acute lung injury in adult mice. Shock. (2019) 52:540–9. doi: 10.1097/SHK.0000000000001303
留言 (0)