
記住我

In the clinical setting, a typical case would usually present with a long duration of Diabetes Mellitus (DM), uncontrolled blood glucose levels, hypertension, dyslipidemia, and obesity predisposing the individuals to diabetic vascular complications (DVC). It is worth noting that hypertension is a vital risk factor for DVC as it is characterized by vascular dysfunction and injury [1]. Additionally, chronic hyperglycemia and insulin resistance play an important role in the initiation of vascular complications of DM and involve a number of mechanisms (Fig. 1). However, these clinical characteristics cannot be used solely to predict the risk of DVC. Emerging evidence suggests MicroRNAs (miRNAs) may play a role in the vasculopathy of DM [2].
Fig. 1
Illustrating the interlinked relationship and overlap of Major Diabetic Vascular Complications (Diabetic Cardiovascular Disease; DCVD, Diabetic Kidney Disease; DKD, Diabetic Retinopathy; DR). Alongside common shared risk factors, shared genetic may contribute to long term diabetic patient’s susceptibility to Micro and Macro vascular complications GFR; glomerular filtration rate
The current treatments for DVC complications do not work to reverse the disease process; instead, they focus almost entirely on preventing or managing problems that have already developed. Studies have shown an apparent decline in both the onset and progression of DVC through intensive glucose-lowering treatments and further reduce the risk of complications by controlling blood pressure with antihypertensive medications (angiotensin-converting enzyme inhibitors and angiotensin II receptor blockers) [3]. These medications inhibit the renin–angiotensin–aldosterone system (RAAS) and reduce the risk of complications [3]. Recently, a new class of anti-hyperglycemic drugs, such as sodium-glucose cotransporter 2 (SGLT2) inhibitors and glucagon-like peptide-1 (GLP-1) receptor agonists, have demonstrated significant renal and cardiovascular benefits in patients with T2DM [4]. This is supposedly due to both glucose-dependent and glucose-independent mechanisms [4]. It is worth noting that mutations in the Solute Carrier Family 5 Member 2 (SLC5A2) gene that encodes SGLT2, the major glucose co-transporter in the kidney's proximal tubule, known to cause familial renal glycosuria [5]. This condition is characterized by decreased renal glucose reabsorption and increased glucose excretion. This paradigm demonstrates that investigating the genetics of disorders associated with glucose dysregulation and the relationship between genetics and therapy may give insight into the clinical effectiveness of innovative treatments. Interestingly, over the last few decades, research in the field of genetic factors in DM has gained much interest and has clearly shown their contribution to the development and its progression.
Genetic explorationThere have been various methods by which the genetic risk for this complex disease has been studied. So far, multiple studies using Genetic Linkage Analysis (GLA) and Alpha Glucosidase (GAA) have shown to cause variants or genes, showing evidence for genetic susceptibility. While GAA is based on the common variant hypothesis [6], GLA was performed using the rare variant hypothesis to identify susceptible genes [7]. However, GLA had its limitations and drawbacks. And more recently, Genetic wide association studies (GWAS) and epigenetics have been used to identify the susceptible variants or genes.
In this literature review, the focus is on the latest advances in genetic (GWAS) and epigenetic (miRNA) studies in T2DM. Moreover, it summarizes data from prior genetic research methodologies on susceptibility genetic variants and epigenetic modifications that influence dorsal vagal complex (DVC). Finally, Understanding the etiological mechanisms underlying DVC can greatly help identify genetic variants, structural variants, and epigenetic changes that either contribute to the development of or protect against DVC, which in turn has important implications for the development of personalized medicine for DVC and potential biomarkers.
Genetic background of T2DM complicationsSeveral important organ systems, such as the eyes, kidneys, and cardiovascular system, are affected by either DVC, which are typically categorized as microvascular complications, such as diabetic nephropathy (DN) and diabetic retinopathy (DR), or macrovascular complications, such as diabetic cardiovascular complications (DCC) [8,9,10,11]. There is currently evidence that genetic factors have a role in DVC, namely DN, DR, and DCC [12]. This complicated diseases’ genetic risk has been investigated using a variety of approaches.
Genetic linkage analysis (GLA) identifies the chromosomal location of disease genes because genes that are physically nearby on a chromosome remain connected during meiosis [13].
For candidate gene analysis, based on their physiological roles, candidate genes with a known sequence and location that may be implicated in the pathogenesis of the disease are identified. In contrast, Genome-wide screens are a more effective method that may be used to screen the completely human genome for gene linkage or association with a disease without assuming anything about disease pathophysiology. This type of approach has been used successfully to identify susceptibility genetic loci for DVC. Genetic linkage study typically includes the following steps: discovering linked loci, verifying linked loci, precise mapping of verified loci, and assessing genes in the linked area through functional tests [14].
GWAS are more sensitive than genetic linkage studies and can identify minor susceptibility genes [15]. Single nucleotide polymorphisms (SNPs) are the most significant genetic variation for genetic association study due to the high density of SNPs throughout the whole human genome [15]. This study showed that a newer method that uses extensive evaluation of SNP in big datasets is GWAS. Type 2 DM (T2DM) is a complex metabolic disease that occurs as a result of insulin resistance and low insulin production. T2DM involves many organ systems that include macro-vascular and micro-vascular complications [16]. Several GWAS and candidate gene studies have suggested a large number of SNPs on several genes such as Hematopoietically expressed homeobox protein (HHEX) that were associated with T2DM susceptibility [16].
Gowing number of studies have demonstrated the association between gut microbiome and T2DM microvascular complications, however the causal relationship remains unclear [17]. Recently, using Mendelian Randomization (MR) methods, it was demonstrated the causal relationship between gut microbiome and microvascular complications in T2DM, providing a new strategy for the prevention and treatment of T2DM [17].
Candidate gene association analysis uses candidate genes of a known sequence and location that are involved in the disease pathology. However, approaches based on prior hypotheses have limited power to detect novel genetic variants. Instead, a non-prior hypothesis is a more powerful approach for identifying gene association with a disease by screening the whole human genome.
Epigenetics is the study of changes in gene expression that are not caused by changes in the DNA sequence. Things like DNA methylation, histone modification, and microRNAs can cause these changes. Epigenetics help explain how cells with the same DNA can change into different cell types with different phenotypes [18]. miRNAs are noncoding RNAs with 22 nucleotides. When they bind to the 3′-untranslated region of target mRNAs, they can either stop the mRNA from being made or break it down [19]. Importantly, miRNAs play important roles in how tissues respond to environmental signals without changing the DNA sequence. They do this by regulating genes quickly and in a way that can be modified. miRNAs may also be controlled by epigenetics, as histone modifications and changes in chromatin structure can affect miRNA transcription and expression [19]. Recent research indicates that microRNAs have a crucial role in a variety of disorders. miRNA expression is changed in diabetic cardiac and skeletal muscle, liver, kidney, and endothelium, all of which are tissues negatively impacted by DM. It has been demonstrated that microRNAs can affect genes in numerous biological processes, such as the secretion of insulin, lipid production, fat metabolism, and adipogenesis, which are essential pathways in the etiology of DM [18].
Microvascular: diabetic kidney diseaseDiabetic Kidney Disease (DKD), more commonly known as DN, is a progressive renal disease associated with long-term DM, which can progress to end-stage renal disease, requiring dialysis or kidney transplantation [20]. Patients typically present in association with long-standing DM, DR, albuminuria, and a progressive decline of estimated Glomerular Filtration Rate (eGFR) [23]. In addition, the presence of chronic hyperglycemia is a risk factor for both micro and macrovascular complications, including DKD, Diabetic Cardiovascular Disease (DCVD), DR, and Diabetic Foot Ulcers (DFU), increasing the mortality rate of T2DM patients [23].
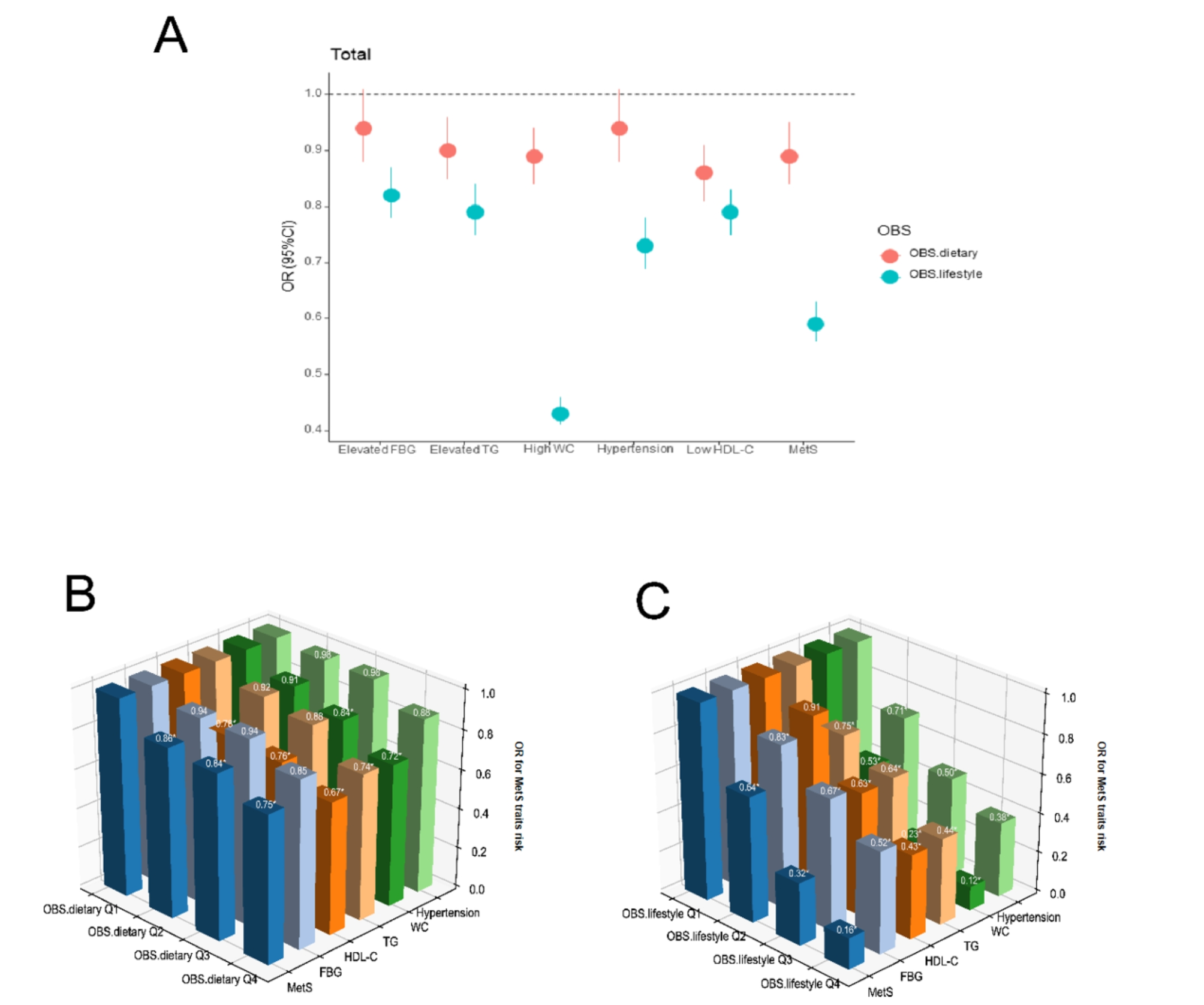
At present, Urinary Albumin Creatinine Ratio (UACR) and eGFR are the only key markers used to identify and assess DKD [24]. The Kidney Disease Improving Global Outcomes (KDIGO) 2012 guidelines have staged Chronic Kidney Disease (CKD) based on these two critical markers which help in predicting the prognosis of the disease (Fig. 2) [25].
Fig. 2
eGFR categories based on the clinical guidelines of KDIGO. Green: low risk (if no other markers of kidney disease, no CKD); Yellow: moderately increased risk; Orange: high risk; Red: very high risk. GFR; glomerular filtration rate
The pathophysiology of DKD a significant microvascular consequence of T2DM is complicated and heterogeneous in nature [26]. Hemodynamic alterations, such as increased intra glomerular pressure and hyper filtration, have already been recognized as effective mechanisms in the initiation and development of DKD [26]. However, the detailed pathogenesis of the disease remains to be understood. Evidence suggests that inherited factors and acquired elements accumulation of Advanced Glycation End products (AGEs) known as “metabolic memory” have been crucial in developing T2DM-induced DKD in recent years [27].In addition, many non-modifiable risk factors for DKD, including ethnicity and genetics, have been documented [27]. Hyperglycemia, hyperlipidemia, and hypertension are some risk factors contributing to the pathologic changes in DKD. Therefore, the current management strategy for DKD includes a multidisciplinary approach including glucose, lipid, and BP management with the help of anti-hyperglycemic, statins, angiotensin-converting enzyme inhibitors (ACEI), and angiotensin receptor blockers (ARBs), respectively. In 2019, SGLT2 inhibitors were added as a new drug of choice for treating DKD [28]. The Canagliflozin and Renal Events in DM with Established Nephropathy Clinical Evaluation (CREDENCE) study showed that SGLT2 inhibitors stopped DKD from progressing [28]. However, despite these measures, many people with DKD still progress to end-stage renal disease [27]. Therefore, novel molecular pathways causing DKD should be studied to identify potential biomarkers for the progression of the disease. The importance of epigenetic processes, particularly miRNAs, has been investigated in this review with the development of technology.
HeritanceMultiple heritability studies conducted earlier have proved a vital link in the family-based studies in DKD. Interestingly, those with DKD-affected siblings had roughly 2–4 times the probability of getting DKD compared to those with DM-affected siblings but without DKD. In T2DM, people estimate single nucleotide polymorphisms (SNPs) heritability to be 8%, perhaps because of the significant phenotypic heterogeneity of kidney disease in T2DM.
Linkage studiesFunctional polymorphisms affecting the activity of candidate pathways, such as the nitric oxide, renin-angiotensin, and bradykinin systems, have been investigated in candidate gene linkage studies for DKD. In addition, other pathways associated with glucose homeostasis, lipid synthesis, and insulin resistance were investigated since they may be involved in multiple disease processes through shared mechanisms. Despite this, there has been no consistent and reproducible discovery of genetic loci or candidate genes for DKD risk or protection; however, this may be due to several factors, such as a small sample size or considerable genetic and phenotypic heterogeneity.
In 1998, the first linkage studies for DKD were performed on Pima Indians, showing a strong association to chromosome 3q24 [29]. Later, in the early 2000s, further studies were conducted in Turkish populations showing a strong association of chromosome 18q to DKD in T2DM with an odds score of 6.1[30]. Furthermore, a genome-wide search for DKD in African American families showed evidence for nephropathy loci on for susceptibility loci on chromosomes 3q, 7p, and 18q [31]. A brief list of detected chromosome regions for DKD is illustrated in (Table 1).
Table 1 Linkage analysis of DKDCandidate gene association studiesThe gene to be examined is chosen based on an understanding of its contribution towards DKD pathophysiology, such as those involving blood pressure management, proteinuria severity, insulin resistance, lipid metabolism, or other pathways implicated in the progression of DKD. In addition, the current review highlights the most recent data on genetic variations (related to RAS, glucose and lipid metabolism, and some cytoskeleton proteins) that confirm the importance of genetic factors in diabetic nephropathy. Over the years, Comprehensive Geriatric Assessment (CGA) studies have identified more than150 important genes that have shown their association with DKD, for example, angiotensin I converting enzyme (ACE), carnosine dipeptidase 1 (CNDP1), fatty acid binding protein (FABP2), Ectonucleotide Pyrophosphatase/Phosphodiesterase(ENPP1) and Glucose Transporter 1 (GLUT1).
ACE helps convert Angiotensin I to Angiotensin II by the potent vasoconstrictor effect. It also deactivates the bradykinin vasodilatory effect by causing proteolysis. In Brazilian T2DM patients, a connection between the Insertion/Deletion (I/D) polymorphism in ACE and the development of DKD has been described [33]. The role of the ACE gene on chromosome 17q23 in DKD has been investigated multiple times and shown an association in a meta-analysis conducted on relevant studies conducted between 1994 and 2004, especially in the Asian population [34]. Clearly, polymorphisms in this gene correlate with circulating ACE levels. Additionally, ACE inhibitors are regularly used as antihypertensive medicines in treating DKD, and the ACE I/D polymorphism is arguably the most well researched candidate gene in DN.
The CNDP1 (carnosine dipeptidase 1) gene is located in chromosome 18q22.3, encoding serum carnosinase (CN-1), found to confer the susceptibility for DKD and end-stage renal disease (ESRD) in T2DM [35]. In a study conducted illustrated that the serum CN-1 concentrations were considerably lower in T2DM patients with poor or moderate renal function (eGFR 60 ml/min/1.73m2) comparing those with adequate renal function (eGFR 60 ml/min/1.73m2), but higher in the latter group compared to healthy individuals [36]. This is consistent with previous observations that a high serum CN-1 concentration increases the likelihood of developing DKD [37]. Serum CN-1 concentrations decline as DKD advances, probably as a result of urine excretion and protein loss [38]. Additionally, the study showed the presence of urinary CN-1 in individuals with low eGFR provides additional evidence for the previously stated notion [36]. Additionally, polymorphisms in protein ezrin, radixin, moesin (FERM) domain containing 3 FRMD3 gene is located in chromosome 9q21.32, are associated with DKD in T2D [37, 38]. Fatty Acid Binding Protein 2 (FABP2) is also a potential gene for DKD susceptibility since it has been associated with albuminuria in patients with T2DM [40, 41].
In addition, ENPP1 polymorphism was associated with integrated reporting (IR) in a meta-analysis conducted in patients with T2DM detected a significant association between the ENPP1K121Q polymorphism and increased susceptibility of DKD in European and Asian populations [42]. In other studies, Glucose transporter type 1 (GLUT1) polymorphisms were investigated as a risk factor for DKD as its association with early kidney changes as it works as a glucose transporter in kidneys, which is pivotal in raising intracellular glucose levels by activating pathogenic pathways. GLUT1 polymorphisms related to DKD were also studied in genomic analysis [43]. A data from GAA in DKD using CGA approach in (Table 2).
Table 2 Data from GAA in DKD using CGA approachGenome-wide association studies and single nucleotide polymorphismsDN is a condition shared by people with both T1DM and T2DM. However, it depicts a diverse collection of disorders that are perpetuated by different processes and could even coexist in various combinations, especially in people with T2DM. It has been shown that the prevalence of DKD is higher in T2DM, with a rapid decline in renal function that could be attributed to concurrent risk factors like hypertension and obesity [44]. In addition, the phenotypic variations in T2DM patients need to be better understood, lending credence to the genetic contribution. Thus, the genetic discoveries in T2DM are challenged by the fact that the prevalence of DKD varies across ethnic populations. It is worth noting that a decline in kidney function is measured by the decline in the eGFR, while glomerular filtration barrier dysfunction, is measured by albuminuria, can occur independently. This indicates that the two fundamental characteristics of DKD are caused by various mechanisms and may contribute to the genetic effects.
Genome-wide association studies and DKD - important discoveriesIn 2005, the first GWAS revealed engulfment and motility protein 1 (ELMO1) as a gene conferring susceptibility for DKD in T2DM in Japanese patients [45]. The ELMO1 locus on chromosome p14.1 encodes a member of the engulfment and cell motility protein family that is hardly detectable in podocytes and tubular epithelial cells in healthy kidneys but significantly elevated in diabetic kidneys and CV-1 (simian) in Origin, and carrying the SV40 genetic material (COS) cells exposed to high glucose. Furthermore, ELMO1 enhances the expression of extracellular matrix protein extracellular matrix genes (COL1A1, MMP2 and FN1) in COS cells in a TGF-β-independent manner, which leads to the accumulation of ECM, thickening of renal tubules and glomerular basement membrane, thus increasing the risk of DN [45].
Further discovery of 6 polymorphism sites (rs741301 rs1345365, rs11769038, rs10951509, rs1882080, rs6462776, rs6462777) of ELMO1 gene showed association to DKD in 200 Chinese subjects (123 T2DM with DN case subjects and 77 T2DM without DN control subjects) [46].
In one of the largest combined GWAS data from four studies of European descent. Genetics of Diabetes Audit, Research in Tayside Scotl, (GoDARTS), Scania Diabetes Registry (SDR), Steno Diabetes Centre, and BENEDICT (phases A and B) conducted in subjects with T2DM. Using eight different DKD phenotypes involved 5,717 T2DM subjects, 3,345 with DKD successfully identified a novel locus, GABRR1 (led by rs9942471) (P < 5 × 10 − 8), for a microalbuminuria phenotype in European subjects with T2DM [47,48,49,50]. However, only one locus reached genome-wide significance: PLCB4 (encoding 1-phosphatidylinositol 4,5-bisphosphate phosphodiesterase β-4) on chromosome 20 on variant rs2206136 was associated with the CKD phenotype (P = 2.1 × 10 − 8).While, two other genes Uromodulin (UMOD) and Protein Kinase AMP-Activated Non-Catalytic Subunit Gamma 2 (PRKAG2), showed association to eGFR, replicate at genome-wide significance [51]. A Genome-wide association studies genetic discoveries in (Table 3).
Table 3 GWAS genetic discoveriesIn another, GWAS comparing African American individuals with T2DM and ESRD with non-diabetic and non-DKD controls identified six independent genome-wide significant associations with ESRD (rs58627064 at chr3q26, independent SNPs rs142563193 and rs142671759 near Ectonucleotide Pyrophosphatase/Phosphodiesterase family member 7 (ENPP7), rs4807299 in Guanine nucleotide-binding protein (GNG7), rs72858591 at chr2q23, and rs9622363 in Apolipoprotein L1 (APOL1) [52].
In 2018, the FTO gene locus showed an association with DKD (SNP rs56094641) (P = 7.7 × 10 − 10) in GWAS of Japanese individuals with T2DM. Furthermore, in 2019, GWAS of UACR was conducted that identified four genome-wide significant signals in the DM subset, namely Kazrin, Periplakin Interacting Protein (KAZN), MIR4432HG- B-Cell CLL/Lymphoma 11A (BCL11A), Forkhead box protein P2 (FOXP2) and Cadherin-2 (CDH2) [52]. The MYH9 (myosin, heavy chain 9)/APOL1 (apolipoprotein L1) locus was associated with T2DM in African Americans, according to results from another high-density GWAS, and these results were replicated in a study of candidate gene and in a familial linkage analysis [55].The MYH9/APOL1 function in DKD is fast growing and remains a hotly debated issue.
The impact of these small risk variants is uncertain on the development and progression of DKD, as it could be induced by the cumulative or synergistic impact of many variations with small risks. However, further studies with bigger patient cohorts are required to find rare but significant variants of DKD. In addition, genetics research utilizing more precisely defined patient populations, such as patients with progressive DKD, will aid in identifying genes related to a more precisely defined disease phenotype. We can only uncover novel rare sequence variations linked with DKD when we can gather sufficiently large sample numbers and use advanced gene aggregation studies.
Epigenetics (non-coding RNA dysregulation)In addition to the qualitative and quantitative control of gene activity provided by genetic variations, epigenetic control of gene regulation adds an extra layer of regulation to genes implicated in the pathogenesis of DKD. Latest developments in large genome-wide screenings and sequencing tools have made it possible to scan epigenetic alteration of the whole genome, known as “epigenome-wide association studies (EWAS),” a significant complementary to GWAS. In the context of DKD, different miRNAs are engaged in pathogenesis-related pathways (apoptosis, fibrosis, and extracellular matrix accumulation). Hyperglycemia causes the release of cytokines, growth factors, and miRNA dysregulation. miRNAs have a role in DKD development by affecting genes involved in different pathways.
miR21The miR-21 located on chromosome 17, has an important role in the development of DN, and enhanced expression has been detected not just in DN tissues but also in DN patients’ plasma and urine [56]. MiR-21 involves in the process of DN, promotes urinary protein excretion, and aggravates renal function damage [57]. Both DN patients and DN mice had higher MiR-21-5p expression in their serum or kidneys. This increased expression in DN correlates with tubulointerstitial fibrosis, renal damage, and decreased eGFR. Furthermore, miR-21-5p influences cardiovascular events associated with Chronic Kidney Disease (CKD) [56].
A prospective case–control study was conducted highlighted the clinical significance of five potential miR’s (miR-21, miR-29a, miR-29b, miR-29c and miR192) in T2DM patients who have existing DR with differential ACR and eGFR was performed using quantitative RT-PCR analysis. It was shown that the miR-21 level was significantly upregulated in the low-eGFR group compared with high-eGFR patients implying the clinical potential of DKD associated miR-21 in monitoring and preventing disease advancement [58].
The first study to identify a specific miRNA involved in DKD discovered that miRNA192 was upregulated in vitro in mesangial cells (MCs) and in vivo in glomeruli from type 1 streptozotocin-induced and type 2 db/db DKD animal models show that MiRNA192 repression may increase collagen deposition in response to TGF‐β [59]. In a systemic review conducted, six miRNAs repeatedly showed to be dysregulated in DKD patients compared to controls in a systemic review: miR-21-5p, miR-29a-3p, miR-126-3p, miR-192-5p, miR-214-3p, and miR-342-3p 59. Bioinformatics investigation revealed that these six miRNAs are involved in DKD pathogenesis pathways such as apoptosis, fibrosis, and extracellular matrix buildup [59].
A study described the link between microRNA-126, T2DM, and DKD expression, comprised of 52 patients with T2DM and normal albuminuria, 50 patients with T2DM with increased albuminuria (29 with moderate to severe and 21 with severe albuminuria), and 50 non-diabetic healthy people [61]. When compared to controls, microRNA-126 expression was considerably lower in T2DM and even lower in people with DKD [59]. A meta-analysis conducted on 2,747 patients showed that the downregulation of miR-126 was significant (OR: 0.57; 95% CI: 0.44–0.74; p-value < 0.0001) in blood from patients with DKD, while its urinary level was significantly upregulated (OR: 2931.12; 95% CI: 9.96–862,623.21; p-value = 0.0059), suggesting microRNA-126 may have important diagnostic and pathogenetic implications for DN [62].
Transforming growth factor-βThe most notable pathological hallmark of ESRD is kidney fibrosis, characterized by increasing tissue scarring that leads to glomerular and tubulointerstitial fibrosis. TGF-β is the primary regulator of this process, acting as the principal driver of matrix production, matrix degradation inhibition, and myofibroblast activation. In addition, TGF-β1, the most abundant isoform of TGF-β, which is released by all types of renal cells and infiltrating inflammatory cells, is a key participant in the pathogenesis of DKD, owing to its intense profibrotic properties. As a result, this cytokine works on various cells in the kidney, including podocytes and tubular epithelial cells and inflammatory cells such as macrophages and T cells.
According to a recent theory, TGF1 may have an essential role in the early stages of DKD development, and several miRNAs and long non-coding RNAs (lncRNAs) control the critical molecules in the TGF1 pathway. These ncRNAs might be used as biomarkers to determine possible targets for DKD prevention and therapy. Specific miRNAs, such as miR-192, miR-216a, miR-217, and miR-377, have been linked to TGF signaling and the pathophysiology of DKD [63].
The predominant pathogenic hallmark of ESRD is kidney fibrosis, which is characterized by increasing tissue scarring that leads to glomerular and tubulointerstitial fibrosis (TIF). The fibrotic role of TGF-β1 is mainly due to the Smad-dependent pathway aft
留言 (0)