
記住我


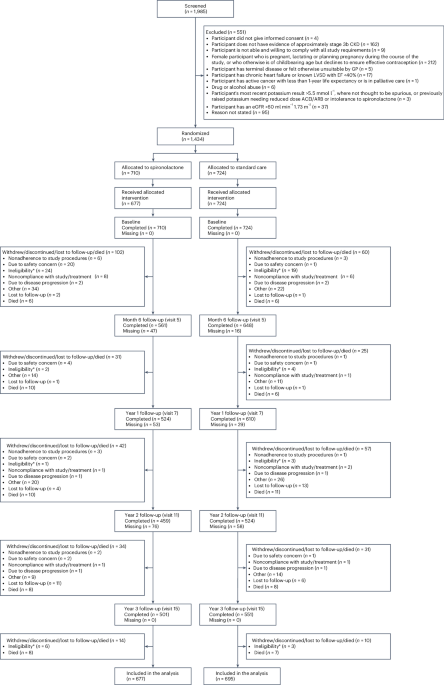






From May 2021 until March 2024, we enrolled 19 healthy adults aged of 20–65 years old (Fig. 1a). The subjects were recruited for BM aspirates 2.5–33 months after receiving the first dose of SARS-CoV-2 mRNA vaccines. All received a total of two to five vaccine doses, and BM aspirates were obtained 0.5–21 months after receiving the last booster (the third, fourth or fifth dose) (Table 1). One subject provided three longitudinal BM samples over a period of 21 months and a second subject had two aspirates over 6 months, resulting in a total of 22 BM aspirates. Five subjects reported infection with SARS-CoV-2 1–16 months before the BM collection, of which three subjects had infection once and two had two PCR-proven SARS-CoV-2 infections. These infections occurred 1–15.5 months after receiving the most recent vaccine dose. All 19 individuals received the quadrivalent influenza vaccine within 1–12 months (relative to the time of each BM aspirate), and one was delayed for 1 year due to the COVID-19 pandemic. All received the childhood series of the tetanus toxoid vaccine with recent boosters ranging from 1 month to 24 years from the time of BM aspirates.
Fig. 1: Demographics of the 19 BM subject cohort.
a, The general FACS gating strategy used for sorting BM ASC subsets. Age given in years. Cauc, Caucasian; Pre-inf, previous (COVID-19) infection. b, BMMCs were first gated for lymphocytes, singlets and viable cells (based on their forward scatter/side scatter (FSC/SSC) and live/death properties). CD3 and CD14 were then used as dump markers to capture CD19+ and CD19− B cell populations. Subsequent subgating from CD19+ population on the IgD− fraction (versus CD27) and using CD138 versus CD38 allowed the breakdown of BM ASC populations into three subsets of interest: PopA (CD19+CD38hiCD138−), PopB (CD19+CD38hiCD138+) and PopD (LLPC; CD19−CD38hiCD138+). PE-Cy7, PE-Cyanine7 tandem fluorophore. c, Representative ELISpot scanned images (left: a vaccinated subject without previous COVID-19 infection; right: a vaccinated subject with previous COVID-19 infection). The numbers of input ASC that were incubated for total IgG: ~687 (left) or ~522 (right), and for antigen-specific ASC IgG: ~2,062 (left) or ~1,566 (right). Each circle represents an individual vaccine subject. The counts were provided by the sorters. Blood ASCs from subjects were collected at the peak time of response (which is 5–7 days post-vaccine). d, Frequencies (%) of antigen-specific IgG ASCs per total IgG ASCs. Data were generated from 3, 10, 13, 6, 15 and 8 different SARS-CoV-2-vaccinated subjects for SARS-CoV-2 antigens NTD, RBD, S1, S2, S2P and NP, respectively. Statistics were assessed using Student’s t-test (two-tailed unpaired t-test) in Excel (Microsoft), and differences were considered significant at P values less than 0.05. For additional antigen selection and validation, see Extended Data Fig. 2. In c and d, it can be seen that S2P was most sensitive to capture SARS-CoV-2-specific blood ASCs isolated after SARS-CoV-2 mRNA vaccines.
Table 1 BM subjects and BM samplesBM ASC subsets and antigen optimizationBM ASC subsets were fluorescence-activated cell sorting (FACS)-sorted according to surface expression of CD19, CD38 and CD13810 (Fig. 1b). To overcome the problem with the rapid death of ASCs ex vivo16, we rested the ASCs overnight in a new human in vitro plasma cell survival system that is capable of maintaining human ASC viability for months16. Since we had previously localized the BM LLPC compartment into PopD (CD19−CD38hiCD138+)10, this population was sorted out of total BM ASCs together with non-LLPC subsets: PopA (CD19+CD38hiCD138−) and PopB (CD19+CD38hiCD138+). All were tested for total IgG secretion as well as influenza (Flu)-, tetanus toxoid (Tet)- and SARS-CoV-2-specific IgG secretion by bulk ELISpots. To optimize antigen detection for the BM ASC ELISpot assays, we collected early-minted blood ASCs (CD27hiCD38hi; Extended Data Fig. 1) 6–7 days after Flu, Tet or SARS-CoV-2 vaccination, which is the peak time for enrichment of vaccine-specific ASCs in the blood after secondary immunization30,31, and performed ELISpots (Extended Data Fig. 2a). Of the SARS-CoV-2 antigens (S1 domain of spike protein (S1), S2 domain of spike protein (S2), receptor-binding domain (RBD), S-2P variant of spike protein (S2P), N-terminal domain (NTD) and nucleocapsid protein (NP)), S2P, a prefusion-stabilized spike trimer32, generated the highest frequency, followed by S1 (with no significant difference; P = 0.21) (Fig. 1c,d), and so S2P was selected for the BM ASC ELISpot assays. We also validated the quadrivalent Flu vaccine (seasons of 2019–2020 to 2023–2024) as the Flu antigen and Tet antigen (Extended Data Fig. 2b,c), using blood ASCs at days 6–7, the peak of the respective ASC vaccine responses30,31.
Absence of SARS-CoV-2-specific IgG BM LLPCsSince BM aspirates can yield variable cell numbers, we included BM aspirates with >3,000 sorted cells in each of the three ASC populations, cultured the cells overnight in a specialized in vitro BM mimetic system16 and performed bulk ELISpots (Fig. 2a; see also Methods). Among the BM subjects, sufficient cells to confidently measure vaccine specificities within PopA, PopB and PopD were obtained from 8, 15 and 17 individuals. As previously shown, all BM ASC subsets had detectable total IgG ASCs. Similar to previous reports10, PopD contained the highest percentage of Flu and Tet IgG ASCs per total IgG ASCs: mean 7.3% (7.31 ± 3.51) and 2.1% (2.14 ± 1.70), respectively (Fig. 2b,c). PopB was readily populated with Flu and Tet IgG ASCs: mean 3.4% (3.43 ± 1.68) and 0.8% (0.77 ± 0.87), respectively, while PopA had the lowest frequencies: mean 1% (1.0 ± 0.66) and 0.2% (0.17 ± 0.17), respectively. Strikingly, within the same subjects, we could rarely detect S2P ASCs in PopD: mean 0.1% (0.14 ± 0.23). In contrast, the S2P specificity was readily found in PopB and PopA at frequencies comparable to Tet and Flu: mean 3.1% (3.13 ± 2.82) and 0.9% (0.89 ± 1.3), respectively.
Fig. 2: Absence of SARS-CoV-2 BM IgG LLPC after SARS-CoV-2 mRNA vaccines by detection of ASC and secreted IgG in the BM ASC culture supernatants.
a, Summary of the techniques and the experimental designs for detection of total, Flu, Tet and S2P ASCs and secreted IgG by ELISpots and MBBA, respectively. MNC, mononuclear cells. b, Representative ELISpot scanned images. The numbers of input ASC that were incubated were ~52 K, ~12.1 K and ~10.1 K for PopA, PopB and PopD, respectively. Each symbol represents an individual vaccine subject for total IgG and antigen-specific ASC from PopA, PopB and PopD. c, ELISpots measuring BM IgG ASC specific for Flu, Tet and S2P. Data were generated from 8, 15 and 17 different SARS-CoV-2-vaccinated subjects for PopA, PopB and PopD, respectively. For individual ratios and statistic comparisons between any two antigens for any subset or between any two subsets for any antigen, see Supplementary Tables 1 and 2, respectively. d, Fold difference (ratios) when comparing different vaccine specificities between non-LLPCs (combined PopA and PopB) versus LLPCs (PopD). e, MBBA measuring IgG specific for Flu, Tet and S2P (normalized to total IgG) from culture supernatant of PopA, PopB and PopD. Supernatant preps were collected from 18–24-h cultures of BM ASCs after revival from the FACS sorters and were quantified for total IgG and vaccine-specific IgG in neat (undiluted). Data were generated from eight different SARS-CoV-2-vaccinated subjects. For individual ratios and statistic comparisons between any two antigens for any subset or between any two subsets for any antigen, see Supplementary Tables 1 and 2, respectively. f, The fold difference (ratios) when comparing normalized vaccine-specific IgG in the supernatants from the culture of non-LLPCs (combined PopA and PopB) versus LLPCs (PopD). For ratio calculation, see Methods. For IgG standard versus MFI curve, see Extended Data Fig. 4. Counts were provided by the sorters. LLPC, boxes in b, c, and e. Sub, subject; Sups, BM ASC culture supernatant preps. For details of subjects and samples, see Table 1.
Although the frequencies of Tet IgG ASC in PopA versus PopB showed no statistically significant difference, the frequencies of Flu IgG were higher in PopB over PopA. For both Flu and Tet IgG ASC, the frequencies in PopD were always higher than in PopB. In contrast, the S2P IgG ASC frequencies were always significantly lower in PopD compared with PopB (Fig. 2b,c). On average, the fold changes of IgG ASC specificities within PopD were 52.8 for Flu:S2P and 15.5 for Tet:S2P (Supplementary Table 1). In comparison, the fold changes of IgG ASC specificities within PopB were 1.1 for Flu:S2P and 0.3 for Tet:S2P. For S2P specificity, the fold changes of PopA:PopD was 6.4 and of PopB:PopD was 22.6 (Supplementary Table 2). In comparison, for Flu or Tet specificities, these fold changes were ≤0.47. Overall, the ratios of non-LLPC:LLPC for Flu, Tet and S2P were 0.61, 0.44 and 29.07, respectively (Fig. 2d). Thus, S2P IgG ASC are largely excluded from PopD.
Absence of SARS-CoV-2-specific IgA BM LLPCSimilar to IgG ASC, the frequencies of Flu and Tet IgA ASC were highest in PopD with a mean of 1.7% (1.70 ± 0.45) and 0.3% (0.31 ± 0.12), respectively, while frequencies in PopA and PopB were lower: for Flu, mean 0.8% (0.82 ± 0.43) and 1.4% (1.35 ± 1.32), respectively, and for Tet, 0.2% (0.24 ± 0.34) and 0.1% (0.11 ± 0.10), respectively (Extended Data Fig. 3a,b). Consistent with previous studies33, these results may be explained by the predominance of IgG responses to the intramuscular tetanus vaccine. S2P IgA ASCs were also detected predominantly in PopA and PopB: a mean of 1.5% (1.46 ± 1.57) and 0.9% (0.90 ± 0.66), respectively, and were virtually absent in PopD: a mean of 0.03% (0.03 ± 0.06) (Extended Data Fig. 3b). On average, the fold changes of IgA ASC specificities within PopD were 50.9 for Flu:S2P and 9.3 for Tet:S2P (Supplementary Table 3). For S2P specificity, the fold change of PopA:PopD was 43.8 and of PopB:PopD was 27.0 (Supplementary Table 4). Thus, similar to IgG ASCs, other class-switched isotypes such as S2P IgA ASC are also mostly excluded from PopD (albeit small sample size).
Absence of SARS-CoV-2-specific IgG in LLPC culture supernatantTo validate the antigen-specific ELISpot results, we measured secreted IgG from BM ASC subsets (Fig. 2a; see also Methods). Briefly, from eight individuals who yielded sufficient sorted cells for all BM ASC subsets (PopA, PopB and PopD), we cultured ASCs in a specialized in vitro BM mimetic system overnight16 and measured the cultured supernatants for secreted IgG specific for Flu, Tet and S2P by multiplex bead-binding assays (MBBAs)34 (Extended Data Fig. 4). The results were similar to the ELISpot: the percentages of Flu and Tet IgG per total IgG were highest in PopD (mean 7.92 ± 7.41 and 7.51 ± 9.98, respectively) compared with PopB (mean 4.09 ± 2.81 and 2.30 ± 2.14, respectively) or PopA (mean 1.12 ± 1.08 and 0.97 ± 2.46, respectively) (Fig. 2e). In contrast, the percentage of S2P IgG per total IgG was lower in PopD (mean 0.12 ± 0.20) compared with PopA (mean 0.31 ± 0.62) and especially with PopB (mean 2.46 ± 1.83).
Of eight individuals, the fold change in PopD for Flu:S2P was 66.5 and for Tet:S2P was 63.1 (Supplementary Table 1). In comparison, the fold change within PopB for Flu:S2P was 1.7 and for Tet:S2P was 0.9, demonstrating similar quantities of IgG to Flu, Tet and S2P in PopB. Within the S2P specificity, the fold changes of S2P IgG levels in the BM culture supernatants for PopA:PopD and PopB:PopD were 2.6 and 20.1, respectively (Supplementary Table 2). In comparison, for Flu or Tet specificities, these fold changes were ≤0.31. Ultimately, using this method of measuring secreted antibodies from the cultured BM ASCs, the ratios of non-LLPC:LLPC for Flu, Tet and S2P from BM ASC culture supernatant were 0.66, 0.44 and 23.26, which was similar to the ELISpot results (Fig. 2f). In all, we validate the antigen specificities observed by the ELISpots using our novel in vitro plasma cell culture methods that also showed exclusion of SARS-CoV-2-specific ASCs in PopD.
No correlation of S2P BM ASC responses and time from first vaccineBecause the time from vaccination to BM aspiration varied among the subjects, we compared the time from the last Flu and Tet vaccine as well as from the first SARS-CoV-2 vaccine with the vaccine-specific BM ASC responses. For Flu and Tet, we saw no correlation between the frequencies of vaccine-specific BM ASC subsets (including PopD) with the time windows since the last Flu or Tet vaccine (R2 ≤ 0.16, P ≥ 0.10; Extended Data Fig. 5a,b). Similarly, the S2P BM LLPC and non-LLPC frequencies did not correlate with the time from the first vaccine (R2 ≤ 0.07, P ≥ 0.29; Extended Data Fig. 5c), the number of vaccine doses (R2 ≤ 0.05, P ≥ 0.39; Extended Data Fig. 6) or age of BM subjects (R2 ≤ 0.03, P ≥ 0.50; Extended Data Fig. 7). These results suggest that more time since SARS-CoV-2 mRNA vaccination or more vaccine doses does not necessarily promote more S2P PopD responses (in our small cohort).
No differences in BM ASC after vaccine or vaccine with infectionWe next compared the S2P BM ASC frequencies in SARS-CoV-2 infected subjects who were also vaccinated (n = 5) with those who were only vaccinated with no self-reported infection (n = 14). Between these two groups, we found no differences in S2P LLPC and non-LLPC responses—stratified either by the time from the first (R2 ≤ 0.10, P ≥ 0.32; Extended Data Fig. 8a) or the last (R2 ≤ 0.19, P ≥ 0.49; Extended Data Fig. 8b) vaccine. Although the small number of samples made it difficult to draw definitive conclusions, these results suggest that SARS-CoV-2 infection may also fail to elicit BM LLPCs.
Declined serum S2P, not Flu, Tet or total, IgGTo assess the kinetics of serum antibodies, we measured total IgG as well as Flu, Tet and S2P IgG responses up to 38 months after the first SARS-CoV-2 vaccine. From subjects with at least two sequential serum samples collected within 5 months of the time of BM aspiration (n = 8), we observed a decline of S2P IgG titers in the serum within 3–6 months post-first SARS-CoV-2 vaccine (Fig. 3a). One subject had a booster at 7 months after the first SARS-CoV-2 vaccine (subject 8) that showed a rise and a rapid fall in antibody titers. While total IgG and Flu and Tet IgG titers in the serum were relatively stable during the period of 38 months after the first SARS-CoV-2 vaccine in this cohort, serum S2P IgG levels declined within 3–6 months of vaccination unless boosted by additional SARS-CoV-2 vaccines.
Fig. 3: Vaccine-specific IgG levels in the serum: kinetics and magnitude, and correlation with BM IgG ASC responses.
a, Kinetics and magnitude of IgG titers from subjects (n = 8) with at least two sequential serum samples (collected before the additional SARS-CoV-2 vaccines). b–d, Serum IgG levels versus BM IgG LLPC, non-LLPC and total ASC responses for Flu (b), Tet (c) and S2P (d) specificities in all examined subjects (n = 19). Sera collected within 5 months of the time of BM aspiration. For b–d, data were generated from eight different SARS-CoV-2-vaccinated subjects and correlations were assessed using simple linear regression analysis performed with GraphPad Prism (GraphPad Software). The exact P values for vaccine-specific LLPCs, non-LLPCs and total ASCs are 0.0043, 0.0196 and 0.0075, respectively (b); <0.0001, 0.91 and <0.0001, respectively (c); and 0.6096, 0.0025 and 0.0008, respectively (d). All serum samples tested at dilutions of 1:1,000–1:100,000 (total IgG) or 1:200–1:16,000 (antigen-specific IgG). For serum total and vaccine-specific IgG standard curves, see Supplementary Fig. 1. For details of subjects and samples, see Table 1.
Correlation of serum S2P IgG and BM IgG non-LLPCsTo investigate the relationship between systemic antibodies and BM ASC responses, we analyzed the IgG titers in the serum and the frequencies of BM IgG ASCs (which include LLPCs, non-LLPCs and the sum of both as the total vaccine-specific ASCs). We saw a modest correlation between serum Flu IgG and total BM Flu IgG ASCs (R2 = 0.35, P < 0.01; Fig. 3b). When separating LLPCs and non-LLPCs from the total vaccine-specific ASCs, there were also modest correlations of serum Flu IgG titers with BM Flu IgG non-LLPCs (R2 = 0.35, P = 0.02) or with LLPCs (R2 = 0.43, P < 0.01). Interestingly, for Tet, we observed a very strong correlation between serum IgG titers and BM IgG LLPCs (R2 = 0.83, P < 0.01) or total ASCs (R2 = 0.78, P < 0.01) but not non-LLPCs (R2 < 0.01, P = 0.91) (Fig. 3c). On the contrary, for S2P, we found a significant correlation between serum IgG levels and BM IgG non-LLPCs or total ASCs (R2 = 0.52, P < 0.01 or R2 = 0.49, P < 0.01, respectively) but no correlation for LLPCs (R2 = 0.02, P = 0.61) (Fig. 3d). Together, these results show that serum Tet and Flu but not S2P IgG levels largely correlate with the vaccine-specific BM IgG LLPC responses; in contrast, serum IgG levels for S2P specificity are associated with the S2P BM IgG non-LLPC frequencies.
Kinetic responses for IgG ASCs in longitudinal BM aspiratesWe next assessed the IgG ASC kinetic responses in a subject (subject 14) who provided three sequential BM aspirates over a period of 23 months. BM aspirates were taken 2.5, 14 and 23 months after the first SARS-CoV-2 vaccine (Table 1). Seven serum samples were collected within months of each BM aspiration. Each BM aspirate provided >3,000 FACS-sorted ASCs in each subset. Again, total IgG ASCs were detected in all BM PopA, PopB and PopD. We observed an increase in the frequencies of S2P IgG ASCs in PopA and PopB at 14 months (1.07% and 9.02%, respectively) and 23 months (3.98% and 6.24%, respectively), compared with the first time point (0.90% and 0.38%, respectively) (Fig. 4a,b). However, in PopD, there were no S2P IgG ASCs detected at the first two time points and only 0.31% at the last one (23 months). Notably, at the earliest time point (2.5 months), the highest S2P IgG ASC frequency was observed in PopA, then at both later time points (14 months and 23 months), it was highest in PopB. In all, regardless of time points, the S2P ASC frequencies were always higher in PopA and PopB compared with PopD (even at 23 months). We observed the highest Flu and Tet frequencies in PopD, followed by PopB, and lowest in PopA. Interestingly, the Flu and Tet BM ASC frequencies were quite consistent over the course of 2 years.
Fig. 4: Kinetics and magnitude of BM IgG ASC responses and of total and vaccine-specific serum IgG levels in the subject who donated three longitudinal BM aspirates over 2 years (subject 14).
a, ELISpot scanned images. The numbers of input ASC that were incubated were ~21K, ~40K and ~4.9K (2.5 months); ~14K, ~12K and ~3.8K (14 months); and ~58K, ~22K and ~7.2K (23 months) for PopA, PopB and PopD, respectively. b,c, The kinetics and magnitude of antigen-specific BM IgG ASC responses (b) and of total and antigen-specific IgG levels in the serum (c). Sera were collected within 1–5 months of the time of BM aspiration. Part of Fig. 4c is reproduced from Fig. 3a for the purpose of kinetics comparison. Sub, subject; K, 1,000. Counts were provided by the sorters. LLPC, boxes in a and b. All serum samples were tested at dilutions of 1:1,000–1:100,000 (total IgG) or 1:200–1:16,000 (antigen-specific IgG). For serum total and vaccine-specific IgG standard curves, see Supplementary Fig. 1. For details of subjects and samples, see Table 1.
In the serum, as expected, total IgG as well as Flu and Tet IgG levels were stable during the examined time periods (Fig. 4c). In contrast, after an initial decline within 3–6 months after SARS-CoV-2 vaccination, S2P IgG titers remained at low levels for about seven months. They then increased significantly, corresponding to the high frequencies of S2P PopB in the BM at 14 months and 23 months (Fig. 4b). Importantly, this increase occurred without any additional SARS-CoV-2 vaccination and stayed elevated for about 6 months, suggesting asymptomatic and/or unreported infections.
In a second subject (subject 19) with two sequential BM aspirates collected at 28 months and 33 months after the first SARS-CoV-2 vaccine (Table 1), the vaccine-specific IgG ASC responses in the BM were quite similar: the highest frequency of S2P ASCs was found in PopB (3.25% and 3.34%), followed by PopA (0.46% and 0.35%) and PopD (0.40% and 0.33%) (Extended Data Fig. 9a,b). During this period, Flu and Tet BM ASC responses remained stable with the highest in PopD (Flu: 14.28% and 13.68%, and Tet: 3.97% and 3.67%, respectively) compared with PopB (Flu: 4.11% and 4.05%, and Tet: 0.43% and 0.24%, respectively) and PopA (Flu: 1.86% and 1.74%, and Tet: 0% and 0%, respectively). Thus, analysis of longitudinal BM aspirates demonstrates that S2P BM IgG ASC responses were consistently higher in PopA and PopB compared with PopD, suggesting S2P ASCs are not established in the BM LLPC compartment after almost 3 years since the primary SARS-CoV-2 mRNA vaccination.
Few subjects show S2P BM LLPC at low frequenciesFinally, when we calculated the number of individuals with S2P-positive responses for each BM ASC subset, S2P IgG ASCs were easily detected in PopA in 6/8 (75%) individuals and in PopB, in all 15/15 (100%) subjects (Fig. 5a). Only 6/17 (35.29%) subjects had S2P IgG ASCs in PopD, and all were extremely low frequencies despite four or five doses of the vaccine and multiple known SARS-CoV-2 infections. As expected, nearly all subjects had easily detectable Flu and Tet specificities in PopD: 17/17 (100%) and 16/17 (94.12%) respectively. Altogether, durable serologic immune response correlates well with the abundance of Flu and Tet BM ASCs in PopD, while short-lived serologic antibody responses to SARS-CoV-2 mRNA vaccines may be explained by the exclusion of S2P ASCs from this compartment (summarized in Fig. 5b).
Fig. 5: SARS-CoV-2 BM IgG LLPCs are not durably established after mRNA vaccination.
a, Antigen specificity strata of all individuals examined for each BM ASC subset. n, number of BM donors. b, Graphical summary. The majority of SARS-CoV-2 plasma cells are not established from the BM LLPC compartment 33 months after mRNA vaccination. Created with BioRender.com.
留言 (0)