
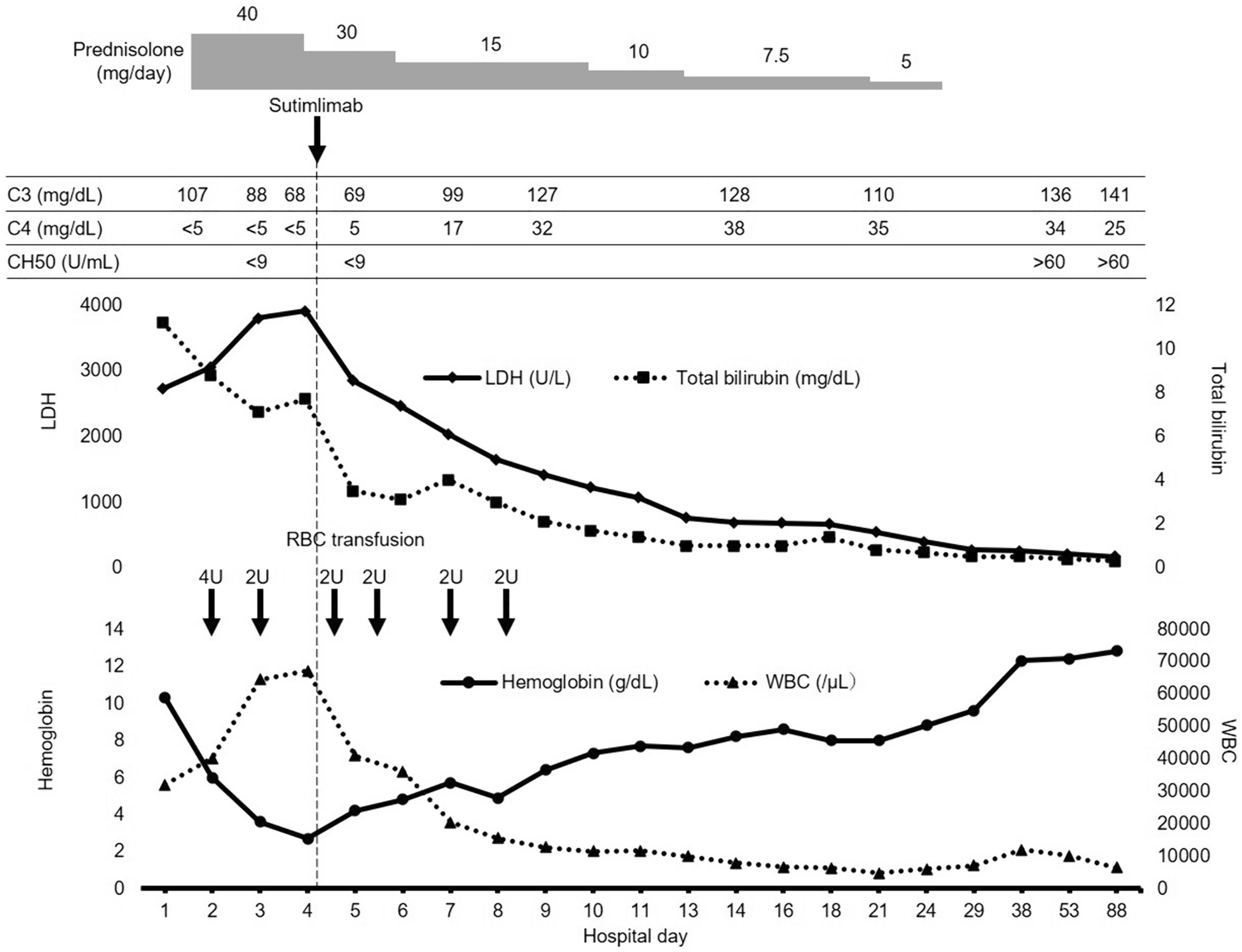
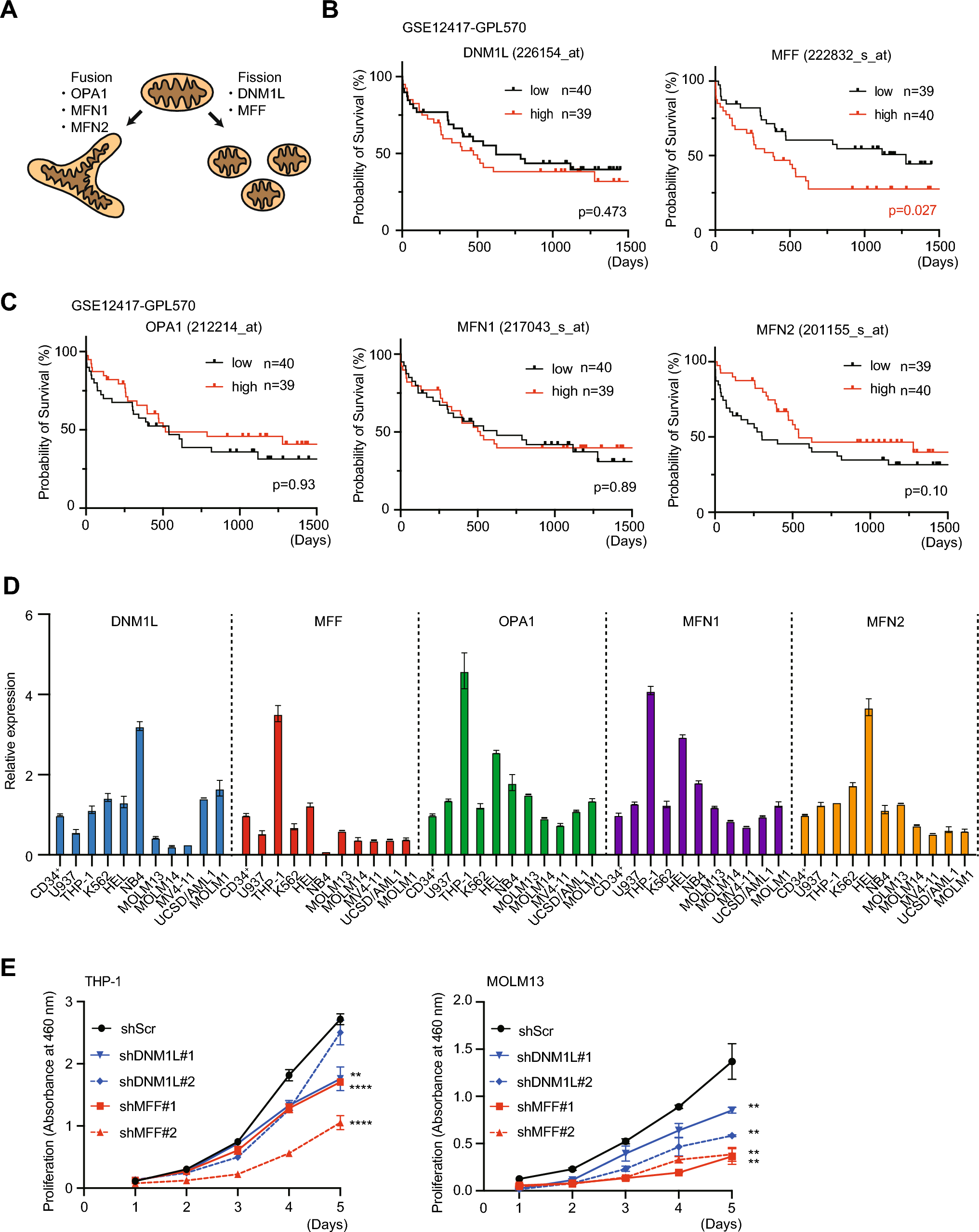
記住我








We examined the expansion effects of romiplostim, rhTPO, and eltrombopag on HSPCs. We used CD34 + cells derived from human CB, as HSPCs are highly enriched in CB. The results showed that all three TPOR agonists induced expansion of CD34 + cells when compared with the vehicle, and a similar result was obtained in samples collected from different donors (Fig. 1a, also see Fig. S1a for the gating strategy). Romiplostim demonstrated a concentration-dependent increase in CD34 + cell count. In contrast, eltrombopag demonstrated a concentration-dependent effect (up to 1000 ng/mL), but the CD34 + cell count declined at a concentration of 10,000 ng/mL. The maximum efficacy of romiplostim was similar to that of rhTPO but higher than the maximum efficacy of eltrombopag. Next, we examined which subset within the CD34 + cell population was affected by each stimulation. The more primitive cells were defined as CD34 + CD38 − , and the more differentiated cells were defined as CD34 + CD38 + (Fig. S1b). The absolute count of CD34 + CD38 − cells at maximum efficacy was different between stimulants, with romiplostim demonstrating higher activity than eltrombopag (Fig. 1b). The absolute count of CD34 + CD38 + cells at maximum efficacy was higher with romiplostim than with eltrombopag, but this difference between the two stimulants was smaller in CD34 + CD38 + cells than that in CD34 + CD38 − cells (Fig. 1c). The maximum effect of romiplostim reached the effect of rhTPO in both subsets (Fig. 1b and Fig. 1c). Interestingly, the expansion effect of eltrombopag at 10,000 ng/mL was lower than that at a lower concentration (1000 ng/mL) in both CD34 + CD38 − cells (Fig. 1b) and CD34 + CD38 + cells (Fig. 1c). Similar results were obtained with the culture condition containing SCF and FLT3L, which are the commonly known cytokines for expanding HSPCs (Fig. S2) [23]. Altogether, these results suggest that romiplostim induces expansion of HSPCs, including the primitive subsets, in a manner similar to that of rhTPO and at higher efficacy than that of eltrombopag in vitro.
Fig. 1
Romiplostim induces expansion of CD34 + cells. (a, b, c) Human cord blood–derived CD34 + cells were treated with romiplostim (0.01, 0.1, 1, 10, 100, or 1000 ng/mL), eltrombopag (0.1, 1, 10, 100, 1000, or 10,000 ng/mL), recombinant human thrombopoietin (10 ng/mL), or vehicle (DMSO) for 7 days. The cells were then collected and counted by flow cytometry. Absolute cell count per assay volume (cells/µL) of total CD34 + cells (a), CD34 + CD38 − cells (b), and CD34 + CD38 + cells (c) are shown. (a) Two different results of the same experiment performed using two different donor cells. Data are shown as mean ± standard error of the mean, n = 3. (b) and (c) Results obtained using CD34 + cells from donor 1
Romiplostim affects the megakaryocytic trajectory in HSPCs similar to rhTPO and eltrombopagHSPCs are heterogeneous cells comprising HSCs, MPPs, and lineage-committed progenitor cells. Given that we had observed the expansion effects of romiplostim on both CD34 + CD38 − and CD34 + CD38 + subsets, we next attempted to identify which subset was affected by romiplostim and to examine whether the effects of romiplostim at the molecular level was similar to those of rhTPO and eltrombopag. For this, we performed a single-cell RNA-seq analysis. Human CB-derived CD34 + cells were stimulated with romiplostim, eltrombopag, rhTPO, or the vehicle DMSO in a serum-free suspension culture for 2 days and proceeded to single-cell RNA-seq analysis. As the expanding effects of romiplostim, rhTPO, and eltrombopag were similar whether or not SCF and FLT3L were included in the culture, the assay was done without SCF and FLT3L to observe the specific effects of these stimulants. This early time point was chosen to investigate the primary effect of stimulation and to avoid the effects secondary to proliferation. The tested concentration was selected at the point approximate to the Cmax value or the serum concentration described earlier (10 ng/mL for romiplostim and rhTPO). Eltrombopag was tested at two concentrations covering the Cmax value; at the concentration of 1 µg/mL, which demonstrated the maximum efficacy in expanding CD34 + cells, and at the concentration of 10 µg/mL, which demonstrated slight efficacy in expanding CD34 + CD38 − cells without expanding CD34 + CD38 + cells (Fig. 1).
Based on single-cell RNA-seq, 21,697 cells were collected and sequenced with adequate reads, that is, approximately hundred thousand or more per cell (Table S1). First, 14 clusters were obtained as a result of unsupervised clustering. The representative blood cell subsets were annotated, as they express specific genes, and were classified as follows: Cluster C6, megakaryocytes with relatively high expression of ITGA2B and PF4; C7, erythroid cells with HBD expression; C9, neutrophils/myeloid cells with expression of IRF8 and ELANE; C10, eosinophils/basophils/mast cells with expression of MS4A2; and C11, lymphoid cells with expression of JCHAIN (Fig. 2a and 2b). Next, we classified cluster C1 as a population enriched in HSCs/MPPs, as they had relatively high expression of AVP (Fig. 2a and 2b). The rest of the clusters were presumed to be either progenitor-enriched cells or difficult-to-classify cells.
Fig. 2
Romiplostim affects the megakaryocytic trajectory in HSPCs in a manner similar to that of recombinant human thrombopoietin (rhTPO) and eltrombopag. Single-cell RNA-seq was performed on human cord blood–derived CD34 + cells stimulated with thrombopoietin receptor agonists for 2 days in culture. (a) Uniform Manifold Approximation and Projection (UMAP) plot of unsupervised clustering of diversely expressed genes against the integrated data of all samples. Major blood cell populations were determined according to the transcriptome profiles and annotated to the corresponding cluster. (b) Expression of representative genes associated with hematopoietic stem cells (AVP), megakaryocytes (ITGA2B and PF4), eosinophils/basophils/mast cells (MS4A2), erythroids (HBD), myeloids (IRF8), lymphoids (JCHAIN), and neutrophils (ELANE) mapped on UMAP plot. (c) UMAP plot of each sample stimulated with either romiplostim, rhTPO, or eltrombopag at the indicated concentration or vehicle (DMSO). (d) Bar graph of cluster distribution for each sample. Cluster distribution was calculated as the relative value of one as a whole. (e) Pseudotime analysis was performed against the integrated data when cluster C1 was selected as the starting point. (f) Thrombopoietin activation score was generated and compared between samples on clusters C1 through C6. Upregulated score (top) and downregulated score (bottom) are shown individually. A dotted line is drawn on the mean value of the vehicle
To characterize the effects of romiplostim on CD34 + cells, we first compared the results of romiplostim with the vehicle, rhTPO, and eltrombopag. Quantitative analysis for the proportion of each cluster (Fig. 2c and 2d) revealed that while the vehicle barely showed distribution at clusters C3, C4, C5, and C6, romiplostim stimulation induced remarkable expansion of these clusters, consistent with the results obtained with rhTPO and eltrombopag. Trajectory analysis revealed that cluster C1, marked as HSCs, proceeded to C6, annotated as megakaryocytes, through C2, C3, C4, and C5 (Fig. 2e) [24]. GO enrichment analysis of specifically expressed genes against clusters C1 through C6 showed that C4 and C5 were enriched for the cell cycle process (Fig. S3 and Table S2). Collectively, as romiplostim stimulation forced cells to distribute into clusters that differentiate into megakaryocytes and because these clusters were significantly cycling, we concluded that romiplostim induced proliferation of cells along the trajectory where HSCs differentiate into megakaryocytes. Moreover, a similar effect was induced by rhTPO and eltrombopag.
We next examined the similarity of rhTPO and romiplostim as well as eltrombopag in terms of TPOR signaling. Therefore, we generated a score that reflects the activation of TPOR signaling established by Comoglio et al. [17] and compared it between stimulants in clusters C1 through C6 individually. This score was calculated based on the gene expression levels of 23 genes with upregulated expression, including Myc, and downstream targets of JAK/STAT signaling, or 20 genes with downregulated expression, such as Sox4 and Hlf, which are rapidly induced by recombinant mouse TPO stimulation. Indeed, rhTPO, compared with vehicle, induced upregulation of the upregulated score and downregulation of the downregulated score on every cluster investigated, assuring that this prediction score is effective in our system (Fig. 2f). The results showed that romiplostim and eltrombopag induced qualitative changes in scores in the same manner as rhTPO from clusters C1 through C6 (Fig. 2f). This suggests that romiplostim, along with eltrombopag, activates TPOR signaling along the megakaryocytic trajectory.
Romiplostim affects the primitive subsets of HSPCsThe durable recovery of hematopoiesis in AA is dependent on the expansion, self-maintenance, and differentiation of HSCs. Therefore, we then aimed to identify the specific effects of romiplostim and other TPOR agonists on the most primitive subsets of HSPCs in the single-cell RNA-seq analysis. Differentially expressed genes (DEGs) between each TPOR agonist and vehicle were generated, and pathway analysis was performed for clusters C1 and C2 individually (Tables S3 and S4). Figure 3a shows the lists of pathways generated using DEGs between romiplostim and vehicle. Figure 3b shows a heat map comparison of all pathways developed by each TPOR agonist. Eltrombopag at a concentration of 1 µg/mL in cluster C1 had a very low amount of DEGs to generate pathway lists (Table S3).
Fig. 3
Romiplostim affects the primitive subsets of hematopoietic stem and progenitor cells (HSPCs). (a) Lists of pathways generated using differentially expressed genes (DEGs) at log fold-change of over 0.18 between romiplostim and vehicle (DMSO) against clusters C1 and C2. The pathways were filtered with − log10 (p-value) ≥ 2, and the top 30 pathways with the highest − log10 (p-value) are listed in the descending order. Pathways for which z-score was not determined were excluded. (b) Heat map of pathways compared between thrombopoietin receptor agonists. Pathways were generated as described above and listed in the order of z-score. Bubble size was associated with − log (p-value)
One of the top pathways significantly induced by romiplostim in clusters C1 and C2 was EIF2 signaling (p-value < 0.01), which is the key pathway for protein synthesis. Moreover, we found this signature to be significantly upregulated by rhTPO (p-value < 0.01) and 10 µg/mL eltrombopag (p-value < 0.01).
Another pathway significantly induced by romiplostim in both clusters C1 and C2 was oxidative phosphorylation (p-value < 0.01). Similar terms such as mitochondrial dysfunction and sirtuin signaling pathway were also listed. Additionally, rhTPO (p-value < 0.01) and eltrombopag (p-value < 0.01) also significantly induced these pathways.
Interestingly, some pathways were induced by romiplostim and rhTPO but not by eltrombopag. For instance, Ras homolog family member A (RHOA) signaling was listed as being significantly induced by romiplostim (p-value < 0.01) in both clusters C1 and C2 but not by 10 µg/mL eltrombopag (p-value = 0.37 and 0.61, respectively). The induction of RHOA signaling by rhTPO was significant in cluster C1 but not in C2 (p-value > 0.01 and = 0.02, respectively). Similar terms such as actin cytoskeleton signaling were also listed.
Taken together, while we observed similarities between romiplostim, rhTPO, and eltrombopag in terms of cluster distribution of HSPCs, as well as in TPO score along the megakaryocytic trajectory, the specific changes induced in the most primitive subsets were partially different.
Eltrombopag at a high concentration induces TFRC expression not observed with romiplostim and rhTPOWhile we observed the inhibition of CD34 + cell expansion with eltrombopag at 10 µg/mL in the proliferation assay (Fig. 1a), single-cell RNA-seq analysis demonstrated that this high concentration induced distribution and TPOR signaling on megakaryocytic trajectory, similarly to rhTPO and romiplostim. Based on this, we presumed that a unique characteristic of 10 µg/mL eltrombopag that is independent of TPOR may exist [25]. Therefore, to gain insights into the biology underlying the inhibitory effect, we searched for DEGs unique to 10 µg/mL eltrombopag. For this, we compared 10 µg/mL eltrombopag to the integrated data of rhTPO, romiplostim, and 1 µg/mL eltrombopag against total cells. The results showed that TFRC, which codes for transferrin receptor, was one of the genes with significantly higher expression (Adjusted p-value < 0.01) when stimulated with 10 μg/mL eltrombopag compared to the integrated data of other groups (Fig. 4a and Table S5). A comparison between individual samples demonstrated that TFRC was uniquely induced by 10 µg/mL eltrombopag (Fig. 4b). Analysis against individual clusters also presented similar results (Fig. 4c).
Fig. 4
Eltrombopag at a concentration of 10 µg/mL has a unique effect. (a) Volcano plot of differentially expressed genes (DEGs) between eltrombopag 10 µg/mL and the integrated data of recombinant human thrombopoietin (rhTPO), romiplostim, and eltrombopag 1 µg/mL against total cells. The adjusted p-value was calculated based on bonferroni correction using all genes in the dataset. Dotted lines are drawn on fold change of 1.25. Red dots represent DEGs with higher expression on stimulation with eltrombopag 10 µg/mL, and blue dots represent DEGs with lower expression on stimulation with eltrombopag 10 µg/mL. (b, c) Gene expression of TFRC in each sample within total cells (b) or individually in clusters C1 through C9 (c)
Eltrombopag has an iron-chelating property, which presumably suggests that upregulated expression of TFRC is a phenotype for the iron demand [26]. Indeed, iron chelation can upregulate TFRC expression and inhibit cell proliferation [27, 28]. Consistent with this, MKI67 expression, a proliferation marker, was relatively low when stimulated with 10 µg/mL eltrombopag (Fig. 4a). Therefore, high concentration of eltrombopag induces TPOR signaling and TPOR signaling-independent chelating activities, and the latter effect may be the cause of the inhibition of cell proliferation.
留言 (0)