
記住我







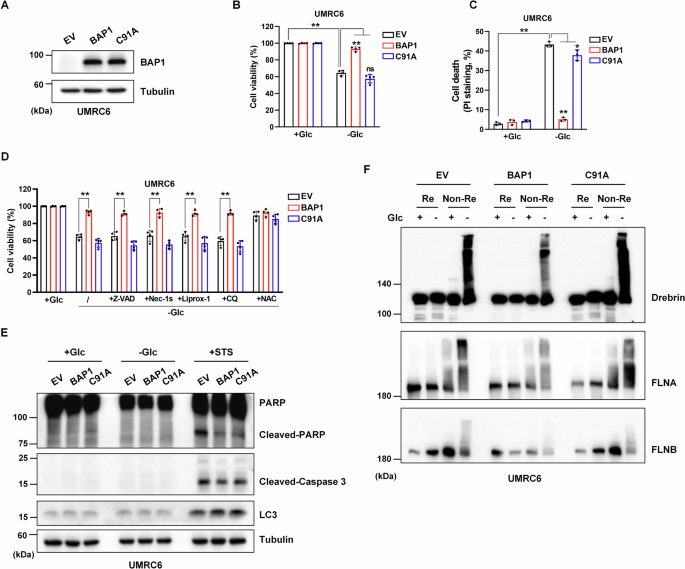
UMRC6, which is deficient in BAP1 gene expression, is an ideal cell line to study the role of BAP1 upon glucose deprivation. First, UMRC6 cell lines stably expressing BAP1, C91A mutant (lack of BAP1 DUB activity), and empty vector (EV) were constructed (Fig. 1A). As shown in Fig. 1B, C, UMRC6-EV cells died in a short period upon glucose starvation, and BAP1, rather than its mutant C91A, protected the cell death under this condition. Next, several known cell death inhibitors were used to figure out the cell death modality that BAP1-protected above. Apoptosis inhibitor Z-VAD, necroptosis inhibitor Nec-1s, ferroptosis inhibitor Liprox-1, and autophagy inhibitor CQ did not prevent cell death in UMRC6-EV and -C91A cells cultured in glucose-free medium, indicating that BAP1 protects a novel type of cell death upon glucose starvation (Fig. 1D). Dai et al. reported that BAP1 inhibited apoptosis under glucose starvation in UMRC6 cells [17]. We observed a similar phenomenon that the expressions of cleaved-PARP and cleaved-Caspase-3 were less upregulated in UMRC6-BAP1 cells than in UMRC6-EV and -C91A cells when they were treated with STS, an apoptosis inducer. While these apoptotic and autophagic markers were not found expressed in the three cell lines during a relative short period of glucose deprivation (Fig. 1E).
Fig. 1: BAP1 suppresses disulfidptosis induced by glucose starvation.
A Validation of the constructed BAP1 WT and BAP1-C91A overexpression in UMRC6 cell lines. B, C Restoring BAP1 WT instead of C91A protects cells from dying upon glucose deprivation. D, E Glucose deprivation induces a unique type of cell death in BAP1-low expression or C91A mutant UMRC6 cells. F Reducing and non-reducing Western blotting analysis of actin cytoskeleton proteins in UMRC6-EV, -BAP1, and -C91A cells upon glucose deprivation condition. The above treatments were terminated when glucose starvation caused significant death of UMRC6-EV cells. Z-VAD, 5 μM; Nec-1s, 2 μM; Liprox-1, 5 μM; CQ, 20 μM; NAC, 2 mM.
Recently, Liu et al. reported a novel cell death modality under glucose deprivation conditions regulated by SLC7A11, namely disulfidptosis [16]. Our previous study revealed that BAP1 could repress SLC7A11 expression by decreasing H2Aub occupancy on the SLC7A11 promoter in a DUB-dependent manner [7]. Here, we assumed that the cell death mode protected by BAP1 upon glucose starvation is disulfidptosis. To confirm our hypothesis, the non-reducing Western blotting assays were used to detect the tailing extent of actin cytoskeleton proteins. When the proteins are overloaded by disulfide bonds, their electrophoretic mobility will be affected under non-reducing conditions, which exhibits a slower migration and trailing band. In addition, the accumulation of disulfide bonds leads to excessive molecular weight of the protein and the inability to transmembrane. Therefore, if there is no trailing band, the reduces basal expression level of protein under non-reducing conditions also suggests excessive disulfide pressure. Under non-reducing conditions, following glucose starvation for about 6 h, actin cytoskeleton proteins including Drebrin, MYH9, FLNA, and FLNB showed significantly slower migration with smears in all the three UMRC6 cell lines. However, band trailing of these actin cytoskeleton proteins were less severe in UMRC6-BAP1 cells than in the other two cell lines (Fig. 1F). Together, BAP1, rather than its DUB-deficient mutant C91A, can rescue glucose starvation-induced disulfidptosis in UMRC6 cells, further demonstrating the importance of BAP1’s DUB activity in regulating disulfidptosis.
BAP1 regulates disulfidptosis through the inhibition of SLC7A11-mediated cystine uptakeWe further studied the role of SLC7A11 in BAP1-mediated protection against disulfidptosis. First, we attempted to construct a stable cell line overexpressing SLC7A11 based on the UMRC6-BAP1 cell line. However, the BAP1 level decreased with the increase of the SLC7A11 expression level, indicating that SLC7A11 may also have a regulatory effect on BAP1 (Fig. 2A). SLC7A11 overexpression restored the sensitivity of UMRC6-BAP1 cells to glucose deprivation-induced cell death (Fig. 2B, C). Compared to BAP1 cells, the SLC7A11 overexpressed BAP1 cells showed an enhanced band trailing under non-reducing Western blotting condition upon glucose starvation (Fig. 2D). Subsequently, the SLC7A11 was knocked out in UMRC6-EV, -BAP1 and -C91A cell lines (Fig. 2E). With the knockout of SLC7A11, the inherent sensitivity of UMRC6-EV and -C91A cells to glucose starvation also disappeared (Fig. 2F). These results suggest that the protective role of BAP1 against disulfidptosis is through SLC7A11.
Fig. 2: BAP1 regulates disulfidptosis through the inhibition of SLC7A11-mediated cystine uptake.
A Validation of BAP1 and SLC7A11 dual-overexpressed UMRC6 cell line. B, C SLC7A11 overexpression counteracts the protective role of BAP1 on UMRC6 cells upon glucose deprivation. D SLC7A11 overexpression enhances the actin cytoskeleton proteins tailing under non-reducing conditions in UMRC6-BAP1 cells upon glucose starvation. E Validation of SLC7A11 knockout in UMRC6-EV, -BAP1, and -C91A cells. F Knockout of SLC7A11 saves glucose starvation-induced cell death in UMRC6-EV and -C91A cells. G, H Erastin (10 μM) treatment saves disulfidptosis in UMRC6-EV and C91A cells upon glucose deprivation. I Cystine deprivation saves glucose starvation-induced cell death in UMRC6-EV and -C91A cells, while 2 mM cystine treatment enhances cell death in UMRC6-EV, -BAP1, and -C91A cells. J Cystine at 2 mM aggravated disulfide stress in UMRC6-BAP1 cells cultured in glucose-free and glucose-containing medium. The above treatments were terminated when glucose starvation caused significant death of UMRC6-EV cells.
SLC7A11-mediated cystine uptake plays a crucial role in promoting glutathione biosynthesis and suppressing ferroptosis. However, high rates of cystine uptake can cause aberrant accumulation of disulfide, leading to disulfide stress and even worse disulfidptosis [16]. It was mentioned in our previous work that restoration of BAP1, but not C91A, inhibits cystine uptake [7]. Thus, we sought to clarify the role of SLC7A11-mediated cystine uptake in BAP1-regulated disulfidptosis. As shown in Fig. 2I, J, cystine depletion rescued UMRC6-EV and -C91A cells from disulfidptosis upon glucose deprivation. While, cystine treatment at 2 mM enhanced disulfidptosis in both cell lines, and even induced disulfidptosis in UMRC6-BAP1 cells under glucose-free condition. High concentration of cystine not only aggravated disulfidptosis of UMRC6-BAP1 cells in glucose deficiency state but also increased the disulfide stress in the state of glucose supplementation. In addition, erastin, a selective inhibitor of SLC7A11 that is able to block SLC7A11-mediated cystine uptake, also saved UMRC6-EV and -C91A cells from disulfidptosis upon glucose starvation (Fig. 2G, H). Together, our data indicate that cystine uptake by SLC7A11 is indispensable in the BAP1-mediated disulfidptosis regulation.
BAP1 decreases NADP+/NADPH ratio within cellsLiu et al. highlighted the close relationship between NADPH depletion and disulfidptosis occurrence [16]. In SLC7A11 high-expression cells, the cystine uptake rate is increased, and the reduction of cystine to cysteine requires the consumption of NADPH. The PPP is the main source of intracellular NADPH. Upon glucose deprivation, the supply of NADPH in SLC7A11 cells was insufficient and the consumption remained high, resulting in the aberrant accumulation of intracellular cystine leading to increased disulfide stress and ultimately disulfidptosis. 2DG, a glucose analog, can mitigate SLC7A11-mediated disulfidptosis by supplying NADPH through PPP. In our study, 2DG reduced the NADP+/NADPH ratio upon glucose deficiency and correspondingly rescued cell death of UMRC6-EV and -C91A cells (Fig. 3A, B). Notably, the NADP+/NADPH ratio was less upregulated in UMRC6-BAP1 cells than in the other two cells lines upon glucose starvation, suggesting that its protective effect against disulfidptosis is partly achieved by regulating the NADP+/NADPH ratio (Fig. 3B). As exhibited in Fig. 3C, band trailing phenomenon was also reduced in 2DG-treatment groups. Notably, UMRC6 cells with high BAP1 expression showed lower NADP+/NADPH levels at a basal level, which may also help them resist disulfidptosis (Fig. 3A, D).
Fig. 3: BAP1 decreases NADP+/NADPH ratio within cells.
A Glucose deprivation upregulates NADP+/NADPH ratio in three UMRC6 cell lines (but to a lesser extent in BAP1 cells), and 2DG reverses the upregulation in this case. B 2DG rescues cell death in UMRC6-EV and -C91A cells upon glucose deprivation. C 2DG alleviates the actin cytoskeleton proteins tailing under non-reducing conditions in all three UMRC6 cell lines upon glucose starvation. D–G SLC7A11 overexpression (D, F) and 2 mM cystine stimulation (E, G) upregulate NADP+/NADPH ratio and intensify actin cytoskeleton proteins tailing in UMRC6-BAP1 cells upon glucose starvation, which can be rescued by 2DG treatment.
Subsequently, we further verified the regulation of BAP1 and SLC7A11 on NADP+/NADPH ratio. Both SLC7A11 overexpression and high-concentration cystine stimulation can increase the NADP+/NADPH ratio in glucose starvation conditions, which can be inhibited by 2DG treatment (Fig. 3D, E). The results of the non-reducing Western blotting experiment were consistent: 2DG alleviated SLC7A11 overexpression- or 2 mM cystine treatment-induced actin cytoskeleton proteins tailing in UMRC6-BAP1 cells upon glucose starvation (Fig. 3F, G).
Positive correlation between BAP1 and NADPH-related genes expression in KIRC patientsWe divided KIRC patients into high- and low-BAP1 expression groups according to the median BAP1 expression level. The expression levels of NADPH-related genes in the two groups were shown in Fig. 4A, B. Except for GPX3, the remaining NADPH-related genes with significant differences were positively correlated with the expression level of BAP1. Subsequently, we used Spearman’s correlation analysis to analyze the correlation between BAP1 and NADPH-related genes, and intersected with the significantly differentially expressed genes in Fig. 4A, and a total of 13 genes were obtained (Fig. 4C). Next, we examined the expression levels of these 13 genes in the three UMRC6 cells, and found that the expression levels of DHFR2, G6PD, ME3 and GLUD1 in UMRC6-BAP1 cells were significantly higher than those in EV cells (Fig. 4D). NADPH provides high energy electrons for antioxidant defense and reductive biosynthesis. Pentose phosphate pathway (PPP) is essential for the recycle of NADP to NADPH in the cytoplasm and is therefore necessary to maintain the normal NADPH/NADP ratio. In addition, it’s also indispensable for mammalian dihydrofolate reductase (DHFR) activity and folate metabolism [18]. G6PD (glucose-6-phosphate 1-dehydrogenase) catalyzes the rate-limiting step of PPP and is regulated by the NADP+/NADPH ratio [19]. Studies have shown that G6PD deficient cells having impaired growth and oxidative defense because they fail to maintain a low NADP+/NADPH ratio [18]. DHFR is a key enzyme in folate metabolism. In most cell types, although folate metabolism does not produce substantial cytoplasmic NADPH, it can markedly promote the amount of mitochondrial NADPH [20]. Malic enzymes (MEs) catalyze the oxidative decarboxylation of L-malate and produce metabolites including NAD(P)H in the presence of divalent cations, among which ME3 presents the highest catalytic efficiency for NADP [21]. GLUD1 functions as a mitochondrial glutamate dehydrogenase that is strictly requires NADP+/NADPH as a cofactor to catalyze the conversion of L-glutamate into alpha-ketoglutarate [22]. The results above suggest that BAP1 may regulate NADP+/NADPH levels through transcriptional regulation of NADPH metabolism-related genes, yet to be further verified.
Fig. 4: Positive correlation between BAP1 and NADPH-related genes expression in KIRC patients.
A Expression level of 30 NADPH-related genes in patients with RCC between BAP1 low and high groups (data from TCGA-KIRC). B Heatmap of the expression levels of NADPH-related genes in BAP1 high and low groups. C Correlation analysis of BAP1 and NADPH-related genes. D Expression levels of significantly differentially expressed NADPH-related genes in UMRC6 cells.
The expression level of BAP1 is correlated with disulfidptosis-related genes (DRGs)Based on Liu’s publication [16], CRISPR/Cas9-mediated genome screening was utilized to investigate the genes involved in the regulation of disulfidptosis induced by glucose starvation. The top genes identified include GYS1, LRPPRC, NCKAP1, NDUFA11, NDUFS1, RPN1, and SLC3A2. Additionally, the paper identified genes expressing cysteine-containing peptides with disulfide bonds that increased at least 1.5-fold upon glucose starvation, as determined by high-performance liquid chromatography-mass spectrometry. The top genes in this category include ACTB, ACTN4, CAPZB, CD2AP, DSTN, FLNA, FLNB, INF2, IQGAP1, MYH10, MYH9, MYL6, PDLIM1, and TLN1. In total, these 21 genes were included in our study and termed disulfidptosis-related genes (DRGs). The expression levels of DRGs in the BAP1 high and low expression groups were shown in Fig. 5A, B. Of the 21 DRGS analyzed, except for NDUFA11, the remaining were positively correlated with BAP1 expression levels. Next, patients in TCGA-KIRC cohort were clustered into three clusters using a consensus clustering algorithm based on the expression data of 21 DRGs (Fig. 5C). In the Kaplan-Meier analysis of patients among the three groups, better survival outcomes were observed in cluster C1 patients (Fig. 5D). Besides, the expression of BAP1 in patients with C1 cluster was higher than that in the other two groups (Fig. 5E). These results reveal the positive correlation between the expressions of BAP1 and DRGs in patients with RCC. However, whether BAP1 regulates disulfidptosis through these DRGs remains further investigation.
Fig. 5: The expression level of BAP1 is correlated with disulfidptosis-related genes (DRGs).
A, B Expression level of 21 DRGs in patients with RCC between BAP1 low and high groups (data from TCGA-KIRC). C The CDF distribution diagram and Delta area diagram of consistency clustering analysis shows the optimal κ Value, and the DRG clusters when κ = 3. D The overall survival of patients with RCC varied among different DRG clusters. E BAP1 expression levels in the three DRG clusters.
Correlation analysis between BAP1 and DRGs expression in RCC patients with or without cancer-associated BAP1 mutationsAs a tumor suppress gene, BAP1 is involved in many physiological activities, such as normal DNA synthesis, interacting with transcription factors, deubiquitylation. There are more than 200 BAP1-mutant families, and individuals inheriting a BAP1 allele are tend to develop one or more malignancies during their lifetime [23]. We have revealed that BAP1 is positively correlated with the expression of DRGs in patients with RCC. Here, we want to further investigate whether there are differences in the expression of DRGs between RCC patients with BAP1 wild type (WT) and mutant (Mut), so as to clarify the influence of BAP1 mutations on DRGs. The pan-cancer analysis of the expression of BAP1 WT and Mut is exhibited in Fig. 6A. Of the 334 KIRC patients, 28 had a BAP1 mutation. Though, the expression level of BAP1 Mut was only slightly lower than that of WT. Patients with BAP1 Mut had worse survival outcomes than those with WT (Fig. 6B). We also compared the expressions of DRGs between BAP1 WT and Mut groups, and only the expressions of INF2 and FLNB were significantly different between the two groups (Fig. 6C). The waterfall diagram in Fig. 6D shows somatic mutations of BAP1 and DRGs, with the probability of BAP1 mutation up to 63.6%. In addition, 13 out of 21 DRGs existed mutation, and the mutation probability of LRPPRC, ATCN4, and IQGAP1 was relatively high. Together, the mutation status of BAP1 appears not to affect the levels of DRGs, indicating that BAP1 regulation of disulfidptosis might not be relevant to its tumor suppressive function.
Fig. 6: Correlation analysis between BAP1 and DRGs expression in RCC patients with or without cancer-associated BAP1 mutations.
A Expression of BAP1 WT and Mut across different tumor types. B The overall survival of patients with RCC between BAP1 WT and Mut groups. C Expression level of 21 DRGs in patients with RCC between BAP1 WT and Mut groups. D The waterfall diagram shows somatic mutations of BAP1 and DRGs.
留言 (0)