
記住我
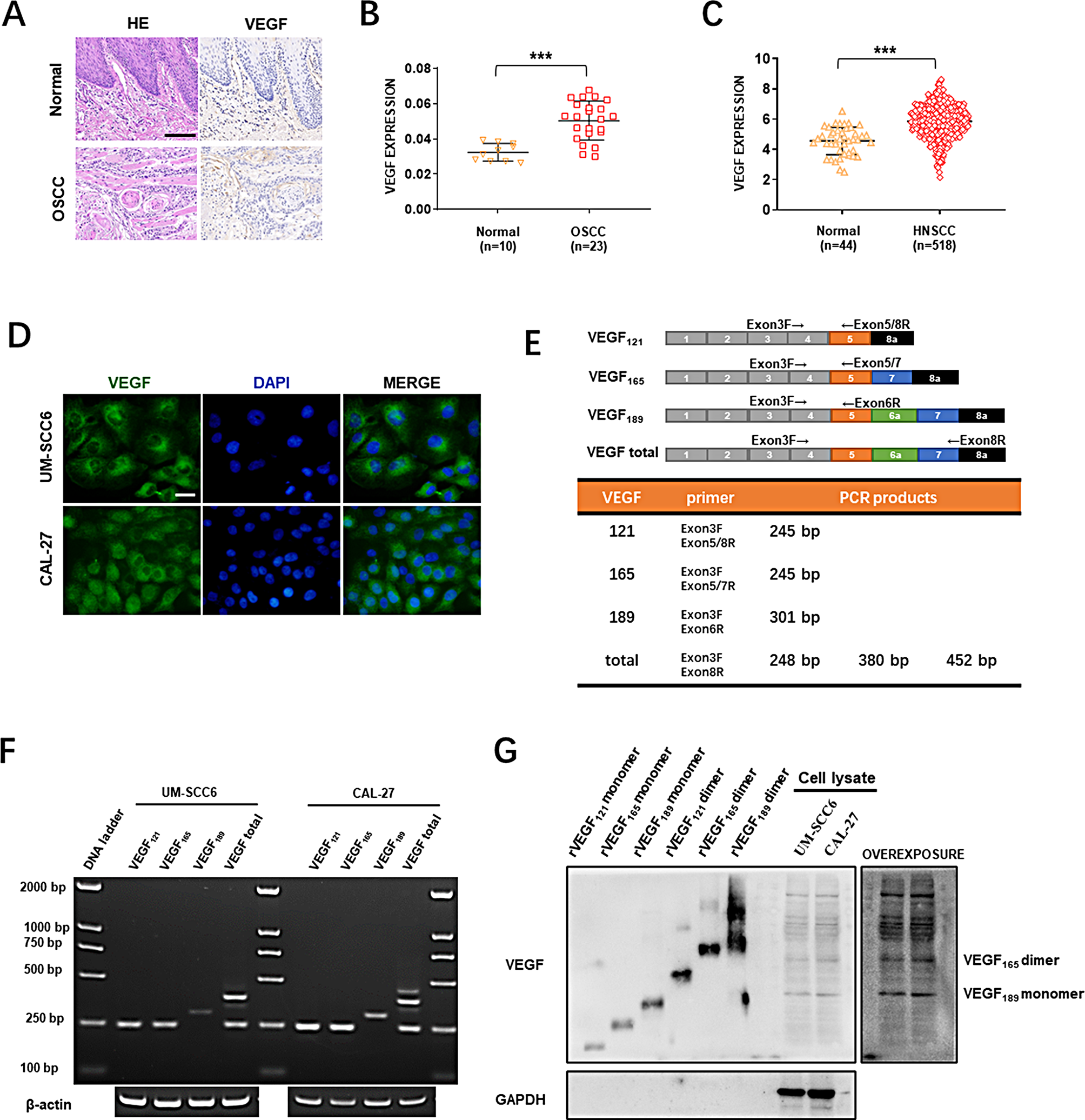



To define the function of circRNAs in the pathogenesis of GC, we collected preoperative plasma samples from three healthy individuals and three patients with GC to perform circRNA microarray analysis. Hierarchical cluster analysis revealed that hsa_circ_0006211 was the most upregulated circRNA in those plasma of patients with GC (Fig. 1A). We further examined the expression of these 15 circRNAs in 60 pairs of GC tumour tissues as well as in matched normal tissues and found that only hsa_cir_0006211 and hsa_cir_0050545 were expressed at higher levels in GC tissues than in paired adjacent normal tissues, and the difference in hsa_cir_0006211 expression was more significant (Fig. S1A). This indicated that hsa_circ_0006211 may play a critical role in GC progression, which was why we selected hsa_circ_0006211 for subsequent studies. We found that hsa_circ_0006211 was spliced from the UBA2 gene positioned at chr19:34,922,765–34,957,919 and formed a loop transcript of 1519 nt, thus, we named this circRNA as circUBA2. Sanger sequencing was performed to identify the splice site of circUBA2 (Fig. 1B). To determine whether the head-to-tail splicing of circUBA2 originates from trans-splicing or genomic rearrangements, we designed two types of primers, diverging primers for circUBA2 amplification and convergent primers for linear UBA2 mRNA amplification, and then extracted gDNA and cDNA from BGC823 cells. The results of gel electrophoresis revealed that circUBA2 was not detected in gDNA, but was detected in cDNA, showing that the specific circular structure of circUBA2 was indeed formed by reverse splicing (Fig. 1C). BGC823 and AGS cells were treated with RNase R to assess the stability of circUBA2. The results revealed that circUBA2 resisted RNase R digestion, whereas the linear structure of UBA2 is significantly degraded (Fig. 1D). The resistance of cirUBA2 to actinomycin D digestion further confirmed its loop structure (Fig. 1E). In addition, nuclear-cytoplasmic isolation and FISH assays illustrated that circUBA2 was predominantly localised in the cytoplasm (Fig. 1F, G). qRT-PCR analysis was conducted to assess the abundance of circUBA2 in eight GC cell lines (Fig. 1H). To assess the influence of cirUBA2 on GC, we examined its expression in 168 pairs of GC tumour tissues and matched normal tissues. The experiments revealed that compared to the corresponding adjacent normal tissues, GC tissues showed overexpression of circUBA2 in 75.60% (127/168) of tissues from the same patients (Fig. 1I). To further analyse the correlation between circUBA2 levels and clinicopathological characteristics, the tissues were split into two groups, a high circUBA2 group and a low circUBA2 group, according to the median circUBA2 expression. Analysis of pathological features suggested that circUBA2 expression was significantly correlated with tumour size, T stage, N stage, and TNM stage (Table 1). Furthermore, Kaplan–Meier analysis clearly illustrated that the survival rate of GC patients with high circUBA2 expression was lower than that of patients with low circUBA2 expression (p = 0.01) (Fig. 1J). In conclusion, circUBA2 was identified as a highly stable circRNA and a novel diagnostic and prognostic marker for GC.
Fig. 1
The expression, identification and prognostic significance of circUBA2 in GC. A Clustered heatmap of the top 15 differentially expressed circRNAs between 3 pairs of plasma specimens. B CircUBA2 (hsa_circ_0006211) is generated from the UBA2 gene that is located on chromosome 19, and the “head-to-tail” splicing sites of circUBA2 were confirmed by Sanger sequencing. C The presence of circUBA2 in BGC823 cells is verified by gel electrophoresis experiments. D qRT-PCR determining the abundance of circUBA2 and linear UBA2 in the BGC823 and AGS cell lines after RNase R treatment. E qRT-PCR determining the abundance of circUBA2 and linear UBA2 in the BGC823 and AGS cell lines after actinomycin D treatment. F Nucleoplasmic separation assays for the relative expression of circUBA2 in BGC823 and AGS cells. G FISH for circUBA2 localization in BGC823 cell. H The expression of circUBA2 in seven GC cell lines and normal gastric epithelial cells (GES-1) was quantified by qRT-PCR. I The T/N ratio of mRNA expression levels of circUBA2 in 168 paired GC samples after log-transformation (log 2). J The Kaplan–Meier method was used to analyse the overall survival rate of 168 patients. 168 patients were divided into two groups based on the median level of circUBA2 expression. All data were presented as mean ± SD. *p < 0.05; **p < 0.01; ***p < 0.001
CircUBA2 promotes the proliferation, migration and metastasis of GC cellsTo evaluate whether circUBA2 influences the function of GC cells, we transfected overexpressed circUBA2 or sh-circUBA2 lentiviruses into BGC823 and AGS cells and verified their efficiency by qRT-PCR. The results demonstrated a corresponding change in the expression of circUBA2, whereas the change in the expression of UBA2 was not statistically significant, confirming that BGC823 and AGS cells with steady overexpression or downregulation of circUBA2 were successfully generated (Fig. S1B). We investigated the effect of circUBA2 on cell proliferation using colony formation and CCK-8 assays. We found that the downregulation of circUBA2 inhibited the proliferation of BGC823 and AGS cells, whereas the overexpression of circUBA2 promoted cell proliferation (Fig. 2A, B). Flow cytometry assessment of the cell cycle revealed that downregulation of circUBA2 increased the proportion of cells in the G0/G1 phase but reduced the proportion of cells in the S phase in both BGC823 and AGS cells, whereas overexpression of circUBA2 resulted in the opposite trend (Fig. S1D). We further examined the expression of cell cycle proteins related to the G1/S transition, and the results suggested that downregulation of circUBA2 decreased the expression of cyclin D1, cyclin E1, and CDK2. By contrast, circUBA2 overexpression increased the expression of these proteins (Fig. S1C). These results showed that downregulation of circUBA2 resulted in cell cycle stagnation in the G0/G1 phase, resulting the in repression of GC cell proliferation. We also found that the downregulation of circUBA2 inhibited cell migration in vitro. Conversely, the upregulation of circUBA2 expression resulted in significantly enhanced mobility (Fig. 2C).
Fig. 2
CircUBA2 promotes the proliferation, migration, and metastasis of GC cells. A, B Colony formation and CCK-8 assays were performed in BGC823 and AGS cells transfected with the vector, circUBA2, control or circUBA2 shRNA. C A transwell assay was performed to evaluate the migratory capacity of BGC823 and AGS cells, scale bar = 200 μm. D Images of nude mice and tumours from the sh-NC and sh-circUBA2 groups (n = 5 mice per group). E Growth curves of xenograft tumours after 4 weeks. F Weight of the xenograft tumours after 4 weeks. G Representative images for in vivo images of lung metastasis models and quantitative fluorescence results (n = 5 mice per group). H, I Representative images and quantitative results of lung metastases and H&E staining in the sh-NC and sh-circUBA2 groups, scale bar = 50 μm. J Representative in vivo images of liver metastasis models and quantitative fluorescence results (n = 5 mice per group). K, L Representative images and quantitative results of liver metastases and H&E staining in the sh-NC and sh-circUBA2 groups, scale bar = 50 μm. *p < 0.05; **p < 0.01; ***p < 0.001
We explored the role of circUBA2 in GC cells in vivo and constructed a subcutaneous xenograft tumour model. After knocking down circUBA2, tumour weight was substantially reduced and the tumour growth rate was significantly lower compared to tumours in the sh-NC group (Fig. 2D–F). After the overexpression of circUBA2, these results were reversed (Fig. S1E-G). We successfully constructed a model of lung metastasis. Four weeks after injection, lung metastases were significantly reduced in the circUBA2 knockdown group of mice, as reflected by fluorescence imaging (Fig. 2G). Observation of the excised lung tissue revealed that the circUBA2 knockout group had fewer lung metastases than the control group (Fig. 3H). This result was supported by the H&E staining of the resected lung tissue (Fig. 3I). Furthermore, a liver-spleen metastasis model was established, which revealed that the downregulation of circUBA2 inhibited the liver metastasis capacity of BGC823 cells (Fig. 2J–L).
Fig. 3
CircUBA2 enhances the stem cell-like properties of GC cells. A Proportion of CD44-positive cells and statistical results for BGC823 cells. B Representative images and statistical result of sphere formation assay in BGC823 cells, scale bar = 200 μm. C IF staining for CD44 (red), NANOG (green), and DAPI (blue) in BGC823 cells. The percentages of CD44 and NANOG staining were quantified, scale bars = 50 μm. D GC tissues of human organoids were stably transfected with the vector, circUBA2, control, or sh-circUBA2 lentivirus, showing that overexpression of circUBA2 promoted organoid growth, whereas circUBA2 knockdown inhibited organoid growth. E IF staining for CD44 (red), NANOG (green), DAPI (blue) in human GC organoids. The percentages of CD44 and NANOG staining results were quantified, scale bars = 50 μm. *p < 0.05; **p < 0.01; ***p < 0.001
In summary, circUBA2 increased the proliferation, migration, and metastasis of GC cells.
CircUBA2 elevates stem cell-like properties in GC cellsAcquisition of cancer stem cell-like properties has been reported to be strongly related to the development, maintenance, and metastasis of GC [4]. To explore whether circUBA2 influenced the biological features of GC cells through the acquisition of CSC-like properties, we first investigated the correlation between circUBA2 expression and stem cell-like markers. qRT-PCR showed that circUBA2 increased the expression of CD44, NANOG, SOX2 and SOX9, but had no correlation with CD24, LGR5, OCT4, and CD133. These results were confirmed using western blotting (Fig. S2A-B). Flow cytometry indicated that the ratio of CD44-positive BGC823 cells to AGS cells decreased after circUBA2 knockdown, whereas these results were reversed when circUBA2 was overexpressed (Fig. 3A, Fig. S2C). Moreover, circUBA2 overexpression promoted the generation of human GC organoids and tumour spheres in BGC823 and AGS cells, respectively. This effect was reversed by circUBA2 knockdown (Fig. 3B, D; Fig. S2D). The transfection efficiency of circUBA2 overexpression or sh-circUBA2 in human GC organoids was verified by qRT-PCR (Fig. S2E). The percentage of CD44-NANOG-positive cells increased in the circUBA2 group but decreased in the inhibition group of circUBA2 (Fig. 3C, E, Fig. S3). We also confirmed the involvement of in the induction of self-renewal using limiting dilution assays in vitro and in vivo (Fig. S2F–K). Moreover, Immunohistochemical (IHC) staining assays of subcutaneous tumours in nude mice showed that the percentage of stem cell-like marker-positive cells was significantly higher in the circUBA2 group than in the control group; however, the opposite was observed in the inhibition group of circUBA2 (Fig. S4A–E). This result was confirmed by western blotting of the subcutaneous tumours (Fig. S4F). These observations indicate that circUBA2 enhances the stem cell-like properties of GC cells.
CircUBA2 affects the expression of STC1 by acting as a sponge for miR-144-5pWe first predicted potential differentially expressed target miRNAs using three databases (TCGA, CircBank, and Starbase) and obtained four miRNAs after taking intersections (Fig. 4A). qRT-PCR revealed that only miR-144-5p expression was significantly elevated in circUBA2-downregulated BGC823 and AGS cells (Fig. 4B). Furthemore, the expression of miR-4766-3p and miR-144-5p was downregulated in 90 pairs of GC samples, whereas miR-144-5p and circUBA2 expression was negatively correlated (r = 0.3069, p = 0.0033) (Fig. 4C, D, Fig. S5). Next, we found that luciferase activity was significantly altered in cells co-transfected with the miR-144-5p and circUBA2-wild-type (WT) reporter genes (Fig. 4E). Subsequently, FISH results showed that miR-144-5p and circUBA2 preferentially co-localised in the cytoplasm (Fig. 4F). The results of the anti-AGO2 RIP assay showed that AGO2-induced endogenous circUBA2 pulldown was specifically enriched in both BGC823 and AGS cells after miR-144-5p overexpression (Fig. 4G), confirming the direct binding of circUBA2 to miR-144-5p.
Fig. 4
CircUBA2 affects the expression of STC1 by acting as a sponge for miR-144-5p. A Venn diagram showing potential miRNAs in TCGA, Starbase, and circBank that bind to circUBA2. B Quantification of the expression levels of four overlapping miRNAs in control and circUBA2 knockdown BGC823 and AGS cells by qRT-PCR. C Correlation between circUBA2 and miR-144-5p expression in GC samples. D The mRNA expression of miR-144-5p in 90-paired GC tissues was determined by qRT-PCR. E The targeting relationship between circUBA2 and miR-144-5p in BGC823 and AGS cells was verified by luciferase activity assay. F FISH assay detecting the colocalisation of circUBA2 with miR-144-5p in the cytoplasm of BGC823 cells, scale bar = 50 μm. G RIP assay was used to detect the enrichment of circUBA2 and miR-144-5p in BGC823 and AGS cells. H A flowchart of the screening process for miR-144-5p downstream candidate genes. I Determination of mRNA expression levels of STC1 in miR-144-5p mimics-treated or inhibitor-treated and control BGC823 and AGS cells using qRT-PCR. J Western blotting of STC1 in BGC823 and AGS cells transfected with the indicated vectors. K The T/N ratio of mRNA expression levels of STC1 in 90 paired GC samples after log-transformation (log 2). L The correlation between miR-144-5p and STC1 expression in GC samples. M The correlation between circUBA2 and STC1 expression in GC samples. N The relationship between STC1 and miR-144-5p in BGC823 and AGS cells was verified using a luciferase activity assay. *p < 0.05; **p < 0.01; ***p < 0.001
Based on these results, we hypothesised that circUBA2 regulates GC malignancy and stemness by sponging miR-144-5p. To test this hypothesis, we immediately transfected miR-144-5p mimics into circUBA2 overexpressing cells, as well as miR-144-5p inhibitors into circUBA2-silenced BGC823 and AGS cells. The experimental results showed that miR-144-5p mimics impaired circUBA2-mediated promotion of GC cell proliferation and migration. Inhibitors of miR-144-5p reverse the inhibitory effects of circUBA2 suppression. These experiments included colony formation, transwell, and cell cycle assays (Fig. S6-7). Western blotting, sphere formation assays, and flow cytometry of CD44-positive cell also demonstrated that circUBA2 promoted stemness by sponging miR-144-5p (Fig. S8). These results illustrate that circUBA2 acts as a sponge for miR-144-5p, promoting cellular malignancy and cancer stemness in GC cells.
To identify the downstream targets of miR-144-5p, we analysed the TCGA and Starbase databases and screened six candidate genes with upregulated expression in TCGA (Fig. 4H). At the mRNA level, only STC1 expression was downregulated in miR-144-5p mimic-treated GC cells and was upregulated in miR-144-5p inhibitor-treated GC cells, and the expression of HOXA13, COL8A1, BCAT1, PDSS1, MTPAP was not statistically significant (Fig. 4I, Fig. S9A-E). Similar changes were observed in the protein expression level of STC1 (Fig. 4J). Moreover, STC1 expression was higher in 90 pairs of GC tissue samples than in healthy tissue samples, not only at the protein level (Fig. S9F-G) but also mRNA levels (Fig. 4K). The mRNA level of STC1 was visibly negatively correlated with miR-144-5p (Fig. 4L) and positively correlated with circUBA2 (Fig. 4M) in these samples. We also assessed the prognostic profile of STC1 in GC by IHC in another internal cohort. A distinct differencet in STC1 staining was observed between tumour and non-tumour tissue (Fig. S9H-I). STC1 expression was low in 78 (37.14%) samples and high in 132 (62.86%) samples. Combined with the survival information from these patients, the STC1 high expression group showed worse survival (Fig. S9J-K). To confirm whether STC1 has the same expression profile and prognosis in different populations, we evaluated patients using TCGA and obtained similar results (Fig. S9L-N). Rescue experiments showed that STC1 knockdown significantly reversed the promoting effects of the miR-144-5p inhibitor on GC cell proliferation and migration (Fig. S10). Dual-luciferase reporter assays demonstrated the binding of miR-144-5p to STC1 (Fig. 4N).
In summary, these results confirmed that circUBA2 regulates STC1 expression by sponging miR-144-5p.
CircUBA2 regulates the expression level of STC1 and promotes GC growth, metastasis and CSC-like propertiesTo determine whether circUBA2 exerted its biological effects by sponging miR-144-5p to affect STC1 expression, we suppressed STC1 expression in circUBA2-overexpressing cells. Sphere formation assays showed that STC1 downregulation decreased the sphere-forming ability of BGC823 and AGS cells, whereas overexpression of circUBA2 in STC1 downregulated cells partially restored this ability (Fig. 5A). Immunofluorescence (IF) experiments revealed a reduction in the percentage of CD44-positive or NANOG-positive cells after STC1 downregulation, and circUBA2 overexpression attenuated the effects of STC1 disruption (Fig. 5B). Similarly, flow cytometry revealed a decreased proportion of CD44-positive cells in the STC1-silenced group, which was reversed by circUBA2 overexpression (Fig. 5C). Based on further cytological assays, including colony formation, transwell assays, and cell cycle assays, circUBA2 overexpression also reversed the suppressive effect of STC1 repression on GC cell proliferation and migration (Fig. S11).
Fig. 5
CircUBA2 regulates the expression level of STC1 and promotes GC growth, metastasis, and CSC-like properties. A Representative images and statistical results of the sphere formation assay in BGC823 and AGS cells, scale bar = 200 μm. B IF staining for CD44 (red), NANOG (green), and DAPI (blue) in BGC823 cells. The percentages of CD44 and NANOG staining results were quantified, scale bars = 50 μm. C Proportion of CD44-positive BGC823 and AGS cells and statistical results. D Images of xenograft tumours from the four groups (n = 5 mice per group). E Growth curves of xenograft tumours after 4 weeks. F Weight of the xenograft tumours after 4 weeks. *p < 0.05; **p < 0.01; ***p < 0.001
To explore the roles of circUBA2 and STC1 in vivo, we also developed a xenograft tumour model and compared it to the group injected with control cells; xenograft tumours following the injection of STC1-silenced BGC823 cells showed a slower growth rate. However, circUBA2 overexpression partially reversed this effect (Fig. 5D–F). IHC staining of these xenograft tumours for previous stem cell-like markers, compared to the control xenograft tumours, showed that the percentage of CD44, NANOG, SOX9, SOX2 positive cells was reduced in the STC1-silenced group, and STC1 shRNA and circUBA2 co-transfection played a reversal role (Fig. S12). The results from the liver and lung metastasis models clearly indicated that circUBA2 elevated GC metastasis in vivo, similar to the effects of STC1 (Fig. S13).
In summary, circUBA2 regulates the expression level of STC1 and promotes GC growth, metastasis, and CSC-like properties.
CircUBA2/miR-144-5p/STC1 regulates IL-6 expression and activates JAK2/STAT3 signalingWe investigated the possible mechanisms through which circUBA2 regulates tumour progression and stemness in GC cells. Cytokine arrays were incubated with conditioned media from BGC823 or AGS cells to determine the relevant cytokines in the cancer cells. The results showed that in BGC823 cells, Interleukin-6 (IL-6), and leptin secretion were reduced and IL-8 secretion was increased in circUBA2-silenced cells relative to control cells. In circUBA2-silenced AGS cells, IL-6 and RANTES secretion was reduced (Fig. 6A, B). ELISA using an anti-IL-6 antibody showed that circUBA2 overexpression promoted IL-6 secretion, whereas knockdown of circUBA2 significantly inhibited IL-6 secretion. Moreover, circUBA2 overexpression was able to reverse the effects of sh-STC1 on IL-6 secretion (Fig. 6C). Changes in IL-6 mRNA levels were confirmed by qRT-PCR (Fig. 6D). Similar, results were obtained for the qRT-PCR of IL-6R and the ELISA of soluble IL-6R (sIL-6R) (Fig. S14A-D). In the TCGA database, both IL-6 and IL-6R expression positively correlated with STC1 (Fig. 6E). Based on the TCGA database, we chose gene set enrichment analysis (GSEA) to identify the pathways enriched in STC1 high-expressing tumours compared to those in STC1 low-expressing tumours. We identified ‘HALLMARK_IL6_JAK_STAT3_SIGNALING’ as one of the clearly upregulated pathways (Fig. 6F). This pathway was enriched in FMUUH_RNA-Seq 1 and FMUUH_RNA-Seq 2 (Fig. S14E-F). These above results suggested that the IL6/JAK2/STAT3 signaling may be a downstream pathway of the CircUBA2/miR-144-5p/STC1 axis.
Fig. 6
CircUBA2/miR-144-5p/STC1 regulates IL-6 expression and activates JAK2/STAT3 signaling. A Cytokine arrays were incubated with the supernatants of BGC823 and AGS cells transfected with knockdown circUBA2 or control lentivirus to identify differential cytokines. B Quantification of the experimental results for the cytokine arrays. C IL-6 expression in the supernatants of BGC823 and AGS cells was determined by ELISA. D Relative expression of IL-6 in stably transfected BGC823 and AGS cells was measured by qRT-PCR. E The correlation between STC1 and IL-6 or IL-6R according to TCGA’s GC samples. F Gene set enrichment analysis (GSEA) revealed a notable relationship between STC1 and ‘HALLMARK_IL6_JAK_STAT3_SIGNALING’ signaling pathway ( p = 0.002473). G, H Protein levels of IL-6/JAK2/STAT3 pathway members in stably transfected BGC823 and AGS cells were determined by Western blotting. I Protein levels of the IL-6/JAK2/STAT3 pathway members in BGC823 and AGS cells stably transfected with different doses of IL-6 were determined by western blotting. J Protein expression of gp130 and sIL-6R in the cell membrane and cytoplasm of sh-NC and sh-circUBA2 groups was detected by western blotting. K The interaction between gp130 and IL-6R in BGC823 and AGS cells was identified by co-immunoprecipitation (co-IP). *p < 0.05; **p < 0.01; ***p < 0.001
The IL6/JAK2/STAT3 signaling pathway is believed to be closely associated with GC development [24]. Western blotting showed that the total levels of JAK2 and STAT3 remained unchanged in circUBA2-overexpressing cells, whereas the levels of phosphorylated JAK2, STAT3, IL-6, Il-6R, and glycoprotein 130 (gp130) were significantly increased. The opposite result was observed in sh-circUBA2 cells (Fig. 6G). Rescue experiments with circUBA2 and STC1 yielded similar results (Fig. 6H). In addition, by treating circUBA2 knockdown or control cells with different doses of IL-6, we found that IL-6 promoted circUBA2-induced IL-6R, gp130, p-JAK2, and p-STAT3 expression (Fig. 6I). The IL-6 pathway is divided into classical IL-6 and trans-signaling pathways. Subcellular fractionation revealed that the levels of sIL-6R and membrane-bound gp130 decreased the most (Fig. 6J). In the co-immunoprecipitation assay, gp130 and IL-6R levels decreased in sh-circUBA2 cells (Fig. 6K). This suggests that circUBA2 knockdown induces fewer gp130 and IL-6R interactions in vivo. Therefore, we suggest that circUBA2/miR-144-5p/STC1 regulates the selective splicing of IL-6R mRNA or the proteolytic cleavage of IL-6R to amplify IL-6 trans-signaling via sIL-6R, but not through the enhancement of classical IL-6 signalling by IL-6R binding to the cell membrane.
In summary, CircUBA2/miR-144-5p/STC1 regulates IL-6 expression and activates JAK2/STAT3 signaling.
CircUBA2 activates IL-6/JAK2/STAT3 signaling to mediate GC stemnessWe performed rescue assays to ascertain the effects of circUBA and IL6 on CSCs proliferation. Tocilizumab (IL6R Ab) is an anti-human interleukin-6 receptor (IL-6R)-neutralising antibody that prevents binding between IL-6 and IL-6R, thus inhibiting both classical and trans signaling [25]. We added tocilizumab to cells transfected with sh-circUBA2 and control cells and observed that tocilizumab treatment further suppressed the mRNA and protein levels of CD44, NANOG, SOX2, and SOX9 in the sh-circUBA2 cell line (Fig. 7A–E). Similarly, tocilizumab further reduced the number of CD44-positive sh-circUBA2 cells (Fig. 7F). Moreover, we tested this hypothesis in BGC823 and AGS cells and human GC organoids and found that sh-circUBA2 and tocilizumab synergistically inhibited the growth of GC 3D cell spheroids and human GC organoids (Fig. 7G, H). Consequently, circUBA2 activates IL-6/JAK2/STAT3 signaling to mediate GC stemness.
Fig. 7
CircUBA2 activates IL-6/JAK2/STAT3 signaling to mediate GC stemness. A–D The mRNA levels of CD44, NANOG, SOX2, and SOX9 were quantified by qRT-PCR in tocilizumab-treated BGC823 and AGS cells. E Protein levels of CD44, NANOG, SOX2, and SOX9 in tocilizumab–treated BGC823 and AGS cells were quantified by western blotting. F Proportion of CD44-positive BGC823 and AGS cells and statistical results. G Stable transfection of knockdown circUBA2 and control lentivirus in spherical BGC823 and AGS cells treated with tocilizumab, showing that the knockdown of circUBA2 and tocilizumab synergistically inhibited spheroids growth, scale bar = 50 μm. H Stable transfection of knockdown circUBA2 and control lentivirus in human organoids further treated with tocilizumab, showing that the knockdown of circUBA2 and tocilizumab synergistically inhibited organoid growth, scale bar = 50 μm. *p < 0.05; **p < 0.01; ***p < 0.001
留言 (0)