
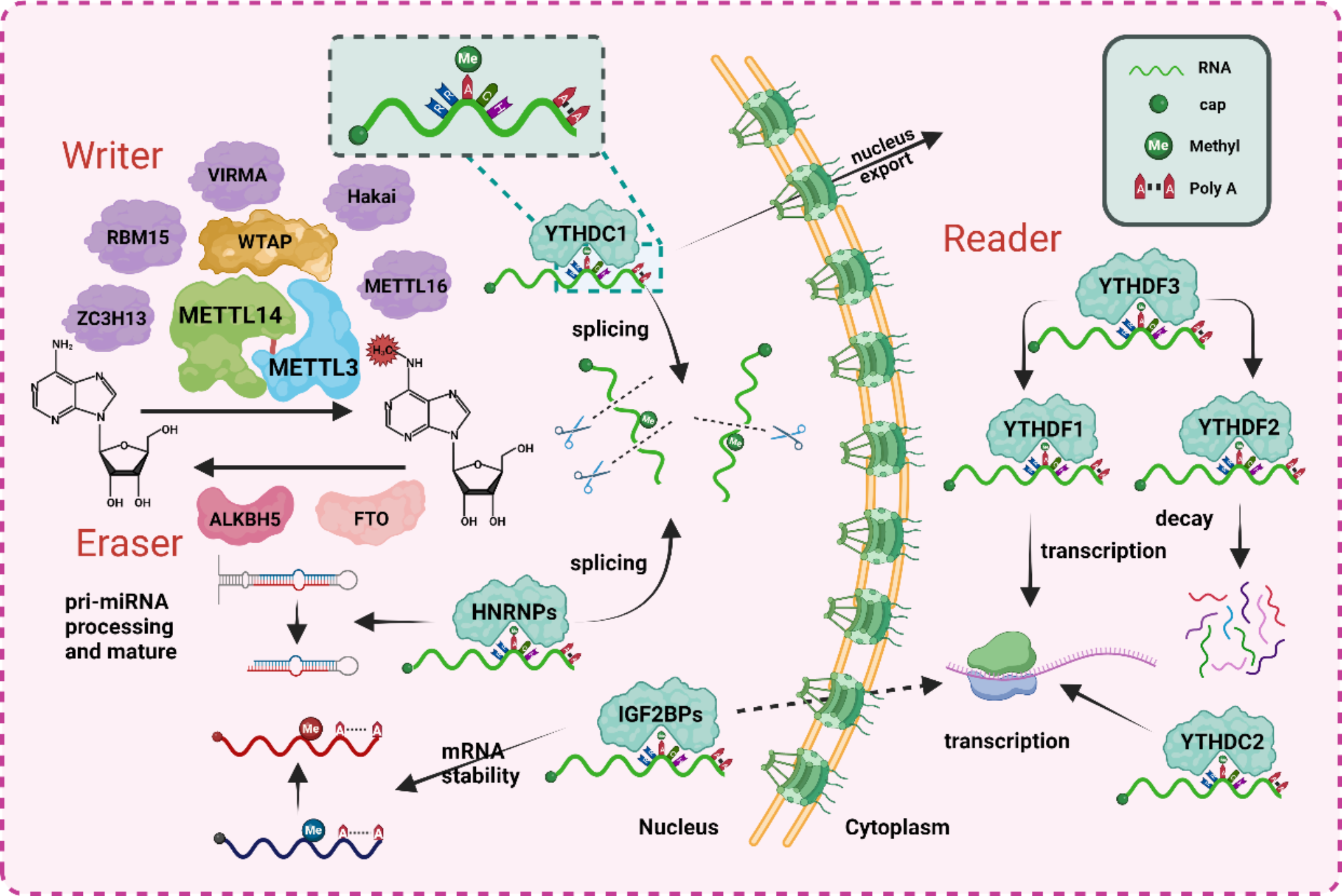
記住我



Using the Genotype Tissue Expression (GTEx) dataset and the Human Protein Atlas (HPA), we identified that ALDH5A1 has a considerably high expression in the human brain. Transcriptomic data from the GTEx dataset revealed that ALDH5A1 mRNA expression is quite high in the various parts of the human brain, including the brain cortex, which is a hotspot for GBM tumors (Figure S1A). The HPA transcript data of the brain also suggested that ALDH5A1 has consistently high expression (> 30 nTPM) in the different parts of the brain (Figure S1B). Furthermore, data from the HPA suggested that ALDH5A1 encoded protein SSADH is localized in the mitochondria (Figure S1C).
Fig. 1
ALDH5A1 is downregulated in GBM and associated with favourable patient survival: A–D ALDH5A1 mRNA expression in non-tumor (NT) vs. GBM tumor patient samples. E–G ALDH5A1 transcript levels in various grades of glioma. H ALDH5A1 mRNA expression in Indian patients. I ALDH5A1 transcript levels in GBM cell lines and normal brain RNA. J Representative images of immunohistochemical staining of glioma tissue samples with HPA029715 anti-ALDH5A1 antibody. Kaplan Meier survival curve of high and low ALDH5A1 expressing GBM patients K and Glioma patients L of the CGGA database. Data was downloaded from GlioVis web portal
After observing that ALDH5A1 has high expression in the human brain, we were next interested to know whether its expression is deregulated in GBM as compared to the normal brain. For this, we downloaded ALDH5A1 mRNA expression data from various brain-tumor datasets using the GlioVis web-tool. ALDH5A1 mRNA levels were found to be significantly downregulated in the GBM tissues of the TCGA GBM, Rembrandt, and Gravendeel datasets as compared to the non-tumor samples. On the other hand, in the Murat dataset, ALDH5A1 downregulation was found to be statistically insignificant (Fig. 1A–D). We also studied the grade-wise expression of ALDH5A1 in the glioma datasets: CGGA, Rembrandt, and Gravendeel. It was found that in all three datasets, ALDH5A1 mRNA expression decreased with an increase in tumor grade, grade IV having the lowest expression (Fig. 1E–G). We further measured the transcript levels of ALDH5A1 in a cohort of Indian GBM patients (control n = 3, GBM n = 33), and found it to be significantly downregulated in Indian patients too (Fig. 1H). Finally, we confirmed this by measuring ALDH5A1 transcript levels in the GBM cell lines U-87 MG, T98G and LN229 as compared to normal brain RNA. We found significant downregulation in all three cell lines (Fig. 1I).
We were further interested to know that whether this downregulation of ALDH5A1 mRNA levels in GBM is also corroborated at the protein level as well. For this, we studied the HPA where immunohistochemical analysis of ALDH5A1 was performed in a cohort of 9 glioma patients. None of the patient tissues had high staining for ALDH5A1, while two samples had medium and one sample had low staining. Six samples did not stain for ALDH5A1 at all (Figure S1D). Figure 1J shows one of each undetected, low and medium staining of high-grade glioma tissues using HPA029715 anti-ALDH5A1 antibody. This suggested that ALDH5A1 had relatively low protein expression in glioma patient samples.
To further understand the clinical relevance of ALDH5A1 in GBM, we downloaded GBM patient survival data along with ALDH5A1 expression from various brain-tumor datasets using the GlioVis tool. Kaplan meier survival analysis was performed and it was observed that high ALDH5A1 expression is associated with better OS of GBM patients of CGGA (Fig. 1K), Gravendeel (Figure S2A), and Vital (Figure S2B), datasets. However, no statistically significant association of ALDH5A1 expression and patient survival was observed in the TCGA GBM dataset (Figure S2C). Interestingly, we found that high ALDH5A1 showed a significant correlation to better survival in multiple glioma patient datasets (Fig. 1L, S2D, S2E).
ALDH5A1 is a direct target of miR-210 and both show inverse expression correlation in GBM patientsWe first investigated whether the downregulation of ALDH5A1 is due to genomic alterations such as mutations. We used the cBioPortal tool to study the alteration frequency of this gene in GBM patients. Among the three datasets studied, none had > 2% alteration frequency (Fig. 2A), suggesting the involvement of other possible regulatory mechanisms. Since miRNAs play a critical role as regulators of gene expression, we looked for possible miRNA binding sites in the 3′ UTR of ALDH5A1 transcript using the TargetScanHuman (release 8.0) miRNA target prediction tool. Among the miRNAs predicted to target ALDH5A1 3′ UTR with > 95 context + + score percentile, miR-210-3p was interesting as it has been reported earlier to be involved in mitochondrial dysfunction. We found a canonical binding site of miR-210 in the 3′ UTR of the ALDH5A1 transcript. The 2–8 nucleotides from the 5′ end of miR-210 (seed region) had complete complementarity with the ALDH5A1 3′ UTR region. TargetScanHuman (release 8.0) predicted this interaction to be an 8-mer having a high context + + percentile score of 97 (Figure S3A).
We then investigated the expression correlation of these two in GBM patients. A correlation analysis between miR-210 and ALDH5A1 mRNA levels in 358 TCGA GBM patients (Affymetrix HT HG U133A) revealed a statistically significant negative correlation between them (Pearson r = −0.253) (Fig. 2B). This suggested that miR-210 might regulate the expression levels of ALDH5A1 in GBM, and encouraged further validation studies.
Fig. 2
ALDH5A1 is a direct target of miR-210 in GBM: A Alteration frequency of ALDH5A1 in GBM patients. B Expression levels of ALDH5A1 and miR-210 are inversely correlated in GBM patients. C MiR-210 levels in GBM cells upon pre-miR-210-based overexpression. D ALDH5A1 mRNA levels in GBM cells upon pre-miR-210-based overexpression. E ALDH5A1 (SSADH) protein levels in GBM cells upon pre-miR-210 based overexpression. F MiR-210 levels in GBM cells upon anti-miR-based knockdown. G ALDH5A1 levels in GBM cells upon anti-miR-210-based knockdown. NC: Negative control. H MiR-210 binding site on the 3′ UTR of ALDH5A1 and mutation of binding site by SDM. I 3′ UTR-dual luciferase assay showing direct binding of miR-210 to the 3′ UTR of ALDH5A1. (*p > 0.01 and < 0.05, **p < 0.01, ***p < 0.001). Error bars denote ± standard deviation (SD)
To further establish the regulation of ALDH5A1 by miR-210 in GBM, we ectopically modulated expression of miR-210 in the GBM cell lines U-87 MG and T98G. We delivered the precursor sequence of miR-210 cloned in a mammalian expression vector (pBabe-Puro) to the GBM cell lines for overexpressing miR-210. Stem-loop RT-qPCR confirmed that upon transfecting the miR-210 constructs, the levels of mature miR-210 increased in both U-87 MG and T98G cell lines by > 3 folds (Fig. 2C). Upon upregulating miR-210, a significant decrease in ALDH5A1 transcript levels was observed in both cell lines (Fig. 2D). This effect was also verified at protein levels by performing immunoblotting for ALDH5A1 after miR-210 overexpression. We observed a downregulation of ALDH5A1 protein (SSADH, succinic semialdehyde dehydrogenase) in both U-87 MG and T98G cell lines upon miR-210 overexpression (Fig. 2E). Upon delivering anti-miR-210 to U-87 MG and T98G cell lines, we first ensured successful knockdown of endogenous miR-210 levels. By performing stem-loop RT-qPCR we confirmed that in both cell lines, the levels of miR-210 decreased significantly. A 65% downregulation of endogenous miR-210 levels was observed upon delivering anti-miR-210 to U-87 MG cells. On the other hand, in T98G cells this downregulation was > 95% (Fig. 2F). This suggested that anti-miR-210 based knockdown of miR-210 levels in GBM cells was successful. We further measured the effect of this on the transcript levels of ALDH5A1. By performing qRT-PCR, we found that miR-210 knockdown led to an increase of 1.71 and 1.61 folds in the transcript levels of ALDH5A1 in U-87 MG and T98G cells, respectively (Fig. 2G).
We finally verified the interaction between miR-210 and ALDH5A1 by performing a 3′ UTR dual luciferase assay. The 3′ UTR of ALDH5A1 containing miR-210 binding site was cloned downstream of a firefly luciferase gene in the pmiReport luciferase vector. We further mutated the binding site complementary to the miRNA seed region by changing 3 nucleotides (GCA->CGT) with the help of site-directed mutagenesis (SDM) (Fig. 2H). Upon co-transfection of the 3′ UTR construct along with miR-210 in the U-87 MG cell line, we observed a decrease in the luciferase activity (Fig. 2I). When the mutated 3′UTR was co-transfected to U-87 MG cells along with miR-210, a rescue in the luciferase activity was observed (Fig. 2I). This suggested that miR-210 mediated downregulation of ALDH5A1 is through its binding site in the 3′ UTR. Taken together, the above experiments established ALDH5A1 as a direct target of miR-210 in GBM.
Hypoxia regulates miR-210 and ALDH5A1 expression in GBMTo study if ALDH5A1 expression is regulated under hypoxic conditions, we first compared the transcript levels of ALDH5A1 with two major markers of hypoxia- VEGFA, and CA9 in GBM patients of the TCGA_GBM dataset using GlioVis web server. A statistically significant negative correlation was observed between ALDH5A1 and VEGFA/CA9 expression (Fig. 3A). Further, we checked its expression in various histological sections of GBM tumor. By exploring the IVY Glioblastoma Project (IVY-GAP) dataset, we observed that ALDH5A1 mRNA expression is significantly downregulated in the hypoxic pseudopalisades of GBM patients, when compared to the leading edge and cellular tumor (Fig. 3B). Finally, we measured the transcript and protein levels of ALDH5A1 under hypoxic conditions. We observed a significant downregulation of ALDH5A1 transcript levels in both U-87MG and T98G cell lines (Fig. 3C). Similar effects were observed at protein levels. A stark decrease in the protein levels of ALDH5A1 was observed in both U-87 MG and T98G cell lines when incubated under hypoxia (Fig. 3D). It is widely known that miR-210 is highly upregulated under hypoxic conditions as the transcription factor HIF1A induces its expression under hypoxia. Since GBM is a solid tumor having a hypoxic core, we verified this by culturing U-87 MG and T98G cells under chronic hypoxia (0.2% O2). When compared with normoxic conditions, it was found that miR-210 levels are elevated under hypoxia by > 2 folds in both cell lines (Fig. 3E). We further wanted to check if miR-210 upregulation under hypoxia has any effect on ALDH5A1 expression. For this, we knocked down miR-210 levels under chronic hypoxia and measured ALDH5A1 transcript levels. Interestingly, it was observed that repressing miR-210 under hypoxia using anti-miR-210 led to a significant increase in the transcript levels of ALDH5A1 (Fig. 3F).
Fig. 3
Effect of chronic hypoxia, epigenetic regulation and IDH mutation on ALDH5A1 and miR-210 levels: A Correlation of ALDH5A1 expression with hypoxia markers. B ALDH5A1 mRNA expression in different tissue sections of GBM. C ALDH5A1 is downregulated under hypoxia at the transcript level. D ALDH5A1 (SSADH) is downregulated under hypoxia at the protein level. E MiR-210 is upregulated under hypoxia in GBM. F MiR-210 knockdown under hypoxia increases the transcript levels of ALDH5A1. G 5-Azacytidine mediated hypomethylation increases ALDH5A1 transcript levels in a dose-wise manner. H ALDH5A1 is upregulated in IDH-mutant GBM cell lines. I MiR-210 is downregulated in IDH-mutant GBM cell lines. (*p > 0.01 and < 0.05, **p < 0.01, ***p < 0.001). Error bars denote ± standard deviation (SD)
Epigenetic regulation of ALDH5A1 and the effect of IDH mutationWe further wanted to study if ALDH5A1 is regulated by other epigenetic regulatory mechanisms apart from miRNA mediated post-transcriptional silencing. For this, we first investigated the presence of CpG islands in the promoter region of ALDH5A1. Using the Database of CpG islands and Analytical Tool (DBCAT), we identified a dense CpG island in the promoter region of ALDH5A1 (Figure S3B). In order to understand whether the CpG Island present in the promoter of ALDH5A1 is methylated, we treated the T98G GBM cells using increasing dosage of the global hypomethylation agent 5-Azacytidine and its vehicle control (DMSO). Interestingly, we found that with increasing dose of 5-Azacytidine, the transcript levels of ALDH5A1 increased (Fig. 3G). This suggested that ALDH5A1 downregulation in GBM might be attributed to epigenetic regulation of its expression by promoter methylation.
Although the 2021 WHO classification of CNS tumors has discarded the term ‘IDH mutant glioblastoma’, mutation in this gene has prognostic significance in glioma patients. Patients harboring an IDH1 R132H mutation have been observed to have a better prognosis than their wild type counterpart. The IDH1 (isocitrate dehydrogenase 1) is a crucial enzyme of the TCA cycle that converts isocitrate to α-ketoglutarate (α-KG), releasing NADPH in the process. The mutant IDH1 however converts α-KG into the oncometabolite D-2-hydroxyglutarate. Studies have suggested that R132H mutation promotes the uptake of lactate and glutamate instead of glucose to facilitate the TCA cycle. Since ALDH5A1 codes for the mitochondrial enzyme succinic semialdehyde dehydrogenase (SSADH) which converts succinate semialdehyde to succinate, feeding the TCA cycle, we were interested to check its expression in IDH mutant cells. For this, we overexpressed a wild type and R132H mutant clone of IDH1 into GBM cells and measured ALDH5A1 and miR-210 transcript levels. We found that ALDH5A1 is significantly upregulated in IDH mutant group of cells in both cell lines (Fig. 3H). Interestingly, we also observed that miR-210 is significantly downregulated in IDH mutant group of cells (Fig. 3I). An analysis of ALDH5A1 and MIR210 RNA in GBM patients of Bao dataset (data downloaded from GlioVis; IDH1 WT (n = 70); IDH Mut (n = 26) revealed ALDH5A1 to be upregulated (Figure S3C) and MIR210 to be downregulated (Figure S3D) in IDH1 mutant patients as compared to the wild-type.
Protein-protein Interaction and functional enrichment of ALDH5A1In order to understand the possible biological role of ALDH5A1 in GBM, we performed an in silico protein-protein interaction (PPI) study of ALDH5A1 using the STRING database. When a high confidence cutoff was applied, a total of 19 interacting proteins were identified that are mostly involved in cellular metabolism (Fig. 4A). Interestingly, several proteins of the isocitrate dehydrogenase (IDH) family such as IDH1, IDH2, IDH3A, IDH3B, and IDH3G, were found to be interacting with ALDH5A1. We further investigated how these interacting partners correlate clinically based on their expression. A correlation analysis in the GEPIA2 web portal revealed that these 19 genes had significant positive correlation with ALDH5A1 at the transcript level in GBM patients (Fig. 4B). To get a better look at the biological processes and pathways maintained by ALDH5A1 and its interacting proteins, we performed a gene ontology (GO) and pathway enrichment analysis of the above set of 20 proteins using the ShinyGO web-tool. Interestingly, we found that the most enriched molecular function (GO MF) was isocitrate dehydrogenase activity. Among the other enriched MFs, various dehydrogenase activities such as oxoglutarate dehydrogenase (succinyl-transferring) and L-malate dehydrogenase were observed (Fig. 4C). Isocitrate metabolic process, tricarboxylic acid metabolic process, and oxaloacetate metabolic process were among the top enriched biological processes (GO BP) (Fig. 4C). Among the top enriched cellular components (GO CC), oxoglutarate dehydrogenase complex, tricarboxylic acid cycle enzyme complex, and mitochondrial matrix were notable (Fig. 4C). Reactome, KEGG, and Panther pathway analysis revealed important pathways such as citrate cycle (TCA cycle), pyruvate metabolism, carbon metabolism and GABA synthesis release uptake and degradation, among others. Interestingly, TCA cycle and pyruvate metabolism were found to be common among all the pathways (Fig. 4D). Thus, we were interested in studying the possible role of ALDH5A1 in GBM metabolism. MiR-210 has been identified previously as a negative regulator of mitochondrial metabolism in other cancers. So, we were further interested to study the role of ALDH5A1/miR-210 axis in the context of GBM metabolism.
Fig. 4
In silico Protein-protein interaction and functional enrichment of ALDH5A1: A PPI network of ALDH5A1 and its interacting proteins (STRING database, confidence cutoff = high). B Correlation of ALDH5A1 transcript expression with its interacting partners in GBM patients (GEPIA2 database). C Gene ontology and D Pathway analysis of ALDH5A1 and interacting proteins. Data downloaded from ShinyGO web server
ALDH5A1 inhibits while miR-210 promotes aerobic glycolysis of GBM cellsPrevious reports have suggested that miR-210 inhibits mitochondrial respiration and might be indirectly involved in promoting the rate of glycolysis. Since ALDH5A1 is a mitochondrial enzyme and it was found to be a direct target of miR-210, we were further interested to investigate the role of miR-210/ALDH5A1 in aerobic glycolysis of GBM cells. We conducted a series of metabolic assays to investigate the role of ALDH5A1/miR-210 axis in GBM metabolism. Before proceeding with functional assays, ALDH5A1 overexpression was confirmed by immunoblotting where strong induction of ALDH5A1 protein was observed in the overexpression group as compared to the empty vector control (Figure S3E).
Fig. 5
Effect of ALDH5A1 and miR-210 overexpression on glycolysis: A Expression correlation analysis of ALDH5A1 with glucose transporters and glycolysis markers in GBM patients of Rembrandt dataset. Data was downloaded from GlioVis web server. B Flow-cytometric analysis of glucose uptake in GBM cells upon ALDH5A1 and miR-210 overexpression. C Effect of ALDH5A1 and D miR-210 overexpression on the transcript levels of glucose transporters and glycolysis markers. E Relative glucose uptake of GBM cells upon ALDH5A1 and miR-210 overexpression. F Relative lactate production of GBM cells upon ALDH5A1 and miR-210 overexpression. G Effect of ALDH5A1 and miR-210 overexpression on total ATP synthesis of GBM cells. (*p > 0.01 and < 0.05, **p < 0.01, ***p < 0.001). Error bars denote ± standard deviation (SD)
Glucose uptakeGlucose is the primary carbon source used for generating ATP, the energy currency of the cell. Cells that undergo rapid glycolytic flux to maintain high proliferation rate import more glucose through the glucose transporters. To measure glucose uptake, we first studied the expression correlation of two major glucose transporters in GBM- solute carrier family 2 member 1 (SLC2A1) and solute carrier family 2 member 3 (SLC2A3) (previously known as GLUT1 and GLUT3) with ALDH5A1 levels. By analyzing the transcriptomic data from the Rembrandt dataset, we found a significant negative correlation of ALDH5A1 mRNA levels with both SLC2A1 and SLC2A3, while the latter two shared a positive correlation between them (Fig. 5A). We further overexpressed both ALDH5A1 and miR-210 in the U-87 MG GBM cell line to study their effects on cellular glucose uptake. First, we performed a flow-cytometry based 2-[N-(7-nitrobenz-2-oxa-1,3-diazol-4-yl) amino]-2-deoxy-D-glucose (2-NBDG) uptake study. 2-NBDG functions as a fluorescent analog of glucose, employed for tracking glucose absorption in viable cells in real-time. Higher uptake of 2-NBDG in no-glucose media was observed more in miR-210 overexpressing group of cells (a 5.87% increase), while cells having ALDH5A1 overexpression showed a 9.51% decrease in the import of the compound as compared to their respective controls (Fig. 5B). Next, we measured the transcript levels of SLC2A1 and SLC2A3 upon ALDH5A1 and miR-210 overexpression. It was found that upon ALDH5A1 overexpression, the levels of SLC2A1 transcript got significantly repressed (Fig. 5C), while miR-210 overexpression led to an increase in the transcript levels of both SLC2A1 and SLC2A3 (Fig. 5D). Moreover, through a colorimetric biochemical assay that employs an improved o-toluidine method, we measured glucose uptake of cells overexpressed with ALDH5A1 and miR-210. It was found that miR-210 overexpression led to higher glucose uptake into U-87 MG cells, while ALDH5A1 overexpression resulted in opposite observation (Fig. 5E). This experiment finally suggested that ALDH5A1 overexpression reduces the uptake of glucose in cells while miR-210 overexpression promotes it.
Lactate productionIncreased lactate production is a direct indicator of highly glycolytic cells. For cancer cells which often have high glycolytic index, the pyruvate molecule is actively converted to lactate with the help of the lactate dehydrogenase (LDH) enzyme. On the other hand, pyruvate dehydrogenase (PDH), the enzyme responsible for converting pyruvate to acetyl-coA and thereby feeding the Krebs cycle, is inactivated by pyruvate dehydrogenase kinase (PDK) mediated phosphorylation. This helps in maintaining high glycolytic levels of actively proliferating cancer cells. We therefore focused on LDHA and PDK1; two very important enzymes that decide the fate of pyruvate and help decide whether cells will undergo glycolysis or OXPHOS. We first studied the expression correlation of LDHA and PDK1 with ALDH5A1 levels in GBM patients. By analyzing the transcriptomic data from the Rembrandt dataset, we found a significant negative correlation of ALDH5A1 mRNA levels with both LDHA and PDK1, while the latter two shared a positive correlation between them (Fig. 5A). Next, we overexpressed both ALDH5A1 and miR-210 in the U-87 MG GBM cell line to study their effects on the transcript levels of LDHA and PDK1. It was found that upon ALDH5A1 overexpression, the transcript levels of both LDHA and PDK1 got significantly repressed (Fig. 5C), while miR-210 overexpression led to an increase in the transcript levels of both LDHA and PDK1 (Fig. 5D). Cells export the generated lactate into extracellular space with the help of monocarboxylate transporters (MCTs) to avoid toxicity. This results in an increase in the extracellular lactate. We therefore measured lactate in the supernatant of miR-210 and ALDH5A1 overexpressed U-87 MG GBM cells. Interestingly, it was found that ALDH5A1 overexpression decreased the extracellular lactate formation while miR-210 overexpression increased it (Fig. 5F).
ATP synthesisAlthough the total ATP generated from the TCA cycle far exceeds than that of glycolysis, the oxidative phosphorylation process is rather slow, and aggressively proliferating cancer cells depend more on glycolysis for rapid ATP turnover. We therefore measured the total ATP production of GBM cells upon miR-210 and ALDH5A1 overexpression in a short time frame (24 h). It was observed that ALDH5A1 overexpression led to a marginal decrease in total ATP formation of both U-87 MG and T98G cell lines. On the other hand, miR-210 overexpression resulted in increased ATP formation of GBM cells (Fig. 5G).
Extracellular acidification rate (ECAR)Cellular glucose is transformed into pyruvate through glycolysis, and this pyruvate can then be further converted into lactate within the cell’s cytoplasm, or into carbon dioxide and water within the mitochondria. This process of glucose conversion, first to pyruvate and then possibly to lactate, leads to the release of protons, which are pushed out into the extracellular space. This proton release contributes to the increased acidity of the environment surrounding the cell. The XF instrument gauges this process by directly assessing the rate of acidification and presents it as ECAR (mpH/min), which is considered as the gold standard for assessing glycolytic rate of a living cell.
We measured the ECAR of GBM cells transfected with ALDH5A1 and miR-210, along with their respective controls. Interestingly, we found that the maximum ECAR was achieved in cells transfected with miR-210, while ALDH5A1 transfected cells exhibited minimum ECAR. On the other hand, the empty vector controls displayed similar ECAR profiles (Fig. 6A). Next we calculated the basal glycolysis (glycolysis), glycolytic capacity (Gly_cap) and glycolytic reserve (Gly_res), by analyzing the raw data. When ALDH5A1 was overexpressed, in U-87 MG GBM cells, we observed a significant reduction in the basal rate of glycolysis, glycolytic capacity as well as glycolytic reserve (Fig. 6B). On the other hand, overexpression of miR-210 in U-87 MG cells resulted in an increased glycolytic capacity and glycolytic reserve. However, we did not observe a significant change in the basal level of glycolysis (Fig. 6C). Taken together, the above experiment showed that ALDH5A1 reprograms GBM cells to undergo less glycolysis, while miR-210 can support higher glycolytic rates.
Fig. 6
Effect of ALDH5A1 and miR-210 overexpression on ECAR and OCR of GBM cells: A ECAR profile of U-87 MG GBM cells transfected with ALDH5A1 and miR-210 along with empty vector controls. B Measurement of glycolysis, glycolytic capacity and glycolytic reserve of control and ALDH5A1 overexpressed cells. C Measurement of glycolysis, glycolytic capacity and glycolytic reserve of control and miR-210 overexpressed cells. D OCR profile of U-87 MG GBM cells transfected with ALDH5A1 and miR-210 along with empty vector controls. E Measurement of basal, ATP-linked, maximal and spare respiration of control and ALDH5A1 overexpressed cells. F Measurement of basal, ATP-linked, maximal and spare respiration of control and miR-210 overexpressed cells. (*p > 0.01 and < 0.05, **p < 0.01, ***p < 0.001). Error bars denote ± standard deviation (SD)
Oxygen consumption rate (OCR)The OCR is a crucial parameter in cellular metabolism that reflects the rate at which cells utilize oxygen during various biochemical processes. OCR is primarily measured to gain insights into cellular respiration, a fundamental energy-producing mechanism in which cells use oxygen to generate ATP, the cell’s primary energy currency. By quantifying OCR, we can evaluate the efficiency of the OXPHOS and metabolic pathways within the mitochondria, offering valuable information about the cell’s overall energy status, metabolic health, and responses to different stimuli or conditions. Our previous findings indicated that ALDH5A1 reprograms cellular metabolism by reducing the glycolysis of GBM cells while miR-210 promoted it. Since ALDH5A1 codes for the mitochondrial SSADH which indirectly feeds the TCA cycle by providing succinate through the GABA-shunt, we were interested in studying the roles of ALDH5A1/miR-210 in mitochondrial respiration.
We measured the OCR of GBM cells transfected with ALDH5A1 and miR-210, along with their respective controls. Interestingly, we found that the maximum OCR was achieved in cells transfected with ALDH5A1, while miR-210 transfected cells exhibited minimum OCR. On the other hand, the empty vector controls displayed similar OCR profiles (Fig. 6D). Next, we calculated the OCR linked with basal respiration, ATP-linked respiration, Maximal respiration and the spare respiratory capacity, by analyzing the raw data. When ALDH5A1 was overexpressed, in U-87 MG GBM cells, we observed a significant increase in the basal, ATP-linked, maximal as well as spare respiratory capacity (Fig. 6E). On the other hand, overexpression of miR-210 in U-87 MG cells resulted in a decreased ATP-linked, maximum, and spare respiration. However, we did not observe a significant change in the basal level of respiration (Fig. 6F). Taken together, the above experiment showed that ALDH5A1 promotes the mitochondrial respiration of GBM cells, while miR-210 inhibits it.
ALDH5A1 inhibits cellular proliferation of GBMAs the functional role of miR-210 in GBM has been well reported [8,9,10,11,12], we were more interested in studying the functional role of ALDH5A1 in GBM. Since cellular proliferation is directly linked with the metabolism of cancer cells, we mainly studied the role of ALDH5A1 in regulating the proliferative capaity of GBM cells. We overexpressed ALDH5A1 in U-87 MG and T98G GBM cells and performed MTT-based cellular proliferation assay. It was observed that ALDH5A1 overexpression significantly inhibited the cell proliferation of both U-87 MG (Fig. 7A) and T98G cells (Fig. 7B) at day 4. The colony formation assay provides insights into the ability of cancer cells to survive, divide, and generate new colonies, reflecting their clonogenic potential and capacity for uncontrolled replication. We measured the colony forming ability of GBM cells after overexpressing ALDH5A1. It was observed that ALDH5A1 overexpression inhibited the clonogenic potential of both U-87 MG (Fig. 7C) and T98G (Fig. 7D) GBM cells. This finding supported our previous observation that ALDH5A1 overexpression reduced the glycolytic potential of GBM cells while promoting mitochondrial respiration. Reprogramming the primary mode of metabolism from glycolysis to OXPHOS might be responsible for the anti-proliferative effect of ALDH5A1.
ALDH5A1 decreases spheroid formation of GBM cellsThe 3D spheroid formation assay has emerged as a dynamic approach for assessing cancer cell proliferation within a more physiologically relevant context. When the cells are grown in the absence of a substratum in an anchorage independent manner, over time, they aggregate and self-assemble into compact spheroid structures, better representing the in vivo conditions compared to traditional two-dimensional monolayer cultures. The ability of cancer cells to form and grow as 3D spheroids is a measure of their proliferation and aggressiveness. We overexpressed ALDH5A1 in GBM cells and observed that ALDH5A1 overexpression reduced the spheroid forming ability of both U-87 MG (Fig. 7E) and T98G (Fig. 7F) cells. However, the decrease in spheroid diameter was more prominent in T98G cells as compared to U-87 MG.
ALDH5A1 induces G0/G1 cell cycle arrest in GBM cellsCell cycle analysis serves as a crucial method for quantifying cancer cell proliferation by examining the distribution of cells across different phases of the cell cycle. Accelerated proliferation in cancer cells often leads to alterations in the cell cycle distribution, such as a higher percentage of cells in S or M phases. We studied the effect of ALDH5A1 overexpression on the cell cycle of T98G GBM cells. It was observed that as compared to control (pC), approximately 8% more cells were arrested in the G0/G1 phase in ALDH5A1 transfected T98G GBM cells (Fig. 7G and H). This increase in cells in the G0/G1 phase might be due to anti-proliferative potential of ALDH5A1 in GBM.
Fig. 7
ALDH5A1 overexpression inhibits proliferation, spheroid formation and ROS levels in GBM: A ALDH5A1 inhibits cellular proliferation of U-87 MG cells. B ALDH5A1 inhibits cellular proliferation of T98G cells. Representative image of colony formation assay and quantification of colony numbers: C U-87 MG and D T98G cells. ALDH5A1 inhibits 3D spheroid formation and quantification of relative spheroid diameter: E U-87 MG and F T98G cells. G ALDH5A1 promotes G0/G1 cell cycle arrest of T98G cells. H Quantification of distribution of cells in various phases of the cell cycle upon ALDH5A1 overexpression. I ALDH5A1 inhibits ROS formation in GBM cells. (*p > 0.01 and < 0.05, **p < 0.01, ***p < 0.001). Error bars denote ± standard deviation (SD)
ALDH5A1 decreases reactive oxygen species (ROS) formation in GBMReactive oxygen species (ROS) derived from the partial reduction of oxygen molecule gives rise to free radicals that stimulate a broad spectrum of activity in cancer cells. ROS might lead to oncogene activation, and increased metabolism of cancer cells. Increased ROS initially promotes tumor progression, however after its excessive accumulation leads to cytotoxicity. Thus we were interested to understand the role of ALDH5A1 in the formation and management of cellular ROS. By using a spectroscopic method, we measured the ROS levels of GBM cells upon ALDH5A1 overexpression. Interestingly, we found a significant decrease in the ROS levels of cells having high levels of ALDH5A1. A spectroscopic analysis revealed that ALDH5A1 overexpression reduces ROS formation in both U-87 MG and T98G (Fig. 7I) cells. This might be due to the fact that ALDH5A1 metabolizes the toxic aldehydes in cells that contribute to higher ROS levels. Thus, ALDH5A1 acts as an anti-oxidant molecule in GBM.
Fig. 8
Schematic diagram summarizing the role of miR-210/ALDH5A1 axis in GBM metabolism
留言 (0)