In this study, extra-islet insulin- or glucagon-positive cells in donors with, and without T1D, have been examined. In concordance with previous studies [1, 4, 5, 12, 13], we report that insulin-positive extra-islet cells are still present even after a long duration of T1D. The reduction of insulin-expressing extra-islet cells was seemingly less pronounced than the loss of beta cells within islets, as extra-islet insulin-positive cells were found in all but one T1D donor. This implies that the extra-islet insulin-positive cells has a capacity to be replenished, and/or that they have a higher resistance to the events causing the beta-cell loss in T1D. Although the extra-islet insulin-positive cells were often present, they were reduced compared to donors without diabetes. This could be a consequence of the same beta-cell destructive process(es) causing the primary demise of islet beta cells in T1D or secondary to hyperglycemia-associated glucose toxicity [4, 5]. In this context, the three donors with remaining ICI in some lobes are of particular interest. A more overarching pathological mechanism, such as glucose toxicity, would likely cause a widespread and equal demise of beta cells in the pancreas. However, in all three donors, there were 2.5 to 15 times more extra-islet insulin-positive cells in the areas where ICI was present, compared with in the areas with IDI. This argues against a system-wide pathological mechanism of beta cell demise in T1D, and for a selective one that affects most, but not all, lobes.
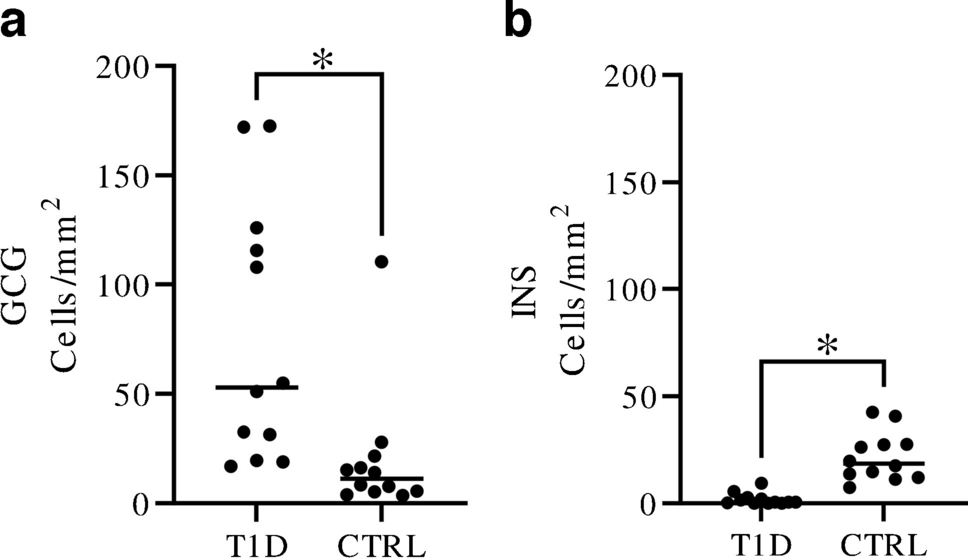
The increased frequency of extra-islet glucagon-positive cells in T1D is noteworthy, and the frequency is considerably higher compared to what has been previously reported for islets (median 53.0 extra-islet cells/mm2 in T1D, median 11.2 extra-islet cells/mm2 in controls, and median 4.4 islets/mm2 in control subjects) [14]. Importantly, the increased frequency was maintained even when glucagon-positive cells in the peri-islet and peri-ductal areas were excluded (data not shown), confirming that the increased glucagon frequency was not merely an effect of, e.g., shattered islets. There are several possible explanations for the increased number of glucagon-positive cells. The reduced pancreas size observed in T1D [15, 16], possibly resulting from loss of acinar but not endocrine cells in the exocrine parenchyma, may contribute to the observed increased frequency of glucagon-positive cells but is unlikely to explain the entire increase that was almost fivefold. Islet alpha cells have been shown to have a reduced function in T1D, with impaired glucagon secretion and altered gene expression [17]. The increase of glucagon-positive cells scattered in the exocrine parenchyma could be a compensatory response to counterbalance the impaired alpha-cell function.
Conversion of alpha cells into beta-cells upon GABA exposure, as a form of beta-cell neogenesis, has been debated but elegantly shown in a mouse model as well as in human islets transplanted into mice [18, 19]. This, in turn, triggered an alpha cell replacement mechanism through neogenesis from ductal precursor cells [18]. The increased number of glucagon-positive extra-islet cells observed herein could suggest that this mechanism of alpha-cell replacement is functional in donors with T1D. Conversion of human alpha-cells into beta-cells has been observed in vitro as well [20]. Furthermore, in this study, we observed a lack of transcription-factor expression in many cells, suggesting that they could be immature, newly formed or plastic, in line with the idea of neogenesis and/or conversion of cells. An alternative hypothesis would be that the increased number of glucagon- and decreased number of insulin-positive extra-islet cells are the result of trans-differentiation of beta cells to alpha cells as shown in experimental studies and in T2D [21,22,23]. However, the mechanism of conversion has also been questioned [19]. In line with this, intermediate phenotypes such as cells co-positive for glucagon and PDX1, which have been described in the context of in vitro transdifferentiation [20], were very rarely found in this study.
Several mechanisms for the expansion of beta-cell mass in non-diabetic settings have been proposed [24,25,26,27]. In the current study, we report a mitotic activity of between 0 and 3% in both donor groups, defined as Ki67-positive expression in the nuclei. However, only 200 cells were examined per donor, and out of these a very limited number of cells showed mitotic activity, adding a level of uncertainty to the results. Nevertheless, the observed mitotic activity is high in comparison to what have been reported for islets [28,29,30], suggesting the extra-islet cells could play a role in the endocrine cell expansion.
In the current study, we characterized the extra-islet endocrine cells in T1D by examining well-preserved pancreatic tissue obtained from heart-beating organ donors previously diagnosed with T1D. However, the sections were only analyzed in 2-D which has the possible consequence that some annotated single cells could be part of the outermost border of an islet. Yet, in a previous study on extra-islet single cells, this possibility was addressed by examining consecutive sections, and it was concluded that no single cells were part of an islet [31]. An additional constraint of the study is that only a limited area of each pancreas was investigated, as well as relatively few cells. However, samples from only the body/tail of the pancreas were deliberately chosen to avoid confounding factors from the islets originating from the uncinate process and the head of the pancreas. Despite the above mentioned limitations, this study represents, to the best of our knowledge, the largest investigation to date conducted on extra-islet cells in T1D.
In summary, we provide a characterization of extra-islet cells expressing insulin or glucagon in donors with or without T1D. Results presented suggest that the pathological mechanisms involved in T1D not only affect islets regionally, but also the extra-islet beta-cells in affected lobes of the pancreas. Surprisingly and of high interest, we also present an increase in the frequency of extra-islet glucagon-positive cells in donors with longstanding T1D. The presence of extra-islet insulin- and glucagon-positive cells with mitotic activity suggests preserved renewal of endocrine cells in donors both with and without T1D.







留言 (0)