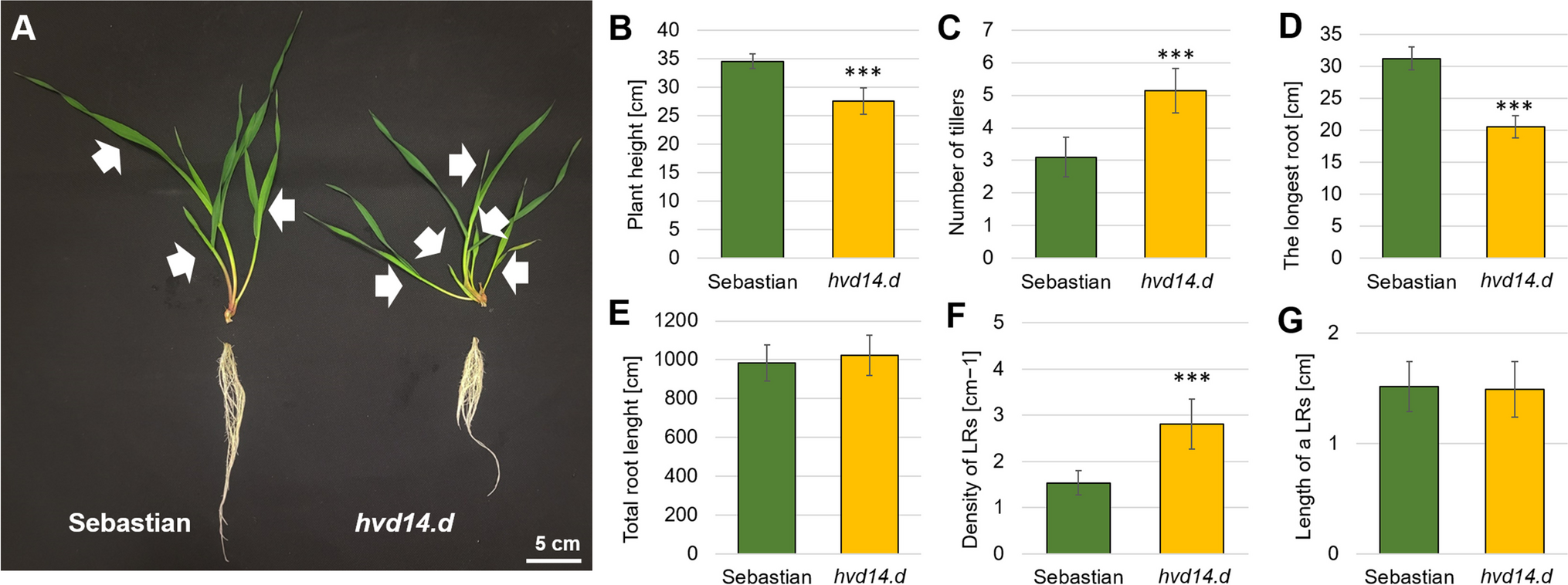
SL insensitivity impacts the development of barley shoot and root under hydroponic conditions
SLs are known mediators of shoot and root development, crucial in plant adaptation to environmental conditions. Photoperiod and temperature (Djennane et al. 2014), light intensity and length (Jia et al. 2014), and above all, nutrient availability (Yoneyama et al. 2013) affect SL biosynthesis/signalling, which influences plant development. Thus, the phenotype of SL mutants may vary depending on growing conditions. Here, we grew barley SL-insensitive mutant hvd14.d for the first time under hydroponic conditions. Previous analyses carried out on plants anchored in the soil or vermiculite revealed that hvd14.d produces twice as many tillers as WT. However, those differences were less pronounced in the first stages of plant development, i.e. 3-week-old hvd14.d plants grown in the soil produce 50% more tillers than Sebastian (3 ± 0.39 and 2 ± 0.21, respectively) (Marzec et al. 2016). Hydroponic culture in ½ Hoagland solution (Hothem et al. 2003) increased the number of tillers in both genotypes at the same level. Still, the differences between genotypes are similar, and a 60% higher number of tillers was observed in hvd14.d (Fig. 1). Plant height was the second feature differentiating both genotypes. The height of mutant hvd14.d grown in soil and hydroponics was reduced by about 20%. However, both genotypes were higher when grown in hydroponics (Sebastian: 34.5 ± 1.25 vs 29.1 ± 3.7 cm; hvd14.d: 27.5 ± 2.32 vs 22.4 ± 4.1 cm) (Marzec et al. 2016) (Fig. 1). It was previously reported that hydroponic conditions promote shoot development when compared to plants sown in soil (Dutta et al. 2023) that may be caused by easy access to water and nutrients in a hydroponic medium throughout development.
Under control conditions, SLs shape root architecture by promoting root elongation and inhibiting lateral root development (Kapulnik et al. 2011; Ruyter-Spira et al. 2011). Similar results were obtained when hvd14.d and Sebastian were grown in a solid medium (vermiculite) and watered with ½ MS medium. The mutant exhibited shorter seminal roots in a solid medium that produced more lateral branches than Sebastian (Marzec et al. 2016). Under hydroponic conditions, the same differences were observed (Fig. 1). In contrast, both genotypes in soil and hydroponic developed the same number of seminal roots, but their length was reduced, which is a well-known effect of hydroponics (Mian et al. 1993). Root elongation in the mutant was reduced by 35% under both conditions, hydroponic: 31.2 ± 1.79 vs 20.5 ± 1.74 cm and soil 47.1 ± 2.51 vs 34.8 ± 1.94 cm, respectively for Sebastian and hvd14.d (Marzec et al. 2016) (Fig. 1). Differences in lateral root density between genotypes were independent of growing conditions and hvd14.d developed a higher number of lateral roots per 1 cm of seminal root (Marzec et al. 2016) (Fig. 1). Obtained results indicate that growing conditions do not affect shoot and root differences between hvd14.d and its WT, as far as plants were supplemented with an optimal nutrient concentration.
Tissue-specific transcriptome alterations caused by SL insensitivity
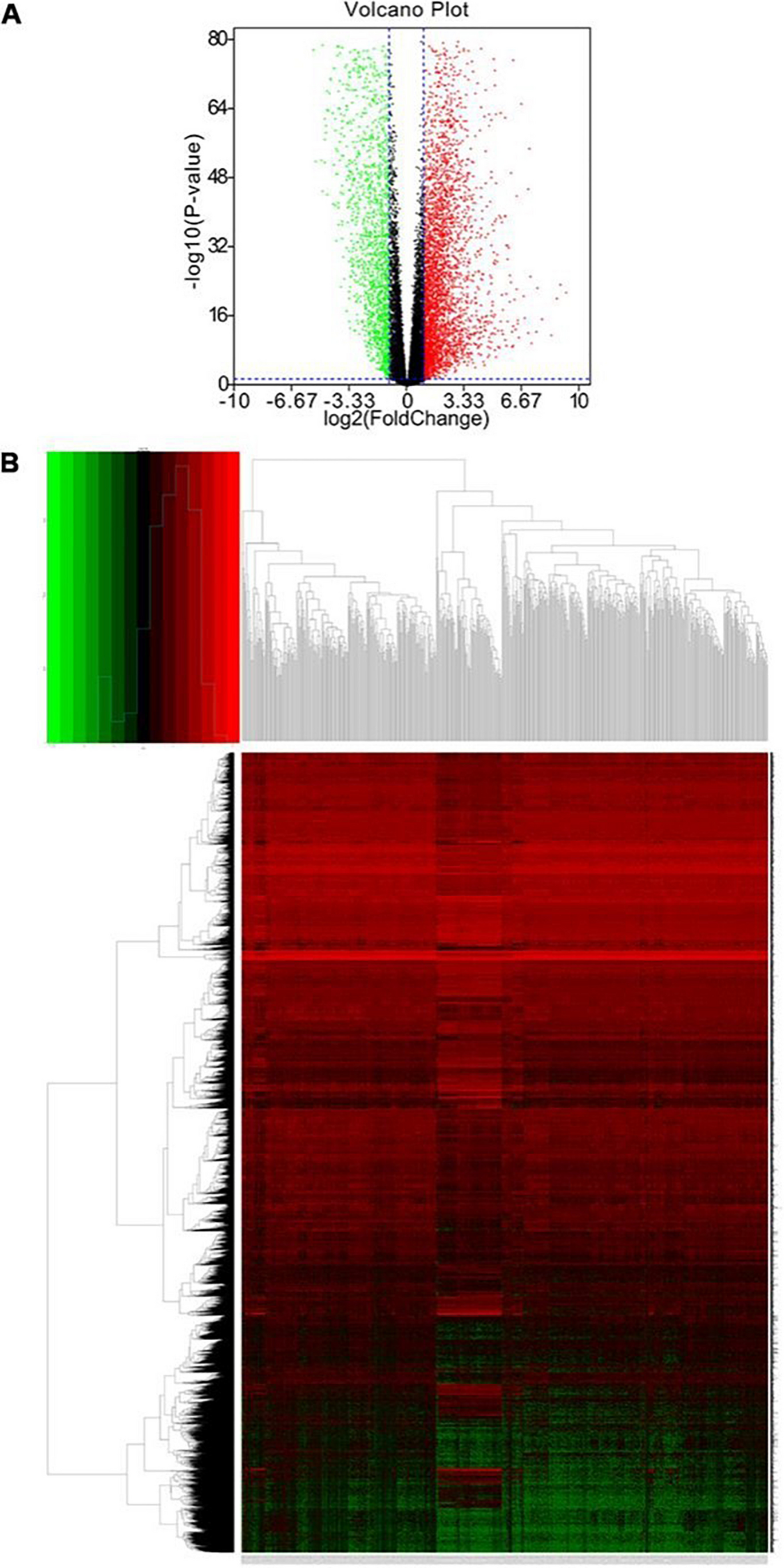
Because mutation in barley SL receptor HvD14 affects shoot and root development, those differences become statistically significant at the third week of plant development (Fig. 1). The transcriptome of shoot and root was compared between hvd14.d and Sebastian, revealing 6702 DEGs. Up to 80% of all identified DEGs were found in roots (5414 among 6702). In both organs, a more significant number of transcripts, around 60%, was downregulated (Fig. 2). Among all identified DEGs, only 3% (222) were found in both shoot and root comparisons. The largest category, up to 77% of all DEGs, was root-specific. These results indicate that SLs play a more pronounced role in root development relative to shoots in 3-week-old barley plants. It might also be related to the higher complexity of the root system at this stage of plant development, such as a large number of different cell types supporting vastly different transcriptional programs as is the case in Arabidopsis (Shahan et al. 2022). A 3-week-old barley seedling’s root system comprises seven seminal and hundreds of lateral roots at the different stages of development: initiation, elongation or branching. On the other hand, the shoot of a 3-week-old barley plant has from three to five tillers at the vegetative stage of development, which do not have developed internodes. Hence, more genes are involved in the development of the root, compared to the shoot, at this stage of plant growth. To date, there are no comparative studies about shoot and root transcriptomes for SL mutants, let alone a major crop species. Global analyses of transcriptome alterations caused by SL treatment or mutation in SL biosynthesis/signalling pathways were investigated for whole plants (Wang et al. 2020) or shoot/root separately (Zhan et al. 2018; Asghar et al. 2022; Haider et al. 2023; Li et al. 2023).
SL-responsive genes found in barley shoot and root
In total, 222 DEGs were common between the shoot and root of hvd14.d compared to Sebastian. Among the 65 upregulated DEGs, 25% (16) were involved in translation along with others. Moreover, DEGs involved in ribosome biogenesis (4) or RNA processing (4) were found to be upregulated in hvd14.d (Supplementary Data 2). It was shown that SLs increase the cold tolerance of Brassica rapa L. seedlings because GR24 (a synthetic analogue of SL) treatment alleviates the damage of low-temperature stress (Zhang et al. 2020a, b). Within DEG between plants pretreated with spraying 0.1 μmol‧L−1 GR24 and non-pretreated, exposed to low temperature (4 °C), the genes encoding translation initiation factors were downregulated. On the other hand, in pea, removing apical meristem promotes the outgrowth of axillary buds, which was linked with increased expression (up to 35-fold) of gene encoding ribosomal protein (Stafstrom and Sussex 1992). The conclusion that SLs affect the translation processes via control of ribosome complex activity can be postulated. However, it cannot be excluded that stronger activity of the translational process observed in hvd14.d is related to the higher number of developing tillers and lateral roots, and those processes are associated with rapid protein synthesis. Hence, changes in expression of translation-related genes are not a direct result of SL activity, but the effect of SL insensitivity, resulting in the development of more meristems.
Surprisingly, both hvd14.d organs showed reduced gene expression related to photosynthesis and plastids (Supplementary Data 2). Changes in the expression of photosynthetic genes in non-green tissue, including roots have been widely reported for various species under different stresses, such as drought (Molina et al. 2008; Cohen et al. 2010; Ranjan and Sawant 2015; Janiak et al. 2019) or phosphate starvation (Wu et al. 2003; Li et al. 2010). It was shown that the suppression of photosynthetic genes is required for sustained root growth of Arabidopsis exposed to phosphorus deficit (Kang et al. 2014). Reduction in photosynthetic genes in roots during stress responses is also linked with decreased production of reactive oxidant species (ROS) (Kang et al. 2014; Janiak et al. 2019). Our previous analyses indicated that hvd14.d exhibits reduced ROS scavenging under drought (Daszkowska-Golec et al. 2023). Because SL treatment seems to decrease ROS content in various species (Trasoletti et al. 2022), including barley exposed to cadmium (Qiu et al. 2021), we may speculate that SL-insensitivity of hvd14.d results in less efficient ROS scavenging. Thus, to reduce ROS production, the mechanisms related to photosynthesis are repressed in SL-insensitive barley mutant under control conditions. In fact, study investigating the effect of SL on photosynthesis confirm these predictions. Treatment with 1 and 5 µM of GR24 increased the net photosynthesis rate (µmol CO2·m−2·S−1) of salt stressed rice seedlings to values observed in control plants (Ling et al. 2020). Further, in cucumber (Cucumis sativus L.), greater photosynthetic efficiency was observed in GR24-pretreated plants than in non-GR24-pretreated plants under salt stress (Zhang et al. 2022). Under control conditions, the foliar application of GR24 on Artemisia annua increased various attributes related to photosynthesis (chlorophyll fluorescence, internal CO2, and net photosynthetic rate) as well as activity of photosynthetic enzymes (carbonic anhydrase, nitrate reductase, RuBisCO) (Wani et al. 2023). The general positive role of SL on photosynthesis was well documented, so the decreased expression of photosynthesis-related genes in the shoot of SL-insensitive hvd14.d confirms these results. Conversely, repression of those genes in roots may be linked with reduction of processes linked to ROS production.
Shoot- and root-specific SL-responsive genes
Within upregulated DEGs described as specific for shoot tissue, the largest group among the enriched GO terms was protein phosphorylation (Supplementary Data 2). Phosphorylation is one of the main post-translational modifications that affect protein interactions and stability, hence has a significant impact on gene expression, signalling pathways and enzyme activity (Khalili et al. 2022). Chen and co-workers indicated that GR24 treatment of rice SL-biosynthesis mutant (d10) changed the phosphorylation status of 8 proteins at a conserved phosphorylation site (Chen et al. 2014). Upregulated DEGs in hvd14.d involved in phosphorylation suggest that SLs may repress phosphorylation in barley shoots. On the other hand, among downregulated shoot DEGs, the large group was annotated as related to the cell wall organization and biogenesis, cell wall polysaccharide metabolic processes or polysaccharide biosynthetic and metabolic processes (Supplementary Data 2). There is a known role for SLs in promoting secondary cell wall formation in cotton (Gossypium hirsutum) where exogenous GR24 increased, and the application of SL biosynthetic inhibitor (Tis108) reduced the thickness of the secondary cell wall (Wen et al. 2023). Moreover, SL biosynthesis genes (MAX3 and MAX4) have been linked with xylan and cellulose deposition in Arabidopsis (Ramírez and Pauly 2019). Further, we previously reported the alteration of cell wall formation in hvd14.d in response to drought (Marzec et al. 2020). Interestingly, this is a conditional phenomenon as under control conditions; there are no differences in the cell wall thickness in the leaves of 3-week-old seedlings of hvd14.d and Sebastian (Marzec et al. 2020); however, there have been no investigations into the chemical composition of the cell wall to date. Thus, the differences in the polymer content between hvd14.d and Sebastian cannot be excluded. Secondary cell walls contain mainly cellulose, xylans and lignin, but their proportions and modifications depend on the functional needs of cell/tissue and, thus may vary between leaves and roots (Kumar et al. 2016). The data obtained, where decreased expression of genes related to cell wall biosynthesis was found specifically in barley shoot (Supplementary Data 2), narrowed down the possible role of SLs in the biosynthesis of cell wall components to that characteristic for shoot.
Within SL-related upregulated DEG found in roots, a significant number was annotated as cell cycle or cell cycle processes. It could be explained by the larger number of developing roots, thus the higher number of fast-dividing meristems in hvd14.d compared to the Sebastian (Fig. 1). On the other hand, both up and downregulated DEGs were annotated as involved in responses to abiotic stresses, stimuli and chemical or oxygen-containing compounds. Because SLs play an important role in plant adaptation to stresses, the insensitivity to SLs may disturb the multiple pathways related to the plant’s stress responses.
SL-dependent TFs
General knowledge of the SL signalling pathway and the individual proteins involved in signal transduction is well established in model species such as Arabidopsis or rice, from the SL signal perception to the degradation of the SL repressor (Marzec and Brewer 2019). However, we still have rudimentary information about the transcriptional responses in crops and non-model plants. Particularly the TFs that regulate the plant’s response to SLs. Here, by simultaneously comparing changes in the shoot and root transcriptome of hvd14.d and Sebastian, we proposed a set of TFs that may play a role in SL signal transduction in barley and which are involved in phenotypic changes observed in the shoot and root architecture of 3-week-old plants described above. In total, 28 TFs were identified as putative SL-related TFs as they (1) exhibit changed expression in hvd14.d versus Sebastian, (2) they are proposed to recognize binding sites in promoters of a multitude of identified DEGs and (3) motifs recognized by those TFs are over-represented (p-value ≤ 0.05) in DEG promoters (Table 1, Supplementary Data 9).
Interestingly, no one TF was differentially expressed in shoot and root barley tissue (SL_C), indicating differences exist in SL signal transduction between these two organs. Four TFs were previously identified as putatively involved in mediated SL-dependent barley response to drought (Daszkowska, 2023) (Supplementary Fig. 2). Two of these HORVU.MOREX.r2.6HG0471210.1 (AT1G80840) and HORVU.MOREX.r2.1HG0074290.1 (AT2G46270) are involved in plant response to abscisic acid (ABA) and were found to be upregulated by drought only in the Sebastian shoot. At the same time, under control conditions, their expression was downregulated in hvd14.d root relative to Sebastian (Daszkowska-Golec et al. 2023). It was previously shown that ABA may regulate lateral root formation (De Smet et al. 2003; Orman-Ligeza et al. 2018). However, the interactions between SLs and ABA have been described in various aspects of plant development under both control and stress conditions (Korek and Marzec 2023). Thus, we may conclude that the higher number of lateral roots observed in hvd14.d is related to the disorder in ABA signalling caused by the SL-insensitivity, similar to a weaker response of hvd14.d to drought stress (Daszkowska-Golec et al. 2023).
Another TF, HORVU.MOREX.r2.1HG0041130.1 (AT4G17980) mediates the auxin response and was upregulated in hvd14.d shoots. Auxin export, which is necessary for the outgrowth of axillary buds, is blocked by SLs to suppress shoot branch development (Shinohara et al. 2013). In hvd14.d, which develops a higher number of tillers, the increased auxin export induces auxin signalling, i.e. via expression of HORVU.MOREX.r2.1HG0041130.1. The last TF identified as SL-dependent under control and drought conditions was HORVU.MOREX.r2.3HG0209060.1 is an ortholog of WRKY6 in Arabidopsis (AT1G62300) and is described as being involved in response to low phosphate (Chen et al. 2009). Under phosphorus deficiency, WRKY6 binds the promoter of PHOSPHATE1 (PHO1) (Chen et al. 2009), increasing the production of lateral roots (Williamson et al. 2001). Given the observed root phenotype observed here, HORVU.MOREX.r2.3HG0209060.1 could play a broader role in SL-dependent repression of lateral root development in barley. Because SL treatment induced HORVU.MOREX.r2.1HG0041130 expression in WT plant, but not in the SL-insensitive mutant d14 (Fig. 4), we may assume that SLs control HORVU.MOREX.r2.1HG0041130 expression in a D14-dependent manner. Moreover, it was also previously shown that in response to drought, the expression of HORVU.MOREX.r2.1HG0041130 increases in Sebastian plants, but not in the d14 mutant (Daszkowska-Golec et al. 2023). On the other hand, exogenous GR24 induced the HORVU.MOREX.r2.1HG0041130 expression 1 (1 and 10 µM) or 3 h (1 µM) after treatment, but not after 30 min (Fig. 4). Thus, the temporal control of SLs on HORVU.MOREX.r2.1HG004113 can be postulated, which also depends on the SL concentration. Because, in older plants (3-week-old plants) grown in hydroponics, the increased expression of HORVU.MOREX.r2.1HG0041130 was observed in d14; the open question remains how plant developmental stage and growing conditions affect the expression of HORVU.MOREX.r2.1HG0041130
Assessment of the association between identified SL-dependent TFs revealed significantly more interactions than expected (PPI enrichment p-value:< 1.0e−16), indicating that the proteins are at least partially biologically connected. Moreover, 42% of all identified SL-dependent TFs were grouped in the single network of known and predicted interactions (Fig. 3). As expected, among all TFs, the proteins annotated as hormonal responsive were overrepresented. However, proteins involved in response to ethylene and salicylic acid were also identified, pointing out the interactions between SLs and those two phytohormones in shaping shoot and root architecture in barley. Finally, two out of three genes related to positive regulation of cutin biosynthesis were identified as SL-dependent TFs (Fig. 3). Cutin is a main component of the cuticle (Fich et al. 2016), with the biosynthesis pathway similar to other plant hydrophobic polymer suberin (Pollard et al. 2008) that accumulates in the apoplastic regions of non-cutinized boundary cell layers, such as root exodermis (Vishwanath et al. 2015). Previously, it was postulated that SLs modulate wax biosynthesis and deposition in plants (Li et al. 2020b; Marzec et al. 2020; Li et al. 2019).
Interestingly, genes controlling camalexin biosynthesis were found among SL-dependent TFs. Camalexin is one of the phytoalexins, which are the antimicrobial compounds produced by plants (Hammerschmidt 1999). SLs may play a dual role in interactions with bacteria and fungi to (1) promote the symbiosis with arbuscular mycorrhizal (AM) fungi (Kodama et al. 2022) or (2) increase the resistance against pathogen bacteria and fungi (Marzec 2016). Thus, it may be postulated that SLs control microbial interactions via camalexin synthesis. However, a new role of camalexin in controlling lateral root formation in Arabidopsis was recently described (Serrano-Ron et al. 2021). Up to now, a similar function of camalexin in monocots has not been reported. Still, it cannot be excluded that SL-insensitivity in barley disturbs camalexin biosynthesis, which affects lateral root development.

留言 (0)