
記住我




All procedures were performed in line with the University of Edinburgh and Home Office guidelines under the 1986 Animals (Scientific Procedures) Act, CPCSEA (Government of India) and approved by the Animal Ethics Committee of the Institute for Stem Cell Science and Regenerative Medicine (inStem).
Breeding and animal husbandry: ColoradoAll studies conformed to the requirements of the National Institutes of Health Guide for the Care and Use of Laboratory Rats and were approved by the Institutional Animal Care and Use subcommittee of the University of Colorado Anschutz Medical Campus (protocol 00411). All rodents were housed in micro-isolator cages with water and chow available ad libitum. Animals were bred in house on the Long Evans Hooded background and housed with littermates on a 12 h light/dark cycle with food and water ad libitum.
Experiments were performed on wild-type (WT; Cdkl5+/y) and Cdkl5−/y rats at post-natal day (P) 28 to 35 unless otherwise stated. All experiments and data analyses were performed blind to genotype.
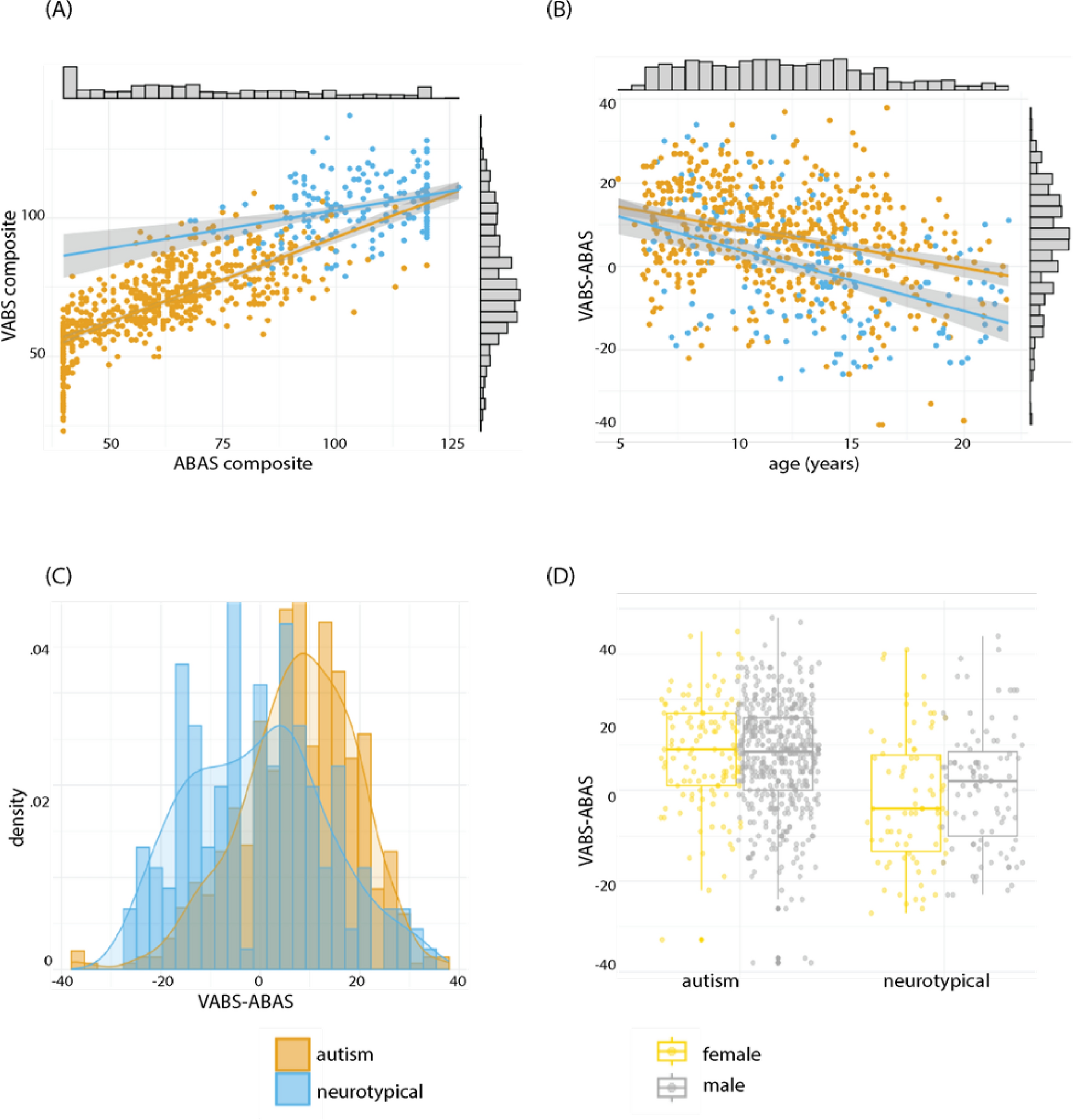
Cdkl5−/y rat generation and genotypingThe Cdlk5−/y rat model was created using CRISPR/Cas9 technology to introduce a 10 bp (bp) deletion in exon 8 of the Cdkl5 gene (Ensembl coordinates X:35,674,763–35,674,772, in the Rnor_6.0 genome assembly). An in-house PCR-based strategy was designed to genotype experimental rats produced from crossing Cdkl5−/y males with WT females. Forward and reverse primers were generated flanking the bp deletion site in exon 8 of the rat CDKL5 gene (F1 and R), a third forward primer which anneals to the 10 bp deletion site in the WT allele (F2) and a further forward primer which anneals over the deleted 10 base pairs in the knock out (KO) allele (F3) were also generated (Fig. 1).
Fig. 1
Validation of Cdkl5-/y rats. (A) Schematic of the Cdkl5 knockout strategy depicting the WT and null alleles. The null allele has a 10 bp (bp) deletion in exon 8 (region shown in blue in WT allele), leading to a frame shift and an in frame, premature STOP codon forming in exon 9 (*). (B) Genotyping results from male WT and Cdkl5−/y animals. Higher band in WT and Cdkl5−/y animals resulting from F1 and R primers product. Lower band in the WT samples resulting from F2 and R primer products is absent in the null samples due to the 10 bp deleted sequence. (C) Western blot showing the absence of CDKL5 in hippocampal and prefrontal cortex tissue preparations from WT and Cdkl5−/y rats. (D) Quantification of CDKL5 western blot protein expression in hippocampal and prefrontal cortex preparations. (E) Western blot showing the absence of CDKL5 in hippocampal synaptosome preparations from WT and Cdkl5−/y rats. (F) Quantification of CDKL5 western blot protein expression in hippocampal synaptosome preparations
F1: 5’ -GGGCTTGTAGCAAATCCATCC- 3’.
R: 5’ -AGCAAGCAGAGTTCTATTTTCCT- 3’.
F2: 5’ -ATACGTGGCTACTCGGTGGTAC- 3’.
F3: 5’ -CAGAATACGTGGCTACCGATC- 3’.
To distinguish between DNA derived from WT and Cdkl5−/y male littermates, primers F1, R and F2 were used in the same PCR reaction. Two bands were detected for WT male animals (356 and 135 bp) whereas only one band was detected for Cdkl5−/y male rats (346 bp) (Fig. 1). To distinguish between DNA derived from WT and heterozygous (Cdkl5+/−) female littermates, primers F1, R and F3 were used in the same reaction. One band was detected for WT female rats (356 bp) whereas two bands were detected for heterozygous female rats (356 and 129 bp).
Genomic DNA was extracted from fragments of tissue using the HotShot method. PCR was carried out as per the manufacturer’s guidelines for GoTaq G2 Polymerase (Promega, M784B) with an annealing temperature of 58 °C and a 1 min extension time. Following initial validation experiments all genotyping was carried out by Transnetyx Inc.
Acute slice preparation for electrophysiologyAcute brain slices were prepared from WT and Cdkl5−/y rats at postnatal day (P) 28 to 35 (unless otherwise noted) similarly to previously described [18]. Briefly, rats were anesthetised with isofluorane and subsequently decapitated. The brain was rapidly removed and placed in ice-cold carbogenated (95% O2/5% CO2) sucrose-modified artificial cerebrospinal fluid (in mM: 87 NaCl, 2.5 KCl, 25 NaHCO3, 1.25 NaH2PO4, 25 glucose, 75 sucrose, 7 MgCl2, 0.5 CaCl2). 400 μm horizontal hippocampal slices were cut on a Vibratome (VT1200s, Leica, Germany). Slices recovered submerged in sucrose-ACSF at 34 °C for 30 min and were then stored at room temperature until needed.
Alternatively, to assess NMDA receptor-mediated EPSCs throughout development and respective pharmacology, P7-22 WT and Cdkl5−/y rats were rapidly decapitated and the brain removed. Parasagittal slices (400 μm) were prepared on a Leica VT 1200 microtome in ice cold solution containing (in mM) 206 Sucrose, 2.8 KCl, 1.25 NaH2PO4, 26 NaHCO3, 10 Glucose, 10 MgSO4, 2 NaAscorbate, 0.4 CaCl2, and 2.5 N-acetyl L-cysteine. Scalpel cuts were made to remove CA3 while retaining the CA1 region of the hippocampus with the overlying cortex and dentate gyrus intact for electrophysiology. Slices were then recovered > 60 min at room temperature in a submersion chamber in standard artificial cerebral spinal fluid (ACSF), containing (in mM) 124 NaCl, 26 NaHCO3, 1.2 NaH2PO4, 10 D-glucose, 3 KCl, 2 NaAscorbate, 1 MgSO4, 2 CaCl2, and 2.5 N-acetyl L-cysteine) prior to all experiments. All solutions were oxygenated with 95% O2 − 5% CO2.
Field LTP recordingsSlices were transferred to a submerged recording chamber perfused with warm carbogenated recording ACSF (in mM: 125 NaCl, 2.5 KCl, 25 NaHCO3, 1.25 NaH2PO4, 25 glucose, 1 MgCl2, 2 CaCl2) at a flow rate of 3–4 mL/min. Extracellular field recording electrode was filled with recording ACSF and placed in the stratum radiatum (Str Rad) of the CA1 region. Single pulses of electric stimulation (200 µs, 0.5 Hz) were delivered through a bipolar electrode (Ni: Cr) placed in the Str Rad to stimulate the Schaffer collateral (SC) pathway. Stimulus intensity was adjusted to produce 50% of the maximum field excitatory post-synaptic potential (fEPSP) amplitude. LTP was induced by tetanic stimulation (two trains of 1 s 100 Hz stimulation, 20 s inter-train interval [19], following 20 min of stable baseline. fEPSP slopes were normalised to baseline values and LTP magnitude reported as the average fEPSP slope in the final 10 min (50–60 min post-induction) of the recording divided by the average fEPSP slope during the baseline period. Data acquisition and analysis were performed on WinLTP [20].
Whole-cell patch-clamp recordingsFor whole-cell recordings, slices were transferred to a submerged recording chamber perfused with warm carbogenated recording ACSF (in mM: 125 NaCl, 2.5 KCl, 25 NaHCO3, 1.25 NaH2PO4, 25 glucose, 1 MgCl2, 2 CaCl2), at a flow rate of 6–8 mL/min. All recordings were performed at 31 ± 1 ºC unless otherwise stated. Infrared differential inference contrast (IR-DIC) video microscopy, using a digital camera (Qimaging) mounted on an upright microscope (Olympus BX51WI) and a 40 × (0.8NA) water immersion objective was used for all experiments. Recordings were obtained with a Multiclamp 700B (Molecular Devices) amplifier, signals were Bessel filtered online at 5 kHz and digitized at 20 kHz (Digidata1440, Molecular Devices) coupled to the Clampex software (pCLAMP™ Software, Molecular Devices). Recording pipettes were pulled from borosilicate glass capillaries (1.7 mm outer/1 mm inner diameter, Harvard Apparatus, UK) on a horizontal electrode puller (P-97, Sutter Instruments, CA, USA), with resistance of 4–9 MΩ when filled with internal solution. For voltage-clamp recordings glass electrodes were filled with cesium based internal solution (in mM: 110 CsOH, 110 D-gluconic acid, 20 CsCl, 10 HEPES, 10 phospho-creatine, 4 MgATP, 4 NaCl, 0.3 Na2GTP, 0.2 EGTA, 5 QX314Cl) unless stated otherwise. A potassium gluconate based internal solution (in mM 120 K-gluconate, 20 KCl, 10 HEPES, 10 phospho-creatine, 4 MgATP, 4 NaCl, 0.3 Na2GTP, 2.7 biocytin, pH = 7.4, Osm = 290–310) was used for whole-cell current-clamp recordings. Cells were rejected if series resistance > 30 MΩ, or the series resistance changed by more than 20% over the course of the recording. No series resistance cancellation or junction potential corrections were performed.
Evoked EPSCsCA3 inputs to CA1 PCs were stimulated by placing a stimulating bipolar electrode (Ni: Cr or insulated tungsten) in the Str rad in hippocampal slices with the CA3 containing portion of the slice severed. A single 100 µs current pulse was delivered by an isolated constant current simulator (DS3, Digitimer.Ltd or WPI, Sarasota, FL). Evoked EPSCs were recorded in voltage-clamp using a cesium based intracellular solution.
NMDAR/AMPAR and paired-pulse ratioAMPA receptor-mediated EPSCs were recorded at -70 mV in the presence of 50 µM picrotoxin to block GABAA receptors. The same cell was then held at + 40 mV to record pharmacologically isolated NMDA receptor-mediated EPSCs in the presence of 50 µM picrotoxin and 10 µM CNQX. NMDAR/AMPAR ratios were calculated from peak amplitude of NMDA receptor and AMPA receptor-mediated EPSCs. We assessed paired-pulse ratio by evoking two EPSCs 50 ms apart, whilst holding the cell at -70 mV in the presence of 50 µM picrotoxin and calculating the ratio of the amplitude of the second EPSC relative to the first EPSC.
NMDA receptor subunit expression development and pharmacologyA cesium based internal solution containing (in mM) 135 CsMeSO4, 10 HEPES, 10 BAPTA, 5 Qx314, 0.3 NaGTP, 4 Na2ATP, 4 MgCl2, and 0.1 spermine, pH 7.25 with 1 M CsOH was used. Recordings were performed at room temperature and extracellular solution was exchanged at a flow rate of 3–4 mL/min. AMPA receptor-mediated EPSCs were recorded at -70 mV, and NMDA receptor-mediated EPSCs were recorded at + 40 mV. Peak current for NMDA receptor-mediated EPSCs was taken at 70 ms after the peak of the AMPA receptor-mediated EPSC. NMDA receptor sensitivity to block by GluN2B receptor antagonist Ro 25-6981 was determined by recording a 5 min baseline, followed by Ro 25-6981 (5 µM) perfusion onto the slice for 20 min.
AMPA-R I-V relationshipTo assess the presence of CP-AMPA receptors, AMPA receptor-mediated EPSCs were recorded in the presence of 50 µM picrotoxin to block GABAA receptors and 50 µM AP-5 to block NMDA receptors, over a range of voltages from − 80 mV to + 40 mV. Rectification index was calculated by determining the difference of peak EPSC amplitude at -80mV and at + 40 mV to EPSC amplitude at 0 mV and calculating the ratio between those two values in order to account for variations in AMPAR reversal potential across recordings [21]; in this instance, a rectification index of 2 would indicate the absence of rectification. The same intracellular cesium based intracellular solution was used with added 0.1 mM spermine to maintain rectification of GluA2-lacking AMPA receptors.
Minimal stimulationMinimal stimulation protocol was used to assess the presence of silent synapses. Once a reliable EPSC was identified at -70mV, stimulus amplitude was reduced until the synaptic response would fail in some of the trials, allowing for the stimulation of a single or a small number of synapses. Following recording of 50 trials at a holding potential of -70 mV, corresponding to AMPA receptor-mediated EPSCs, the cell was depolarised to + 40 mV, to reveal mixed AMPA and NMDA receptor-mediated EPSCs and an additional 50 trials were recorded. To determine response probability the traces for each holding potential were visually inspected and the number of traces with a visible EPSC was divided by the total number of traces for each cell. The ratio of response probability at the two holding potentials was used as an estimate for the relative abundance of silent synapses [22, 23].
Miniature EPSC recordingsMiniature EPSCs (mEPSCs) were recorded in voltage clamp while holding the cell at -70 mV, using a cesium gluconate based internal solution. Recordings were performed in recording ACSF in the presence of 50 µM picrotoxin and 300 nM TTX to block voltage gated sodium channels and consequently action potential firing. Analysis of mEPSC frequencies and amplitudes over two minutes of recording was performed using a template matching algorithm [24] in Stimfit [25]. A similar number of mEPSC events were analysed for each condition – around 500 events, on average, per cell.
Intrinsic physiologyPassive and active membrane properties were assessed to examine intrinsic excitability as previously described [18]. Passive membrane properties, including membrane time constant and input resistance, were measured from the voltage response to a 500 ms hyperpolarizing 10 pA step. Rheobase current and action potential (AP) firing frequency were determined from a series of depolarising current steps (0 to + 400 pA, 500 ms) while holding the cell at -70mV with a bias current. AP properties were determined from the first AP elicited. All analysis of electrophysiological data was performed using the open source software package Stimfit [25], blinded to genotype.
Synaptosome preparationWT and Cdkl5−/y rats were killed by exposure to increasing CO2 and decapitated. The hippocampus from each hemisphere was dissected in ice-cold 1x sucrose-EDTA buffer (0.32 M sucrose, 1 mM EDTA, 5 mM Tris, pH 7.4). The tissue was snap-frozen and stored at -80 °C until used for synaptosome preparation. On the day of preparation, the tissue was quickly thawed at 37 °C and homogenized in ice-cold 1x sucrose/EDTA buffer using 5–6 up-and-down strokes of a pre-chilled Teflon glass with motorized homogenizer [26]. Homogenates were centrifuged at 950 x g for 10 min at 4 °C. The discontinuous (3% uppermost, 10% middle and 23% bottom) Percoll-density gradient was prepared prior to homogenization. The supernatant (S1) was added gently on 3% Percoll-sucrose (Percoll, P1644, Sigma Aldrich, UK) and centrifuged at 47,808 x g for 8 min at 4 °C. The fraction between 23% and 10% was collected and re-suspended in HEPES-Buffered-Krebs (HBK; in mM: 118.5 NaCl, 4.7 KCl, 1.18 MgSO4, 10 Glucose, 1 Na2HPO4, 20 HEPES, pH 7.4 balanced with Trizma) followed by centrifugation at 20,199 x g for 15 min at 4 °C. The pellet containing pure synaptosomes was dissolved in RIPA buffer (phosphatase inhibitor and protease inhibitor added). Protein quantification was performed with MicroBCA Assay kit (Pierce BCA protein estimation kit, 23,225, ThermoFisher Scientific).
Western blotsApproximately 10 µg of synaptosome protein was separated on a precast gradient gel (NuPAGE 4–12% Bis-Tris Protein Gels, NP0336BOX, Thermo Fisher) and transferred to nitrocellulose membrane (AmershamTM Protran® Western Blotting Membrane, Nitrocellulose, GE10600002, Sigma Aldrich) using Bio-Rad transfer apparatus. Total proteins were stained using reversible protein stain kit (Memcode 24,580, Thermo Fisher Scientific) according to the manufacturer’s instructions. After removing the stain, membranes were blocked with 1:1 TBS1X: Odyssey Blocking Buffer (P/N-927-50003, LI-COR Biotech.) for an hour at room temperature, followed by overnight incubation with primary antibodies (CDKL5- 1:1000, #HPA002847, Atlas Antibodies-Sigma Aldrich; NMDAR1-1:1000, #700,685, Thermo Fisher; NMDAR2A-1: 1000, #ab169873, Abcam; NMDAR2B- 1:1000, #610,417, BD Biosciences; PSD95- 1:2000, #76,115, Abcam; GluR1- 1:1000, #MAB2263, Millipore; GluR2- 1:1000, #MABN1189, Millipore; RIM1/2 − 1:2000, #140,203, SYSY; Munc18-1- 1:2000, #116 011, SYSY; SNAP25- 1:1000, #111 011, SYSY; Synapsin1- 1:1000, #ab64581, Abcam; Synaptophysin- 1:10,000, #ab32127, Abcam; VAMP2- 1: 10,000, #ab3347, Abcam) at 4 °C. Membranes were washed with TBST1 × (0.1% Tween 20), and incubated for an hour at room temperature with secondary antibodies (IRDye 800CW Goat anti Rabbit IgG- 1:10,000, #P/N 925-32211; IRDye 680LT Goat anti Mouse IgG- 1:10,000, #P/N 925-68020, LI-COR Biotechnology). Membranes were washed with TBST1X, dried and digitally scanned using Fc Odyssey Infrared Imaging System, LI-COR, UK Ltd. Odyssey software, Licor Image Studio Lite (LCOR Biosciences) was used to quantify individual bands. Data was normalised to respective total protein and then normalised to WT.
Sample preparation and mass spectrometryPurified synaptosomes were lysed in 8 M urea with protease and phosphatase inhibitor cocktail, sonicated for 30 s max amplitude and power on the UP200St sonicator (Hielscher – Ultrasound Technology), and protein quantified by BCA using a Nanodrop 1000. Samples (100 µg) were reduced with 5 mM TCEP for 30 min at RT, and 30 mM Iodoacetamide was added for 30 min at RT in the dark. Samples were diluted to 1 M Urea with 50 mM TEAB pH 8.5, and digested with 2 µg Trypsin overnight at 37 °C, before being desalted using a macro-spin column (Harvard Apparatus) according to manufacturer protocol.
For mass spectrometry analysis, peptides were resuspended in HPLC-grade water containing 2% MeCN and 1% TFA to make a final concentration of 1 µg/µl (based on protein concentration prior to digest). Then, 5 µl of samples were injected for analysis. Peptides were separated using 50 cm Acclaim PepMap 100 analytical column (75 μm ID, 3 μm C18) in conjunction with a PepMap trapping column (100 μm × 2 cm, 5 μm C18) (Thermo Scientific) analysed with Orbitrap Fusion Tribrid mass spectrometer (Thermo-Fisher Scientific). Peptides were loaded on to the column in solvent A (3% MeCN, 0.1% FA in HPLC water). A 218 min gradient was performed with 3% solvent A to 35% solvent B (80% MeCN, 0.08% FA), before increasing solvent B to 90% for 5 min, and re-equilibrating with 3% solvent A for 10 min. Settings for data dependent acquisition were MS1 with 120,000 resolution, scan range 400–1,600, charge state 2–7, AGC target of 400,000 and dynamic exclusion of 50 ms with repeat count 1. Peptide ions were fragmented using HCD (30% collision energy) with a resolution of 30,000, and AGC target of 10,000 with a maximum injection of 40 ms. The whole duty cycle was 3 s, during which the instrument performed “top speed” analysis.
Proteome quantificationLabel-free quantification was performed using MaxQuant (v.1.5.7.4) with the following modifications: fixed modification: carbamidomethyl (C); variable modifications oxidation (M), acetylation (protein N-terminus); label-free quantitation with minimum ratio count 2; maximum five modifications per peptide, and two missed cleavages. Searches were conducted using a Rattus norvegicus Uniprot-Trembl database (downloaded 27th July 2017 with 36,078 entries) and a list of common contaminants. Identifications were filtered at a 1% false-discovery rate (FDR). Quantification used only razor and unique peptides with a minimum ratio count of 2. “Re-quantify” was enabled. “Match between runs” was used with alignment time window 20 min and match time window 0.7 min. LFQ intensities were used for data analyses. The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE partner repository (https://www.ebi.ac.uk/pride/) with the dataset identifier PXD043246 and can be viewed by reviewers with the following details: Username: reviewer_pxd043246@ebi.ac.uk; Password: L0K4DUac.
HistologySlices used for electrophysiology experiments were fixed in 4% paraformaldehyde (PFA) overnight and stored in PBS (phosphate buffered saline) at 4 ºC until used for histology. Slices were washed in PBS and incubated in PBS with 0.3% triton-X and Alexa488 or Alexa568-conjugated streptavidin (1:500 dilution, Molecular probes, Invitrogen, USA) over night. Slices were then washed in PB and mounted on glass slides using Vectashield Hardset mounting medium (H-1400, Vector Labs).
Image acquisition and analysisTo reconstruct cells and examine their morphology, multiple Z-stacks were taken in order to capture the entire biocytin-filled cell on an inverted confocal microscope (Axiovert LSM510, Zeiss) under a 20x Plan Neofluar (NA 0.5) objective (Zeiss). The Z-stacks obtained for a given cell were stitched using the 3D stitching plug in FIJI (ImageJ), and the cell was reconstructed using the Simple Neurite Tracer plug-in [27]. Sholl analysis was then performed on the skeletonised paths to examine dendritic complexity. To examine spine distribution in biocytin-filled cells, Z-stacks were taken from basal and apical (oblique and tuft) dendrites (2–3 dendrite sections per dendrite type per cell). Spines were imaged under a 63x Plan Apochromat (NA 1.4) oil immersion objective on an inverted confocal microscope (Axiovert LSM510, Zeiss), with a 2.8x zoom, 2x average line scan, 1024 × 1024 resolution, 0.14 μm Z step. Huygens Essential software (Scientific volume imaging, Netherlands) was used for deconvolution. The deconvolved images were used for analysis on FIJI (ImageJ). Z-projections of the deconvolved Z-stacks were used to manually count spines using the cell counter tool. For each dendrite section, the number of spines was normalised to the length of the section of dendrite analysed.
Statistical analysisAll experiments and data analysis were performed blind to genotype. All statistics were performed using linear mixed effects modelling (or the generalised form), when such analysis was not possible data were compared using animal averages to avoid the risk of type-1 statistical errors. Mixed modelling was implemented using the R package lme4 in RStudio [28]. Genotype was set as fixed effect and animal, slice (and cell where relevant) as random effects, allowing for direct measurement of genotype effect while accounting for the variability resulting from random effects. For alternative statistical tests Graphpad prism 7 was used to perform comparisons across groups with two-tailed unpaired T-tests, repeated measures two-way ANOVA, or non-parametric tests as appropriate. In many figures data are shown from both cell and animal, the latter delineated through the use of the prime (‘) symbol. Where only one dataset is shown, these are generated from animal averages unless otherwise stated in the figure legend. Details on sample size and statistical test used are presented in the results text and figure legends.
留言 (0)