
記住我
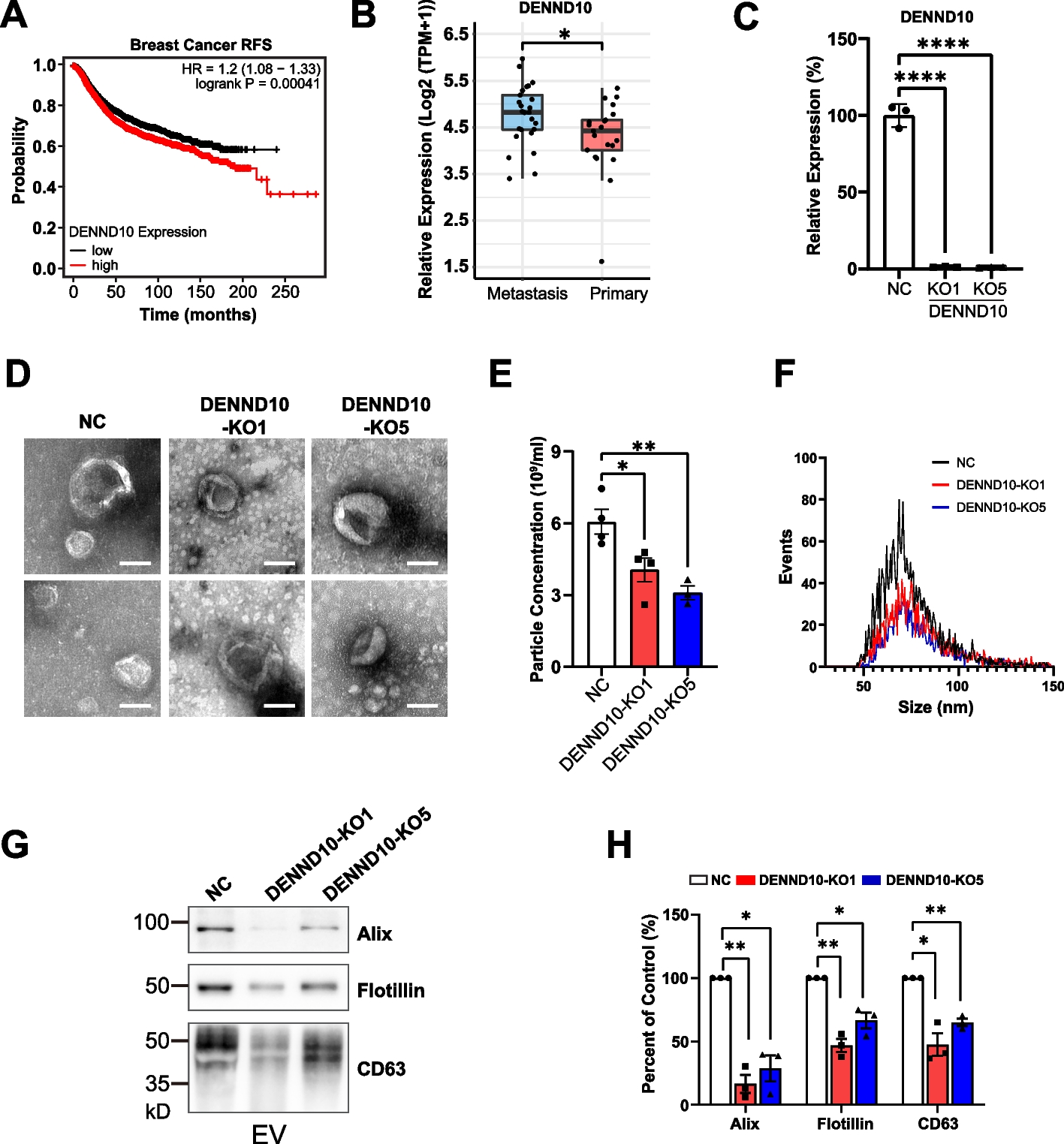


To shed light onto the evolution of fructophily and alcoholic fermentation, we expanded the number of species in the W/S clade for which whole-genome data is available (Additional file 1: Table S1). Raw data were assembled and annotated using the same pipelines for all species, irrespective of the origin of the data. The resulting phylogenomic tree encompasses 72 species (63 in the W/S clade and nine outgroups) and is based on 273 single-copy orthogroups (SCO) present in all species (Fig. 1). Its topology confirmed previous observations indicating that several Wickerhamiella species cluster robustly with species currently assigned to the genus Starmerella (subclade 4) [7]. These include the type species of the genus, Wickerhamiella domercqiae. For this reason and others that will become apparent, we shall refer to these (five) Wickerhamiella species together with all Starmerella species as the Starmerella subclade (subclade 4) and to the remaining species as the Wickerhamiella subclade (comprising subclades 1, 2, and 3). This topology is not a consequence of the fact that many, possibly all, W/S-clade species have numerous genes acquired horizontally from bacteria and from filamentous fungi (Pezizomycotina), since it is very unlikely that any such genes were represented in the set of SCO used to infer the phylogeny; only SCO present in all species were considered, and HGT-derived genes tend to be present only in a fraction of the taxa [8,9,10,11]. The expanded number of species included in this tree produced a more complex topology within both the Wickerhamiella and Starmerella subclades, thereby improving the resolution of the framework onto which previously identified evolutionary events can be mapped.
Fig. 1
Phylogenomic tree of the W/S clade and distribution of fructophily and fermentation-related genes and phenotypes. A Maximum likelihood phylogenomic tree comprising 63 W/S-clade species and nine outgroups, inferred from the concatenated alignment of 264 SCO and rooted with Tortispora caseinolytica based on the phylogenetic analysis of Shen et al. 2018 [2]. The presence/absence of genes related to fructophily and alcoholic fermentation, as well as phenotypes determined for a subset of species, are shown to the right of the tree. Subclades (labeled 1–4) were defined according to the ADH1 genotype. B Relevant metabolic pathways for production of fermentation byproducts in the W/S clade (mannitol, glycerol, and ethanol). Names of the enzymes carrying out the represented reactions are shown next to the respective arrow: Hxk—hexokinase, Glk—glucokinase, Pgi—phosphoglucoisomerase, Mtdh—mannitol dehydrogenase, Pdc—pyruvate decarboxylase, Aro10—phenylpyruvate decarboxylase, Adh—alcohol dehydrogenase, Gpd—glycerol 3-phosphate dehydrogenase, Gpp—glycerol 3-phosphate phosphatase. C Sugar consumption profiles for eight W/S-clade species cultivated in medium containing glucose and fructose (20FG) are shown, highlighting the diversity in sugar consumption rates and preferences across the W/S clade. For each species, the presence/absence profile of genes and traits depicted in A is shown above the plots. Sugar consumption profiles and fermentation byproduct formation profiles all species as well as respective replicates are shown in Additional file 5: Fig. S4
Multiple independent HGT events involving alcoholic fermentation-related genesThe distribution and phylogenetic relationships of alcoholic fermentation-related genes across the W/S clade show three clearly separated groups in the ADH1 phylogeny (Additional file 2: Fig. S1) and four in the ADH6 phylogeny (Additional file 3: Fig. S2), suggesting that seven independent HGT events were involved in the acquisition of ADH1-like (total of three events) and ADH6-like genes (total of four events).
Starmerella bombicola was previously shown to harbor one ADH1-like and two ADH6-like genes [8]. Considering the entire W/S clade, many species have similarly one ADH1 gene, with some harboring two and W. versatilis containing three ADH1 genes (Fig. 1). ADH6 genes are almost always present in multiple copies ranging in number from two to ten. Phylogenetic analyses of the ADH1 and ADH6 genes found in W/S-clade species show that whenever more than one gene is present, duplications occurred after speciation, as multiple ADH1 or ADH6 genes present in any given species always clustered together (Additional file 2: Fig. S1, Additional file 3: Fig. S2). The distinct bacterial origin of the ADH genes, suggesting multiple independent acquisitions, is remarkably consistent with the topology of the phylogenomic tree (Fig. 1). For ADH1, three different bacterial origins were identified in three different subclades. All species in the Starmerella subclade (subclade 4) possess ADH1 genes from Acetobacteraceae (named Adh1a). In subclade 1, all species possess a ADH1 gene (named Adh1b) possibly derived from the Enterobacterales, while in subclade 3, all but two species have an ADH1 gene possibly originating in Acinetobacter (named Adh1c). The two exceptions are W. pagnoccae and W. jalapaonensis, both lacking ADH1 genes, probably as consequence of a secondary loss in their common ancestor. Finally, subclade 2 contains five species all seemingly lacking ADH1 genes. Plotting the distribution of the genes onto the phylogenomic tree, we postulate that the three types of ADH1 genes found in Wickerhamiella and Starmerella species were each independently acquired by the ancestor of the subclade in which they are presently found while subclade 2 seems never to have acquired a bacterial ADH1 gene.
A more complicated pattern emerges for ADH6 genes, where four different bacterial genes seem to have been acquired by three different subclades, as inferred from the distribution of ADH1. Specifically, subclade 4 harbors Adh6a enzymes possibly originating in the Sphingomonadales whereas most species of subclade 3 contain multiple ADH6c genes possibly originating in the Alteromonadales. However, subclade 1 seems to lack ADH6 genes, while two species in the Adh1null subclade 2 contain ADH6 genes that seem to be of different bacterial origins (Adh6d and Adh6e).
W/S-clade species harboring different bacterial alcohol dehydrogenases exhibit distinct fermentative phenotypesWe showed previously that in St. bombicola Adh1a was mainly responsible for alcoholic fermentation but also ensured ethanol assimilation [8]. By contrast, Adh6 enzymes in St. bombicola are unconnected with alcoholic fermentation, as in Saccharomyces cerevisiae, although they might make a minor contribution to this process in the absence of Adh1 [8]. This agrees with the fact that in most if not all species studied so far, the interconversion of acetaldehyde and ethanol is ensured by a NADH-dependent Adh1 type of enzyme [22,23,24], while Adh6-type enzymes are deemed to be broad range, NADPH-dependent enzymes that accept a variety of aldehydes as substrates (Fig. 1B) [25, 26]. In conformity with this, all species tested in the Starmerella subclade (subclade 4) conducted alcoholic fermentation except for W. bombiphila (Fig. 1). However, almost without exception, species encoding Adh1b or Adh1c types of enzymes seemed to be poor ethanol producers (< 5 g/L) under the conditions tested. Unlike Adh1a, these enzymes may be involved mainly in ethanol assimilation (Additional file 1: Table S1). If that is the case, Adh1b and Adh1c would functionally resemble S. cerevisiae Adh2, an enzyme specialized in ethanol assimilation and that does not participate in the inverse reaction [27]. Nevertheless, we found two exceptions in subclade 1, the closely related species W. spandovensis and W. sergipiensis, which produce ethanol in substantial amounts, presumably using Adh1b. The most intriguing case is, however, W. slavikovae which is capable of robust fermentation while presumably lacking both Adh1 and Aro10, the enzyme deemed to have replaced Pdc in alcoholic fermentation in the W/S clade [8]. Absence of fermentation genes in W. slavikovae was confirmed by inspection of an additional high-quality assembly produced using long reads (Nanopore technology) (GCA_954870865). Even if we assume that one or more of the products of the five ADH6 genes found in this species may be responsible for ethanol formation, the enzyme responsible for the conversion of pyruvate to acetaldehyde remains elusive. An alternative pathway for ethanol production might involve an AdhE type of multifunctional enzyme, commonly found in bacteria [28]. However, a gene encoding this kind of enzyme, which might fulfill the roles of both Adh1 and Pdc, was also absent from the W. slavikovae genome. One additional species, W. kazuoi, also lacks Pdc, Aro10, and Adh1. These observations indicate that the loss of Adh1 and Aro10 might be connected, suggesting that the two enzymes are functionally linked.
In summary, we identified at least seven independent events of acquisition of ADH genes, involving four W/S subclades. ADH1 or ADH6 genes were each acquired by the ancestor of a subclade, except for Adh6d and Adh6e, which are found each in a single species. The ancestors of subclades 1 and 4 acquired Adh1 and Adh6 at the same point in evolution but from different bacterial donors (Additional file 2: Fig. S1, Additional file 3: Fig. S2). In the Starmerella subclade (subclade 4), this formed the basis for a fermentative capacity that persisted in almost all species, while in the two Wickerhamiella subclades possessing Adh1 (subclades 1 and 3), ethanol assimilation might be the most likely role of the enzyme. As for Adh6, an involvement in alcoholic fermentation seems so far negligible, with the possible exception of Adh6d in W. slavikovae; a likely hypothesis may be that Adh6 play a role in detoxification, carrying out the conversion of aldehydes of environmental origin to the less toxic corresponding alcohols [29,30,31]. Whatever the role, multiple copies of bacterial ADH6 genes are widespread across the entire W/S clade, while most species carry only one ADH1 gene. As to the evolution of Pdc/Aro10, it is notable that only W. versatilis acquired bacterial Pdc genes (Additional file 4: Fig. S3), as previously reported [8]; no other bacterial Pdc genes were found in other W/S-clade species (Additional file 4: Fig. S3). It cannot be excluded that the inability of species in the Wickerhamiella subclades to conduct alcoholic fermentation is due to the absence of Pdc activity, which is not required for ethanol assimilation. In other words, it cannot be excluded that co-optation of Aro10 for alcoholic fermentation occurred only in a subset of W/S-clade species. At variance with this possibility, the very few Wickerhamiella species lacking Aro10 are among the few that also lack Adh1, including one likely to have experienced Adh1c secondary loss, suggesting a functional link between the two enzymes. Finally, we found a strong ethanol producer, W. slavikovae in the Adh1-null subclade (subclade 2), that also lacks Pdc/Aro10 (Fig. 1). Together, these observations suggest a complex evolutionary pattern involving Aro10 and its role in fermentation in the W/S clade. These observations also imply the existence of yet unidentified enzymes promoting ethanol production in substantial amounts in species lacking Adh1 and Pdc/Aro10, like W. slavikovae.
The phenotypic landscape across the W/S clade reveals a wide range of fermentation ratesThe general trend observed in Fig. 1 is that Starmerella species conduct alcoholic fermentation while species in the Wickerhamiella subclades generally do not. However, the amount of ethanol (and other fermentation byproducts—mannitol and glycerol) (Fig. 1B) produced by fermentative species and their production rates vary considerably (Additional file 5: Fig. S4, Additional file 6: Table S2). The correlation between global (considering all products together) maximum fermentation byproduct production rates determined for 42 species studied and the extremely variable sugar consumption rates observed is shown in Fig. 2A. Not surprisingly, the rate of global fermentation byproduct formation robustly correlates with the sugar consumption rate measured using the same time point (r = 0.96, p-value < 2.2e−16, Pearson correlation test) (Fig. 2A), showing no major byproduct of fermentation was overlooked. Hence, as usually observed [32], sugars are consumed more rapidly during fermentative metabolism to compensate for the lower energetic yield of fermentation. Interestingly, we found that the phenotypic landscape of fermentation and sugar consumption rates covers a wide spectrum of values (Fig. 2A), indicating that the proportion of fermentative metabolism vs respiratory metabolism is extremely diverse across the clade under the conditions tested. Globally, the correlation between fermentation rates and sugar consumption is mainly driven by ethanol production (r = 0.95, p-value < 2.2e−16, Pearson correlation test), since, as shown on the inset of Fig. 2A, the correlation between sugar consumption rates and fermentation rates is barely affected when glycerol and mannitol are excluded (Fig. 2A, inset). We corrected these correlations for the effect of phylogenetic relatedness using PIC (phylogenetically independent contrasts) (Additional file 7: Fig. S5) and showed that the robust correlation is maintained in both cases (all fermentation byproducts: adjusted R-squared = 0.837, p-value < 2.2e−16; only ethanol: adjusted R-squared = 0.779, p-value < 6.62e−15). In line with our previous observations, the highest sugar consumption and fermentation rates are found in species belonging to the Starmerella subclade (subclade 4) (Fig. 2A, Additional file 8: Fig. S6). This is also noticeable when the absolute amounts of fermentation byproducts (in the form of individual byproduct yields) are compared between the two groups (subclade 4 and the remaining three Wickerhamiella subclades) (Fig. 2B). In Wickerhamiella, mannitol and glycerol account for a much larger fraction of the fermentation byproducts than in Starmerella (Fig. 2B), which is consistent with the observation that the global fermentation byproduct formation rates drop to zero in most of these species when only ethanol is considered (Fig. 2A, inset). Hence, Starmerella species resort almost without exception to alcoholic fermentation for energy conservation often with concomitant formation of mannitol and/or glycerol, while the majority of Wickerhamiella species use respiration and exhibit a much wider diversity concerning byproduct formation, ranging from considerable amounts of mannitol and/or glycerol to nearly none (Fig. 2B).
Fig. 2
Comparison of respiration vs fermentation preferences across the W/S clade. A Correlation between maximum global fermentation byproduct production rates (ethanol, glycerol, and mannitol) and sugar consumption rates across the W/S clade, highlighting Wickerhamiella (subclades 1–3) and Starmerella (subclade 4) species in different colors. The inset (top, right) shows the correlation between sugar consumption rate and ethanol production rate determined at the same time points for the same species, denoting that the correlation is mainly driven by ethanol production. Each datapoint represents a single measurement for each species, but results for a replicate assay can be assessed in Additional file 6: Table S2 and Additional file 8: Fig S6A. B Fermentation byproduct (ethanol, glycerol, and mannitol) individual yields determined for each byproduct at the time point at which the maximum global yield was measured for each species (replicate assay can be assessed in Additional file 6: Table S2). The more pronounced bias towards fermentative metabolism in the Starmerella clade (subclade 4) when compared to Wickerhamiella (subclades 1–3) is noticeable. The manifest difference between Starmerella (subclade 4) and Wickerhamiella (subclades 1–3) concerning the main fermentation byproduct formed is highlighted in the inset (top, right) where the proportions of the three byproducts formed by each species are shown
Presence of HGT-derived FFZ1 is pervasive across fructophilic speciesPossibly the most unusual trait that is widely spread in the W/S clade is the preference for fructose over glucose when both sugars are present in large amounts. The preference is often far from subtle as in some of the species studied so far, glucose consumption only picked up after a considerable drop in fructose concentration (Fig. 1C). The cornerstone of this marked fructophilic behavior is a remarkable high-capacity fructose transporter (Ffz1) that evolved from a Drug/H+ antiporter relatively recently, therefore lacking the usual signatures of sugar porters [12]. All the evidence indicates that FFZ1 has been horizontally acquired from the Pezizomycotina early in the evolution of the W/S clade [12]. Initial research found an almost perfect correlation between the presence of FFZ1 in the genome and fructophily [12, 33]. To find out whether this strong correlation persisted after the substantial increase in the size of the genome sequence dataset here reported, we scored the presence of the FFZ1 gene in all species represented in the phylogenomic tree in Fig. 1A, reconstructed the Ffz1 phylogeny (Additional file 9: Fig. S7), and determined sugar preference for a large fraction of the species (Additional file 5: Fig. S4). In the past, we put forward a hypothesis to explain the patchy distribution of FFZ1 across the Saccharomycotina which involved two HGT events (from the Pezizomycotina to the W/S clade and from the W/S clade to Zygosaccharomyces) [12]. However, the support for the HGT event from the W/S clade to Zygosaccharomyces was limited by the low number of genomes available. The updated Ffz1 phylogeny inferred with additional W/S species sequences (Additional file 9: Fig. S7) further supports the two HGT events, as yeast Ffz1 sequences cluster within filamentous fungi, whereas all Zygosaccharomyces sequences form a deeply embedded clade within the Starmerella subclade. We found again an almost perfect correlation between the presence of the gene and fructophily in the Starmerella subclade (subclade 4).
FFZ1-independent fructophily is patent in a restricted group of Wickerhamiella speciesIn the three Wickerhamiella subclades, FFZ1 was found only in a small number of species, most of which were fructophilic. Surprisingly, however, in subclade 3 (Fig. 1A), fructophily was detected despite the absence of FFZ1 in seven species. When compared with the sugar consumption profiles observed in FFZ1-harboring species (indicated as FFZ1 + in Fig. 1C, e.g., W. cachassae), the species lacking the gene are generally slower consumers of fructose (indicated as FFZ1—in Fig. 1C). Also, the sugar consumption profiles in Fig. 1C and Additional file 5: Fig. S4 show that in fructophilic species of the Wickerhamiella subclades, both with and without FFZ1, glucose tends to be left nearly untouched, which might denote a defect in glucose metabolism. If this were the case, fructophily might be the result of this defect, as opposed to a competition between the two sugars for uptake or phosphorylation. To assess that, we cultivated four fructophilic species lacking FFZ1 (FFZ1 −) species as well as four fructophilic species harboring FFZ1 (FFZ1 +) on glucose and fructose separately (using, respectively, 20G and 20F medium) and measured sugar consumption rates under the different conditions (Fig. 3). No defects on glucose utilization were observed in any of the species; even when very small amounts of glucose were consumed, high cell densities were attained reflecting a higher biomass yield on glucose than on fructose (Additional file 6: Table S2). When compared with the individual glucose and fructose consumption rates in medium containing both sugars, rates were generally higher in the single sugar condition (p-value = 0.0148, Wilcoxon test) (Fig. 3), showing that there is competition between the utilization of the two sugars as might be expected as a consequence of their very similar metabolisms. Notably, the presence of FFZ1 appears to slightly mitigate the negative effect of glucose on fructose consumption rates in mixed sugar cultures, while it slightly increases the negative effect of fructose on glucose consumption rates under the same conditions. This competition between the two sugars suggests that they use the same or a largely overlapping set of transporters, with Ffz1 tipping the balance towards fructose uptake when it is present. In line with this, no major differences regarding the number or type of sugar transporters were observed between fructophilic and glucophilic species lacking FFZ1 (Additional file 10: Table S3), suggesting that the milder fructophily observed in the absence of Ffz1 might be achieved in the absence of an alternative specific fructose transporter. In summary, the fructophilic character of the FFZ1-lacking species in subclade 3 suggests either that the available transporters have more affinity for fructose than for glucose or that fructophily in these cases is determined by the kinetic properties of the hexokinases.
Fig. 3
Comparison of glucose and fructose consumption rates in fructophilic species with and without FFZ1. Plots denote glucose (left panel) and fructose (right panel) consumption rates in culture media containing either glucose (20G medium) or fructose (20F medium) as sole carbon source or both sugars simultaneously (20FG medium), as indicated below the plots. Within each panel, results for FFZ1 + and FFZ1 − species are shown separately, as indicated, denoting the impact of the presence of the high-capacity fructose transporter on sugar consumption rates under the various conditions. Each datapoint represents a single experiment (replicate experiment can be found in Additional file 6: Table S2 and Additional file 8: Fig. S6B)
Differences in hexokinase activity do not explain FFZ1-independent fructophilyAs glucose and fructose metabolisms are so similar, the only two metabolic reactions potentially determining a preference for one or the other of the two hexoses are the transport and phosphorylation steps (Fig. 4A). Early in the study of strongly fructophilic Zygosaccharomyces species [34], it was established that the transport step was the culprit, later leading to the identification of FFZ1 [35]. The finding of fructophily independent of FFZ1 in some Wickerhamiella species led us to re-assess whether sugar phosphorylation kinetics might determine a preference for fructose in these species. To that end, glucose and fructose phosphorylation kinetics were determined in cell-free extracts of several Wickerhamiella species exhibiting different trait combinations (Fig. 4B; Additional file 11: Table S4), with a focus on sugar preference and the presence/absence of FFZ1. The results, shown in Fig. 4B, indicate that a bias of hexokinase activity towards fructose is unlikely to determine fructophily in any of the species examined, as the Km for fructose is always considerably higher than the Km for glucose. Nevertheless, the various species exhibited markedly different fructose/glucose Km and Vmax ratios (Fig. 4C). The two glucophilic species had the lowest Vmax ratios and highest Km ratios, which is in line with their preference for glucose, while the fructophilic species tended to have lower Km ratios and higher Vmax ratios. This suggests that although sugar phosphorylation does not seem to determine fructophily, hexokinase activity seems to have evolved towards improving fructose metabolism in fructophilic species, both with and without FFZ1. Hence, taken together, our results suggest that the transport step is mainly responsible for fructophily in FFZ1-lacking species, probably due to the higher affinity for fructose of one or more of the hexose transporters, but that there is also some adjustment of the kinetics of hexokinase to fructophily.
Fig. 4
Kinetic properties of hexokinase activity in Wickerhamiella species with different trait combinations. A First steps of glucose and fructose metabolism after internalization, catalyzed by hexokinase (Hxk), glucokinase (Glk), and phosphoglucoisomerase (Pgi). B Km of hexokinase activity using either fructose or glucose as substrates, measured in cell-free extracts of the species listed in Additional file 11: Table S4; dots are colored according to species traits, as indicated. Km of hexokinase activity for fructose is significantly higher than for glucose (p-value = 0.0018, t-test). C Variation in substrate bias of hexokinase activity as denoted by Km (fructose)/Km (glucose) ratios for the various species studied, plotted against the respective Vmax(fructose)/Vmax (glucose) ratios, showing that hexokinase activity tends to be less unfavorable towards fructose utilization in fructophilic species, color coded as in B
Higher fructose consumption rates in Wickerhamiella are associated with a metabolic shift towards fermentationHigher sugar consumption rates are usually associated with fermentative metabolism. For that reason, we decided to investigate whether the often marked difference between fructose and glucose consumption rates in fructophilic Wickerhamiella species (Fig. 3 and Additional file 6: Table S2) reflected a sugar-dependent preference for respiration or fermentation. Notably, even when glucose is left nearly untouched and fructose is efficiently consumed (for instance in W. nectarea or W. jalapaonensis, Additional file 6: Table S2), the maximum cell density reached on glucose is similar to or higher than that reached on fructose-based medium, suggesting that glucose-grown cells are shifting their metabolism towards the more energy effective respiration at the detriment of fermentation. To ascertain this, we determined the global fermentation byproduct yields (i.e., ethanol, mannitol, and glycerol considered together) for eight fructophilic Wickerhamiella species grown separately on glucose and on fructose (Additional file 6: Table S2). In all species, the fermentation byproduct yields were significantly higher on fructose compared to glucose (p-value = 0.002742, Wilcoxon Test) (Fig. 5), ranging from 2 × in W. occidentalis to 25 × in W. azyma (Additional file 6: Table S2). This increase in byproduct formation when fructose is the carbon-source was not restricted to mannitol formation, which might be expected because mannitol can be formed directly from fructose (Additional file 6: Table S2). Instead, in some species, like W. cachassae, W. nectarea, W. azyma, and W. jalapaonensis, an increase in glycerol and/or ethanol formation is also observed on fructose relative to glucose based medium. Taken together, these results suggest that in fructophilic Wickerhamiella species fructose is more conducive to fermentation than glucose and that this effect seems to be more pronounced for FFZ1 harboring species.
Fig. 5
Growth on fructose promotes fermentation. Comparison of global fermentation byproduct formation (ethanol, mannitol, and glycerol together), in the form of global maximum yields, in fructophilic Wickerhamiella species possessing or lacking FFZ1 and cultivated on either glucose (20G medium) or fructose (20F medium) as sole carbon source, as indicated below the plot. Each datapoint represents a single measurement (replicate experiment can be found in Additional file 6: Table S2 and Additional file 8: Fig. S6C)
留言 (0)