
記住我
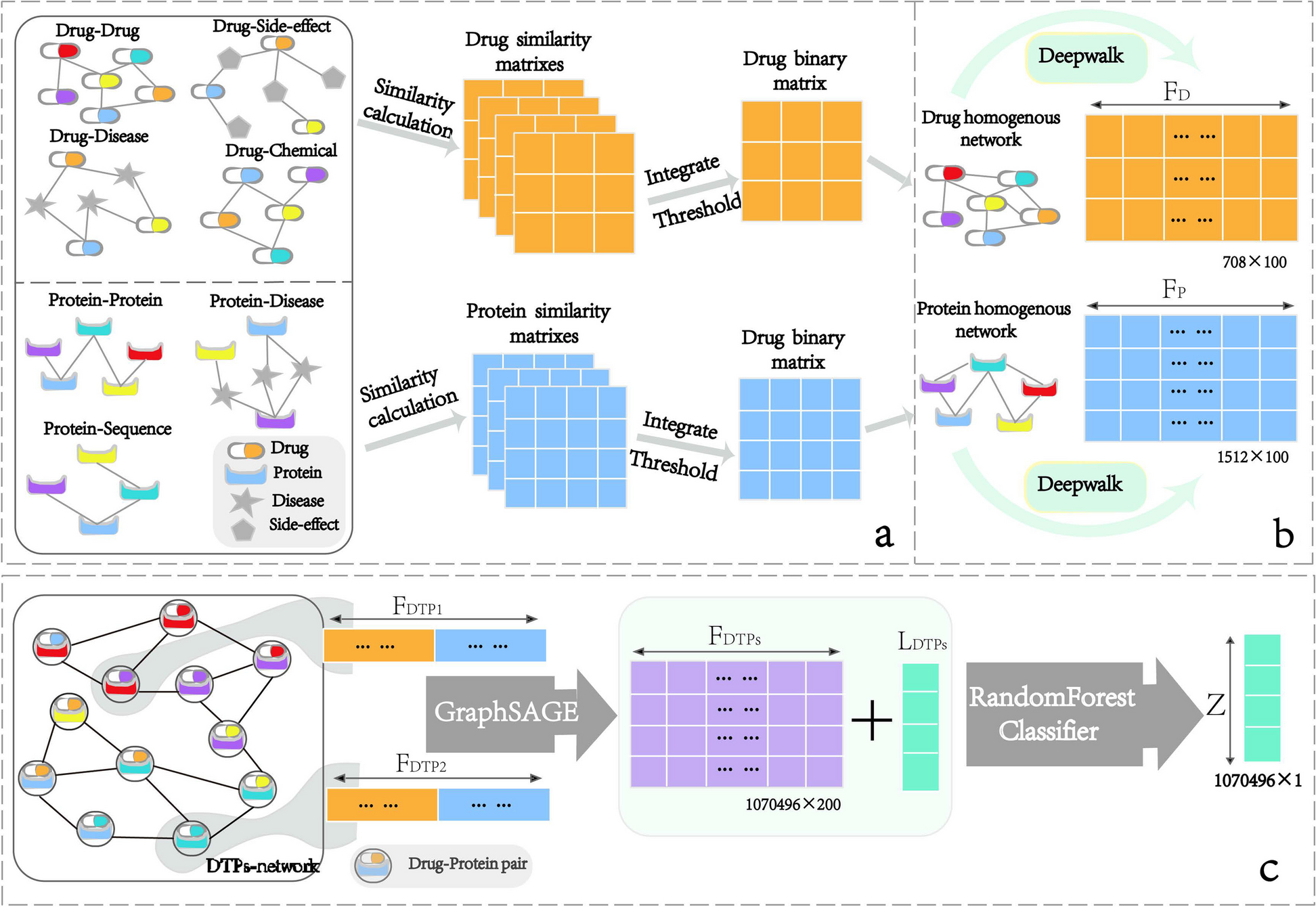


To directly assess the possibility that Grb10 interacts with the Igf1r to influence growth we performed genetic crosses between both Grb10Δ2-4 and Grb10ins7 (collectively referred to as Grb10 KO strains) and Igf1r KO mice. Grb10Δ2-4 offspring were analysed at PN1 and e17.5 whereas Grb10ins7 offspring were analysed at PN1 only. To increase statistical power, both sexes were pooled together and considered in a single analysis, with mean weights ± standard error of the mean stated in the text and shown graphically for offspring genotype groups. PN1 data were consistent between offspring of the two Grb10 KO strains (as summarised in Table 1) and consequently all subsequent experiments were carried out with only the Grb10Δ2-4 strain.
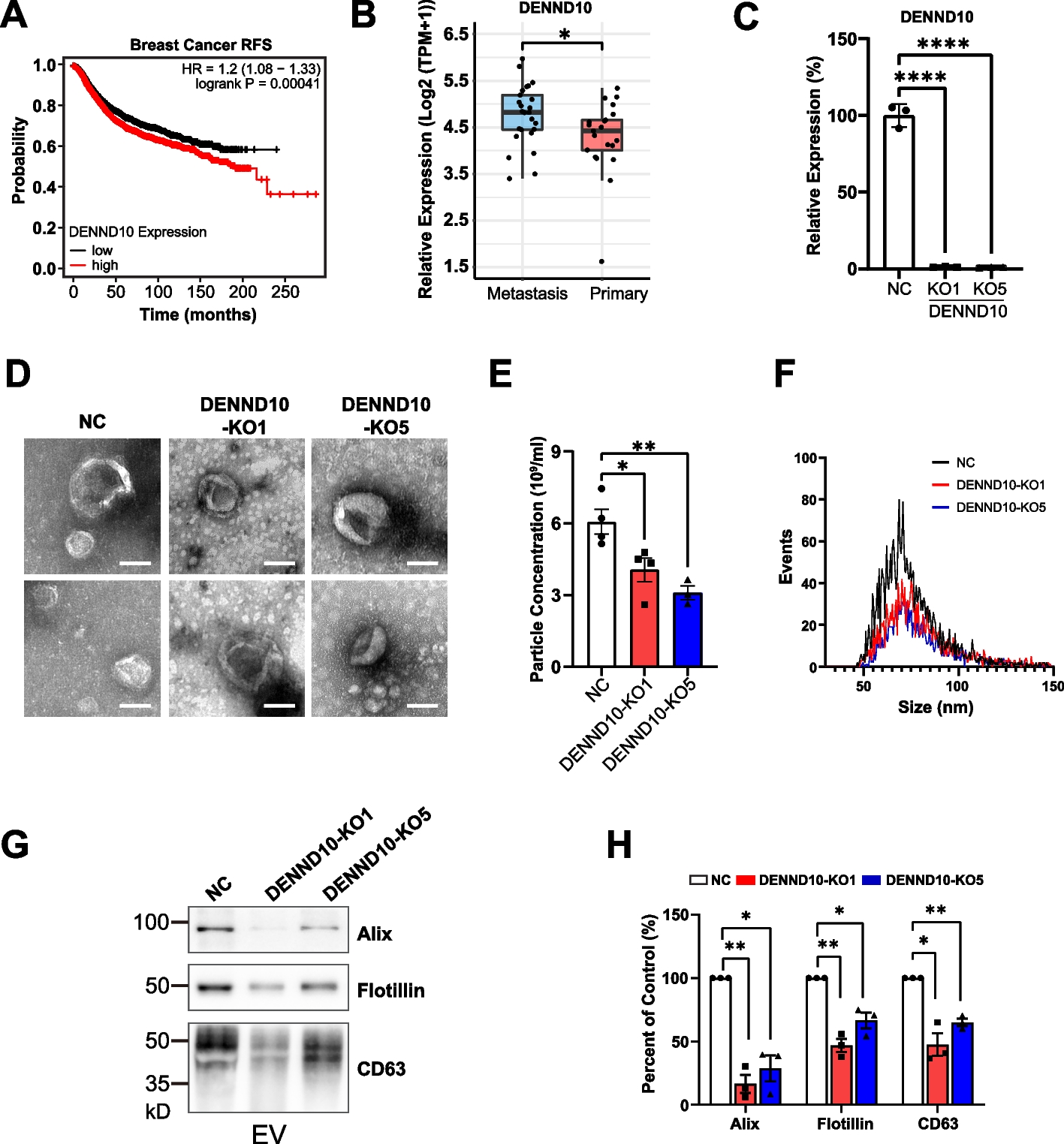
Table 1 Summary of PN1 body and organ weight data for progeny of crosses between Grb10 KO strains and Igf1r KO mice. Mean weights are shown for each genotype together with changes relative to wild type (%WT) for each mutant genotype. A) Grb10ins7 KO data. B) Grb10Δ2-4 KO data Grb10ins7 KO x Igf1r KO offspring PN1 body massProgeny of crosses between Grb10ins7+/p:Igf1r+/- females and Grb10ins7+/+:Igf1r+/- males were collected at PN1 for body and organ weight analysis (Fig. 2). Progeny with six genotypes were reduced to four groups by pooling Grb10ins7+/+:Igf1r+/- with Grb10ins7+/+:Igf1r+/+ (wild type group) and Grb10ins7m/+:Igf1r+/- and Grb10in7m/+:Igf1r+/+ (Grb10ins7 KO group), for comparison with the Igf1r KO and Grb10ins7:Igf1r DKO groups (Table 2A). This was done following initial analysis of the data which confirmed that Igf1r+/- animals had a normal fetal growth phenotype (Additional file 1: Fig.S1), as previously shown [41]. Pooling allowed us to strengthen statistical analyses, while simplifying data analysis and presentation, without materially affecting the outcome. If Grb10 regulates growth through an interaction with the Igf1r, Grb10:Igf1r DKO animals would be expected to be phenotypically indistinguishable from Igf1r KO animals (Fig. 1B). Body mass data (Fig. 2A; Table 1A) immediately indicated that we should reject this hypothesis. Grb10ins7 KO pups (mean weight 1.7670±0.0360g) were approximately 26% larger (p<0.0001) and Igf1r KOs (0.6395±0.0267g) 54% smaller (p<0.01) than wild type controls (1.401±0.0297g), respectively, whereas Grb10ins7:Igf1r DKO mutants were intermediate in size (1.1650±0.0554g). Thus, Grb10ins7:Igf1r DKO pups displayed an additive effect of both parental genotypes, being significantly different from Grb10ins7 KO (p<0.0001) single mutants, but not from both Igf1r KOand wild type neonates (Fig. 2A). This was supported by a two-way ANOVA test which showed both Grb10 (p<0.0001) and Igf1r (p<0.0001) are significant factors affecting body weight, in opposite directions, but detected no interaction between the two genotypes (p=0.1017).
Fig. 2
Weights at PN1 from progeny of crosses between Grb10ins7 KO and Igf1r KO mice. Data were pooled into four groups for analysis as described in the Methods, wild type, Igf1r KO, Grb10 KO and Grb10:Igf1r double knockouts (DKO). Body weights are shown for the four offspring genotype groups (A). Actual weights of brain (B), liver (C), lungs (D), heart (E) and kidneys (F) are shown alongside relative weights of the same organs, expressed as a percentage of body mass (G-K). Values represent means and SEM, tested by one-way ANOVA using Kruskal-Wallis and Dunn’s post hoc statistical tests. Summaries of Two-way ANOVA outcomes beneath each graph show the percentage of total variation (%var) and a p value for each source, namely the two single KO genotypes and any interaction (Inter.) between the two (values significant at p<0.05 in bold). Sample sizes were, for body, wild type (WT) n=38, Igf1r KO n=7, Grb10 KO n=26, Grb10:Igf1r DKO n=12; brain, WT n=38, Igf1r KO n=3, Grb10 KO n=25, Grb10:Igf1r DKO n=8; liver, WT n=38, Igf1r KO n=2, Grb10 KO n=25, Grb10:Igf1r DKO n=7; lungs, WT n=38, Igf1r KO n=7, Grb10 KO n=12, Grb10:Igf1r DKO n=7; heart, WT n=37, Igf1r KO n=2, Grb10 KO n=8, Grb10:Igf1r DKO n=7; kidneys, WT n=38, Igf1r KO n=2, Grb10 KO n=25, Grb10:Igf1r DKO n=7. Asterisks indicate p-values, *p <0.05, **p <0.01, ***p <0.001, ****p<0.0001
Table 2 Genetic crosses used in the study, showing parent and offspring genotypes with their expected Mendelian ratios. A) crosses between either Grb10 KO strain, (Grb10Δ2-4 and Grb10ins7) and the Igf1r KO strain. For statistical analysis Igf1r+/- heterozygous offspring were grouped with their respective Igf1r+/+ wild type counterparts, as indicated. B) Crosses between the Grb10Δ2-4 KO and Insr KO strains. For statistical analysis Insr+/- heterozygous offspring were grouped with their respective Insr+/+ wild type counterparts, Grb10+/p with respective Grb10+/+ wild types and Grb10m/p with respective Grb10m/+ as Grb10 maternal allele knockouts, as indicated. DKO = double knockoutGrb10ins7 KO x Igf1r KO offspring PN1 organ massTo assess body proportions selected individual organs (brain, liver, lungs, heart, kidneys) were dissected at PN1 and their weights were analysed directly (Fig. 2B-F) and as a percentage of total body weight (Fig. 2G-K). The pattern of organ weight difference across the genotypes was again consistent with the DKO pups having an additive phenotype, comprising the sum of the two single KO phenotypes (summarised in Table 1A). First, the brain from Grb10ins7 KO (mean mass 0.0860±0.0017g) pups was spared from the general overgrowth phenotype indicated by body mass and was only 4% larger than wild type brain (0.0824±0.0019g) (Fig. 2B). Meanwhile, brains from Igf1r KO (0.0491±0.0021g) and Grb10ins7:Igf1r DKO (0.0535±0.0012g) pups were strikingly similar, being smaller than wild type brain by 40% (p<0.05 ), and 35% (p<0.001), respectively. Thus, while Igf1r KO brains were roughly proportionate with body size, both Grb10ins7 KO (p<0.0001) and Grb10ins7:Igf1r DKO (p<0.0001) brains were disproportionately small within larger bodies (Fig. 2G). In other words, the Grb10ins7:Igf1r DKO phenotype was dominated by brain size being severely reduced, as in Igf1r KO pups, which can therefore be attributed to loss of Igf1r expression. In keeping with this Two-way ANOVA indicated that brain weight was influenced mainly by Igf1r (p<0.0001).
In direct contrast, the livers of Grb10ins7 KO (0.1231±0.0051g) and Grb10ins7:Igf1r DKO (0.1115±0.0083g) pups were each at least double, by 124% (p<0.0001) and 103% (p<0.01), respectively, the size of wild type (0.0550±0.0016g), while the Igf1r KO (0.0469±0.0036g) liver was some 15% smaller (Fig. 2C). Consequently, while the liver was disproportionately enlarged within the heavier Grb10ins7 KO body (p<0.0001), liver disproportion was exaggerated in Grb10ins7:Igf1r DKO (p<0.0001) pups, due to DKOs having a body size similar to wild type (Fig. 2H). Due to their greatly reduced body mass relative to wild types, although Igf1r KO livers were smaller in actual mass than in wild type controls, Igf1r KO pups also had disproportionately large livers. Although neither actual nor relative liver weight was significantly different between Igf1r KO and wild type (likely due to the small Igf1r KO small sample size), the Grb10ins7:Igf1r DKO liver weight phenotype was clearly dominated by the massive size increase also seen in Grb10 KO single mutants and therefore associated with loss of the maternal Grb10ins7 allele. Two-way ANOVA analysis reflected this with only Grb10 significantly (p<0.0001) contributing to liver weight.
The remaining organs followed a pattern of size difference like that seen in the body mass data, in that Grb10ins7:Igf1r DKO mass was intermediate between that of the two single KO values. Compared to wild type (0.0387±0.0014g) lungs from a single Igf1r KO sample (0.0079g) were 80% lighter (not statistically significant due to very small samples size) and Grb10ins7 KO (0.0480±0.0019g) 24% heavier (p<0.01), whereas Grb10ins7:Igf1r DKO (0.0287±0.0013g) lungs were 26% smaller (p<0.05) and intermediate in size (Fig. 2D). Relative to total body mass, Igf1r KO lungs appeared disproportionately small while Grb10ins7 KO and Grb10ins7:Igf1r DKO lungs were roughly proportionate with their respective body sizes (F igure 2I). According to two-way ANOVA, both Grb10 (p<0.0024) and Igf1r (p<0.0001) contributed significantly to lung weight. Similarly, in comparison with wild type (0.0092±0.0005g), hearts from Igf1r KO (0.0078±0.0004g) pups were some 15% smaller and Grb10ins7 KO hearts (0.0127±0.0006g) 39% larger (p<0.0001), with Grb10ins7:Igf1r DKO hearts (0.0094±0.0006g), intermediate in size, being only 2% larger than wild type (Fig. 2E). While these weight differences were not all statistically significant, in relative terms, the heart from Grb10ins7 KO (p<0.05) and Igf1 KO (p<0.05) single mutants were disproportionately large, whereas the Grb10ins7:Igf1r DKO heart was not (Fig. 2J). Two-way ANOVA indicated Igf1r (p<0.0395) as the major contributor to heart weight.
In the case of kidneys, those from Grb10ins7 KO (0.0166±0.0006g) were only slightly enlarged, by 5%, compared with wild type (0.0159±0.0005g), while both Igf1r KO (0.0111±0.00134g) and DKO (0.01287±0.0009g), were smaller by 31% and 20%, respectively (Fig. 2F). The only significant difference in kidney weights was between Grb10ins7 KO and Grb10ins7:Igf1r DKO (p<0.05). Relative to wild type body mass, this meant that Grb10ins7 KO pups alone had disproportionately small kidneys (p<0.001) (Fig. 2K). Two-way ANOVA indicated Igf1r (p<0.001) as the major contributor to kidney weight. For each individual organ two-way ANOVA tests indicated there was no interaction between the genotypes, just as for the whole body (Fig. 2A-F).
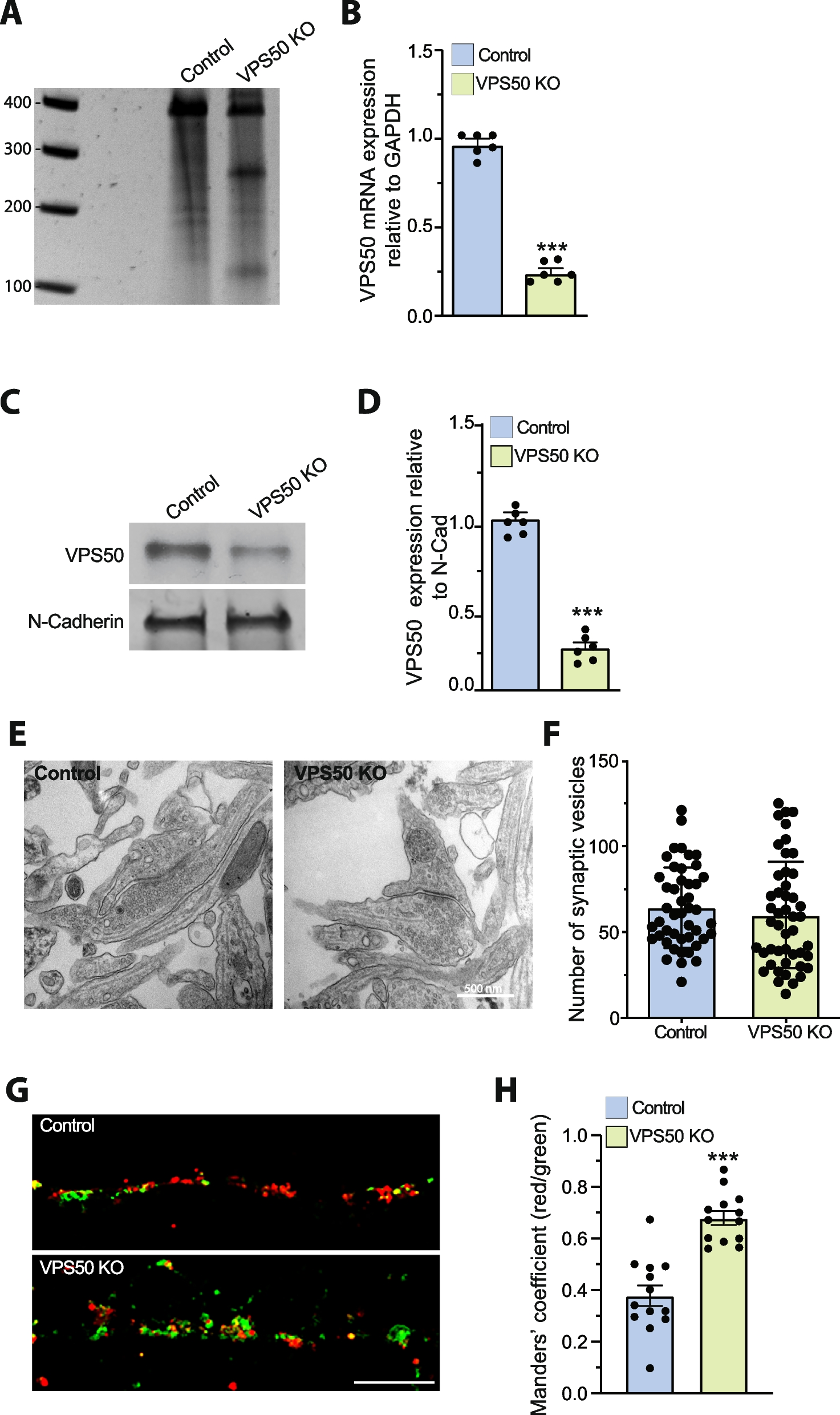
Grb10Δ2-4 KO x Igf1r KO offspring PN1 body massTo corroborate data from the Grb10ins7 strain, similar PN1 data were collected using the Grb10Δ2-4 strain. Progeny of crosses between Grb10Δ2-4+/p:Igf1r+/- females and Grb10Δ2-4+/+:Igf1r+/- males were again collected at PN1 and whole body weights recorded along with weights of selected organs (Fig. 3). As before, data for the six offspring genotypes were pooled to generate four groups for analysis, combining Grb10Δ2-4+/+:Igf1r+/- with Grb10Δ2-4+/+:Igf1r+/+ (wild type group) and Grb10Δ2-4m/+:Igf1r+/- with Grb10Δ2-4m/+:Igf1r+/+ (Grb10Δ2-4 KO group) progeny (Table 1B), which was again supported by our initial data analysis (Additional file 1: Fig. S2). As for the previous cross, while Grb10Δ2-4 KO pups (mean weight 1.887 ±0.0239g) were around 33% larger (p<0.0001) and Igf1r KOs (0.6205±0.0192g) 56% smaller (p<0.001), respectively, than wild type controls (1.422±0.0189g), Grb10Δ2-4:Igf1r DKO mutants (1.278±0.0381g) were intermediate in size, just 10% smaller than wild type (Fig. 3A; Table 1B). Grb10Δ2-4:Igf1r DKO pups were smaller than Grb10Δ2-4 KO pups (p<0.0001) but not significantly smaller than wild type neonates (Fig. 3A, B), while Grb10Δ2-4 KO pups were significantly larger than both wild type (p<0.0001) and Igf1r KO (p<0.0001) pups. The two-way ANOVA test showed both Grb10 (p<0.0001) and Igf1r (p<0.0001) contributed significantly to body weight and indicated a possible interaction between the genotypes, but at a relatively high significance level (p=0.0135). Despite this, it was clear that Grb10Δ2-4:Igf1r DKO pups were not small, to the extent consistently shown for Igf1r KO pups, and instead their intermediate size must result from an additive effect of the two mutant parental genotypes.
Fig. 3
Weights at PN1 from progeny of crosses between Grb10Δ2-4 KO and Igf1r KO mice. Data were pooled into four groups for analysis as described in the Methods, wild type, Igf1r KO, Grb10 KO and Grb10:Igf1r double knockouts (DKO). Body weights are shown for the four offspring genotype groups (A). Gross physical appearance of typical WT (Grb10+/+:Igf1r+/+) and Grb10:Igf1r DKO (Grb10-/-:Igf1r-/-) pups, noting in the DKO the small head relative to body size and enlarged liver (l) obscuring the milk filled stomach (s), clearly visible through the skin of the wild type (B). Actual weights of brain (C), liver (D), lungs (E), heart (F) and kidneys (G) are shown alongside relative weights of the same organs, expressed as a percentage of body mass (H-L). Values represent means and SEM, tested by one-way ANOVA using Kruskal-Wallis and Dunn’s post hoc statistical tests. Summaries of Two-way ANOVA outcomes beneath each graph show the percentage of total variation (%var) and a p value (values significant at p<0.05 in bold) for each source, namely the two single KO genotypes and any interaction (Inter.) between the two. Sample sizes were, for body wild type (WT) n=104, Igf1r KO n=13, Grb10 KO n=92, Grb10:Igf1r DKO n=28; brain, WT n=102, Igf1r KO n=6, Grb10 KO n=90, Grb10:Igf1r DKO n=24; liver, WT n=104, Igf1r KO n=5, Grb10 KO n=90, Grb10:Igf1r DKO n=23; lungs, WT n=104, Igf1r KO n=4, Grb10 KO n=90, Grb10:Igf1r DKO n=23; heart, WT n=103, Igf1r KO n=5, Grb10 KO n=88, Grb10:Igf1r DKO n=23; kidneys, WT n=100, Igf1r KO n=5, Grb10 KO n=90, Grb10:Igf1r DKO n=23. Asterisks indicate p-values, *p <0.05, **p <0.01, ***p <0.001, ****p<0.0001
Grb10Δ2-4 KO x Igf1r KO offspring PN1 organ massAs before, body proportions were assessed by dissecting and weighing selected organs at PN1. Organ weights were analysed directly (Fig. 3C-G) and as a percentage of total body weight (Fig. 3H-L). The genotype-dependent differences in organ weights were again consistent with the Grb10Δ2-4:Igf1r DKO pups having an additive phenotype in comparison with the two single KO genotypes (summarised in Table 1B). First, the brain from Grb10Δ2-4 KO (0.0931±0.001g) pups was spared from the general overgrowth phenotype indicated by body mass and was only 10% larger (p<0.0001) than wild type brain (0.0849±0.0011g) (Fig. 3C), Meanwhile, brains from Igf1r KO (0.058±0.0018g) and Grb10Δ2-4:Igf1r DKO (0.058±0.0012g) pups were strikingly similar, being smaller than wild type brain by 42%, (p<0.01) and 32% (p<0.0001), respectively. Thus, while Igf1r KO brains were proportionate to their small bodies, both Grb10Δ2-4 KO (p<0.0001) and Grb10Δ2-4:Igf1r DKO (p<0.0001) brains were small within larger bodies (Fig. 3H). In other words, the Grb10Δ2-4:Igf1r DKO phenotype was dominated by brain size being severely reduced, as in Igf1r KO pups, and is therefore associated with loss of Igf1r expression.
In contrast, the livers of DKO (0.1259±0.006g) and Grb10Δ2-4 KO (0.1279±0.0036g) pups were again each more than double (122% and 127% larger, respectively) the size of wild type (0.0568±0.0012g) liver (p<0.0001), while the Igf1r KO (0.0454±0.0016g) liver was some 20% smaller (Fig. 3D). Consequently, the liver was disproportionately enlarged within the heavier Grb10Δ2-4 KO body (p<0.0001), and in the Grb10Δ2-4:Igf1r DKO liver disproportion was exaggerated (p<0.0001), due to DKOs having a body size similar to wild type (Fig. 3I). Due to their greatly reduced body mass relative to wild types, although Igf1r KO livers were smaller in actual mass than in wild type controls, Igf1r KO pups also had disproportionately large livers (p<0.001). Similar to our findings using the Grb10ins7 KO strain, the Grb10Δ2-4:Igf1r DKO phenotype was clearly dominated by the massive size increase associated with loss of the maternal Grb10Δ2-4 allele.
The remaining organs followed a pattern of size differences like that seen in the body mass data, in that DKO mass was intermediate between that of the two single KO values. Lungs from Grb10Δ2-4:Igf1r DKO (0.0333±0.0018g) pups were only 16 % smaller than wild type (0.0398±0.0009g) but differed to those of both single mutants, with Igf1r KO (0.0085±0.0005g) approximately 79% lighter than wild type and Grb10Δ2-4 KO (0.0539±0.0012g) 35% heavier (p<0.0001) (Fig. 3E). Grb10Δ2-4 KO lung weight was significantly different to all three other genotypes (p<0.0001 in each case).. Relative to total body mass, Igf1r KO lungs were disproportionately small (p<0.01) while Grb10Δ2-4 KO lungs were roughly proportionate with their respective body sizes and Grb10Δ2-4:Igf1r DKO marginally (p<0.05), disproportionately small (Fig. 3J). Similarly, in comparison with wild type (0.009±0.0002g), hearts from Igf1r KO (0.0073±0.0005g) pups were some 18% smaller and Grb10Δ2-4 KO hearts (0.0134±0.0003g) 51% larger (p<0.0001) (Fig. 3F). Grb10Δ2-4:Igf1r DKO hearts (0.0112±0.0005g) were intermediate in size, being 26% larger (p<0.01) than wild type but not significantly different to Grb10 KO single mutants In relative terms, the hearts from Igf1r KO pups were proportionate and those of Grb10 KO (p<0.0001) and DKO (p<0.01) of Grb10 KO (p<0.0001) and DKO (p<0.01) disproportionately large compared to wild type controls. (Fig. 3K).
Compared to wild type kidneys (0.0157±0.0003g), Igf1r KO (0.0099±0.0009g) kidneys were reduced in size, by 36% (p<0.05), while Grb10Δ2-4 KO kidneys (0.0179±0.0004g) were larger by 14% (p<0.01) and Grb10Δ2-4:Igf1r DKO (0.0138±0.0006g) were intermediate, being 12% larger, but not significantly different to wild type (Fig. 3G). Notably, Grb10Δ2-4 KO kidney weights were still significantly different to those of Igf1r KO (p<0.001) and Grb10Δ2-4:Igf1r DKO (p<0.0001). Relative to body mass (Fig. 3L), this meant kidneys were proportionate in Igf1r KO pups, but disproportionately small in the larger body of Grb10Δ2-4 KO (p<0.0001) pups. As in the previous cross, two-way ANOVA tests for individual organs indicated there was no interaction between the genotypes in each case (Fig. 3C-G). In almost all cases both Grb10 and Igf1r contributed significantly to organ weight. The exception was liver where Grb10 (p<0.0001) was the major influence on weight and the influence of Igf1r did not reach significance.
The organ disproportion evident in Grb10Δ2-4:Igf1r DKO PN1 pups was reflected by their appearance (Fig. 3B). Despite being similar in size to wild types, Grb10Δ2-4:Igf1r DKO pups had small, flattened heads and livers that were distended such that they largely obscured the milk-filled stomach.
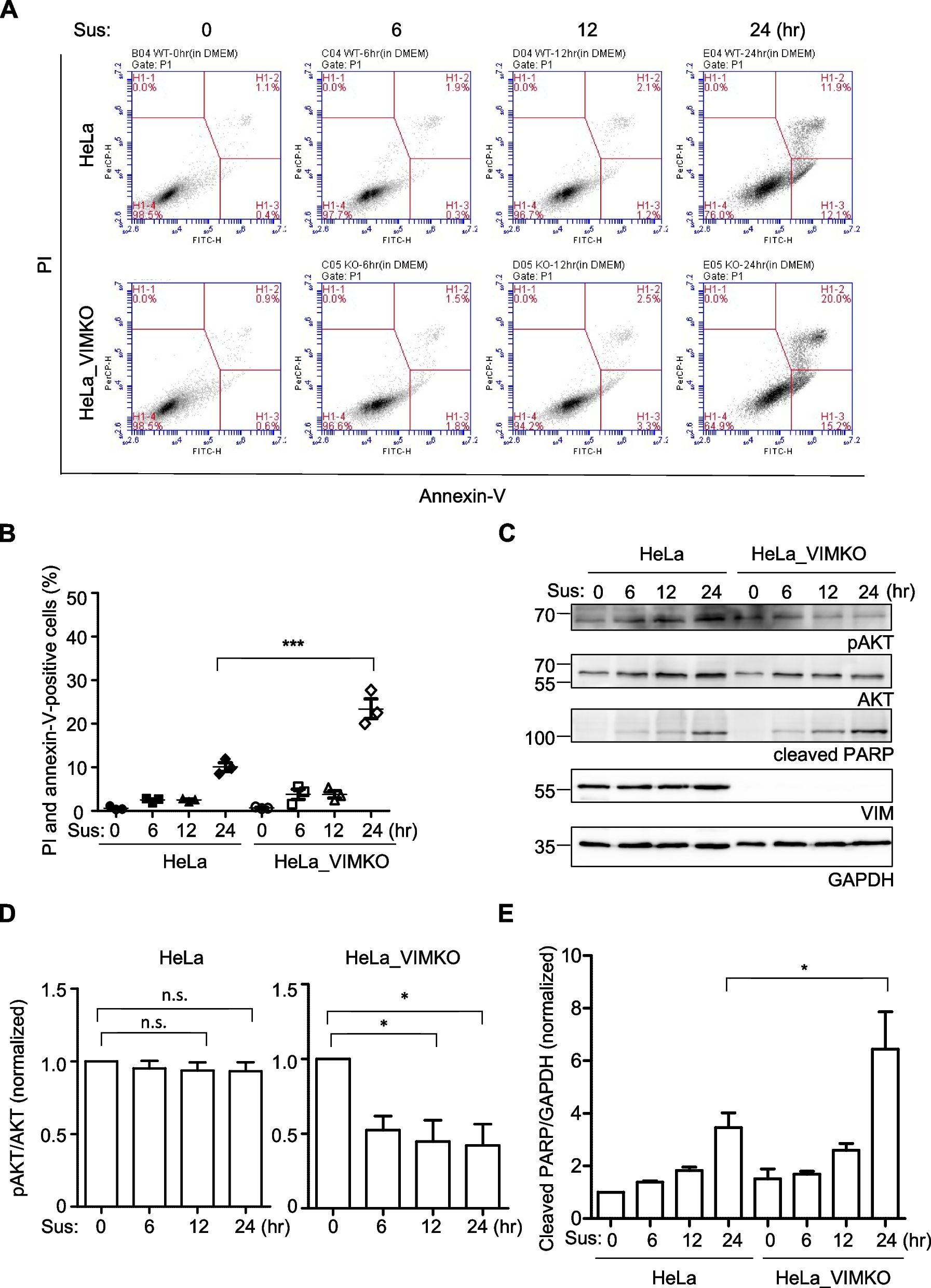
Grb10Δ2-4 KO x Igf1r KO offspring e17.5 embryo and placentaTo investigate the potential for interaction between Igf1r and Grb10 to regulate growth by acting within the placenta we analysed weights of the whole embryo and placenta at e17.5 (Fig. 4). We chose a time-point late in gestation when any size differences between conceptuses of different genotypes would be relatively large. The pattern of size differences observed was very similar to that seen for pups at PN1. Grb10Δ2-4 KO embryos (1.085±0.0450g) were 35% larger than wild type (0.8031±0.0371g), whereas the single Igf1r KO (0.4029g) embryo collected was 50% smaller and Grb10Δ2-4:Igf1r DKO embryos (0.6330±0.0286g) intermediate in size, at 21% lighter than wild types (Fig. 4A). Unsurprisingly, the one Igf1r KO embryo showed no statistical differences in size compared to any of the other genotypes, however, Grb10Δ2-4 KO embryos were significantly larger than wild type (p<0.05) and Grb10Δ2-4:Igf1r DKO (p<0.0001) embryos.
Fig. 4
Weight analysis of e17.5 conceptuses from crosses between Grb10Δ2-4 KO and Igf1r KO mice. Data were pooled into four groups for analysis as described in the Methods, wild type, Igf1r KO, Grb10 KO and Grb10:Igf1r double knockouts (DKO). Weights are shown for the four offspring genotype groups for embryo (A) and placenta (B) and these have been used to calculate the embryo:placenta weight ratio as a measure of placental efficiency (C). Values represent means and SEM, tested by one-way ANOVA using Kruskal-Wallis and Dunn’s post hoc statistical tests. Summaries of Two-way ANOVA outcomes beneath each graph show the percentage of total variation (%var) and a p value (values significant at p<0.05 in bold) for each source, namely the two single KO genotypes and any interaction (Inter.) between the two. Sample sizes were, for wild type (WT) n=17, Igf1r KO n=1, Grb10 KO n=11, Grb10:Igf1r DKO n=9. Asterisks indicate p-values, *p <0.05, **p <0.01, ****p<0.0001
Placental weights followed a similar pattern (Fig. 4B), with Grb10Δ2-4 KO (0.1073±0.0060g) 22% larger than wild type (0.0882±0.0036g), the single Igf1r KO placenta (0.0729g) 17% smaller and Grb10Δ2-4:Igf1r DKO placentas (0.0916±0.0040g) in between at only 4% larger. The only statistically significant difference was between wild type and Grb10Δ2-4 KO samples (p<0.05). Next, the ratio of embryo to placental mass was calculated for each genotype as an estimate of placental efficiency (Fig. 4C). Although not statistically significant, the trend was for Grb10Δ2-4 KO placental efficiency (10.41) to be slightly higher than wild type (9.39), while both Igf1r KO (5.53) and Grb10Δ2-4:Igf1r DKO (6.98) were lower than wild type, with the only significant difference between Grb10Δ2-4 KO and Grb10Δ2-4:Igf1r DKO (p<0.01). A two-way ANOVA test found no evidence of an interaction between the genotypes for either embryo or placenta size (Fig. 4A,B).
Survival of Grb10 KO x Igf1r KO progeny at PN1 and e17.5During collection of offspring the small, presumptive Igf1r KO pups seemed scarce and chi-square tests of observed versus expected numbers generally supported this notion (Additional file 2: Table S1). Testing of PN1 data from the Grb10Δ2-4 KO x Igf1r KO cross, which had the largest sample size (n=237), indicated that paucity of Igf1r KO pups was statistically significant (p=0.0174; Additional file 2: Table S1A), with 44% of the expected numbers surviving. The same was true for offspring collected from the same cross at e17.5 (p=0.015), though in this case the sample size was lower (n=38) and only one Igf1r KO embryo was obtained, with the expected number being closer to 5 (Additional file 2: Table S1B). In the case of the Grb10ins7 x Igf1r KO PN1 dataset (n=83), the lack of Igf1r KO pups was less evident (67% of the expected number) and the chi-square test indicated no significant deviation from expected mendelian ratios (p=0.4711; Additional file 2: Table S1C). In both crosses it was clear that Igf1r KO pups found alive on the day of birth were failing to thrive, as previously reported [42]. Strikingly, this did not appear to be true for Grb10:Igf1r DKO PN1 pups in either cross which typically had milk-filled stomachs, appeared to be doing well on PN1 and were not underrepresented (Additional file 2: Table S1).
Genetic interaction tests show that Grb10 inhibits fetal growth largely independently of the Insr, except in liver, where excessive enlargement in Grb10 KO neonates is due to Insr-mediated lipid accumulationGrb10Δ2-4 KO x Insr KO offspring PN1 body massTo address the question of whether Grb10 regulates growth in vivo through an interaction with the Insr, we next performed intercrosses between Grb10Δ2-4+/p:Insr+/- double heterozygous mice, giving rise to twelve offspring genotypes, which were reduced to four groups for analysis (Table 2B). In addition to combining animals with Insr+/- and Insr+/+ genotypes (Insr wild type groups), we also pooled Grb10Δ2-4+/+ with Grb10Δ2-4+/p genotypes (Grb10 wild type) and Grb10Δ2-4m/+ with Grb10Δ2-4m/p (Grb10 KO). This is because the Grb10 paternal allele is silent in the majority of tissues and its knockout is well established to have no effect on fetal growth [24,25,26, 49]. Similarly, only Insr-/- animals have been shown to have a mutant phenotype affecting either growth or glucose regulation [43, 45, 50]. Initial analysis of our data prior to pooling was in line with these earlier studies (Additional file 1: Fig. S3). As asserted in the case of the Igf1r, should Grb10 regulate growth through an interaction with the Insr, Grb10Δ2-4:Insr DKO animals would be phenotypically indistinguishable from Insr KO single mutants (Fig. 1B).
Progeny were first collected at PN1 for body and organ weight analysis (Fig. 5). Just like the crosses involving the Igf1r KO, body mass data (Fig. 5A, Table 3) indicated that we should reject this hypothesis for crosses involving the Insr. Insr KO pups (1.2680±0.0483g) were not significantly different to wild type controls (1.3440±0.0297g), being only 6% smaller. In contrast, both Grb10Δ2-4 KO (1.8140±0.0447g) and Grb10Δ2-4:Insr DKO (1.6990±0.0853g) pups were substantially larger than wild type, by 35% (p<0.0001) and 26% (p<0.05), respectively, but not significantly different to each other. Thus, the overgrowth associated with loss of the maternal Grb10 allele is maintained in DKO pups despite loss of Insr expression. A two-way ANOVA test supported this, showing that body weight was mostly driven by Grb10 (p<0.0001) with little influence from Insr, and no evidence of an interaction between the genotypes (Fig. 5A).
Fig. 5
Analyses of PN1 progeny from crosses between Grb10Δ2-4 KO and Insr KO mice. Data for numerical analyses were pooled into four groups for analysis as described in the Methods, wild type (WT), Insr KO (IKO), Grb10 KO (GKO) and Grb10:Insr double knockouts (DKO). Body weights are shown for the four offspring genotype groups (A). Actual weights of brain (B), liver (C), lungs (D), heart (E) and kidneys (F) are shown alongside relative weights of the same organs, expressed as a percentage of body mass (G-K). Values represent means and SEM, tested by one-way ANOVA using Kruskal-Wallis and Dunn’s post hoc statistical tests. Summaries of Two-way ANOVA outcomes beneath each graph show the percentage of total variation (%var) and a p value (values significant at p<0.05 in bold) for each source, namely the two single KO genotypes and any interaction (Inter.) between the two. Sample sizes were, wild type (WT) n=42, Insr KO n=6, Grb10 KO n=44, Grb10:Insr DKO n=9. Histological sections of liver, stained with haematoxylin and eosin, are shown at 100x magnification for WT (L), Insr KO (M), Grb10 KO (N) and Grb10:Insr DKO (O) mice, and at 300x magnification for the same animals (L’-O’). Images are representative of at least three biological replicates per genotype and were taken at 100x magnification (scale bars show 50μm for the lower power images and 20μm for the higher power images). Asterisks indicate p-values, *p <0.05, **p <0.01, ***p <0.001, ****p<0.0001
Table 3 Summary of PN1 body and organ weight data for progeny of crosses between the Grb10Δ2-4 KO strain and Insr KO mice. Mean weights are shown for each genotype together with changes relative to wild type (%WT) for each mutant genotypeGrb10Δ2-4 KO x Insr KO offspring PN1 organ massAs for the earlier crosses involving Igf1r KO strains, the same selection of organs was collected and weighed at PN1 to evaluate body proportions of offspring involving the Insr KO. Organ weights were analysed directly (Fig. 5B-F) and as a percentage of total body weight (Fig. 5G-K). The patterns of weight differences displayed across the genotypes was consistent with the Grb10Δ2-4:Insr DKO pups having an additive phenotype compared with the two single KOs (summarised in Table 3). The brain from Grb10Δ2-4 KO (0.0918±0.0014g) pups was once again largely spared from the general overgrowth phenotype indicated by body mass, being only 9% larger than wild type (0.0841±0.0014g), which was a significant difference (p<0.001) in this cross (Fig. 5B). Brains from Grb10Δ2-4:Insr DKO (0.0930±0.0044g) pups were similarly some 11% larger than wild type, whereas Insr KO brains (0.0856±0.0039g) were almost indistinguishable at only 2% larger. This meant that Grb10Δ2-4 KO and Grb10Δ2-4:Insr DKO brains were disproportionately small within larger bodies (Fig. 5G), compared with wild type (p<0.0001 and p<0.05, respectively) and Insr KO (p<0.0001 and p<0.05) brains. Thus, Grb10Δ2-4:Insr DKO brain size followed the pattern of the Grb10Δ2-4 KO and not the Insr KO single mutant phenotype. This interpretation is supported by two-way ANOVA which showed Grb10 (p<0.0074), but not Insr to be a significant influence on brain weight.
Liver displayed a particularly interesting pattern of weight differences (Fig. 5C). Wild type (0.0533±0.0017g) and Insr KO (0.0475±0.0036g) liver sizes were very similar, while Grb10Δ2-4 KO (0.1112±0.0049g) liver was more than twice normal size, at 109% larger than wild type (p<0.0001), as seen in the previous crosses. However, in this case Grb10Δ2-4:Insr DKO liver (0.0622±0.0023g) was only 17% larger than wild type and was significantly different to Grb10Δ2-4 KO liver size (p<0.05) but not to wild type or Insr KO liver, indicating that the disproportionate liver overgrowth associated with loss of Grb10 expression was largely Insr-dependent. This conclusion was reinforced by the finding that only Grb10Δ2-4 KO liver was disproportionately enlarged, in comparison with wild type (p<0.0001), Insr KO (p<0.001) and Grb10Δ2-4:Insr DKO (p<0.0001) (Fig. 5H). Further, a two-way ANOVA test found an interaction between the genotypes for liver weight (p=0.0013) but not for any other organ (Fig. 5A-F). To investigate the liver phenotype further we carried out histological analysis and found that the accumulation of excess lipid previously observed in neonatal Grb10Δ2-4 KO pups [26] was abrogated in Grb10Δ2-4:Insr DKO pups. Viewed at lower magnification (100x), the enlargement of hepatocytes through excess lipid storage was seen throughout Grb10Δ2-4 KO (Fig. 5N), but not wild type (Fig. 5L), Insr KO (Fig. 5M) or Grb10Δ2-4:Insr DKO (Fig. 5O) liver sections. A degenerate fatty histopathological phenotype, that has previously been described in neonatal liver of Insr KO homozygotes [45, 50], is seen more clearly at higher magnification (300x) (Fig. 5M’). This was also evident in Grb10Δ2-4:Insr DKO (Fig. 5O’) sections, is distinct from the lipid engorged cellular phenotype of Grb10Δ2-4 KO liver (Fig. 5N’) and absent in wild type sections (Fig. 5L’). Thus, the disproportionate hepatic overgrowth in Grb10 KO neonates was due to Insr signalling-dependent lipid deposition.
Lungs and heart followed a pattern of size differences like that of body mass. Grb10Δ2-4 KO (0.0442±0.0015g) and Grb10Δ2-4:Insr DKO (0.0431±0.0034g) lungs were similar in size, being 31% (p<0.0001) and 28% larger, respectively than wild type (0.0337±0.0013g), whereas Insr KO (0.0350±0.0011g) lungs were only 4% larger (Fig. 5D). Lungs from animals of all four genotypes remained proportionate with body weight (F igure 5I). Similarly, Grb10Δ2-4 KO (0.01378±0.0004g) and Grb10Δ2-4:Insr DKO (0.0112±0.0006g) hearts were both larger than wild type (0.0091±0.0002g) hearts by 51% (p<0.0001) and 23%, respectively, while Insr KO (0.0087±0.0006g) hearts were 4% smaller and indistinguishable from wild type (Fig. 5E). Hearts from animals of all four genotypes were proportionate with body weight (Fig. 5J). In this cross, Grb10Δ2-4 KO (0.0163±0.0005g) kidneys were 14% larger than wild type (0.0143±0.0004g) (Fig. 5F) but remained disproportionately small (p<0.0001) (Fig. 5K). Conversely, Insr KO (0.0163±0.0013g) kidneys were 14% larger than wild type and disproportionately large. Grb10Δ2-4:Insr DKO (0.0173±0.0013g) kidneys were 21% larger than wild type controls and roughly proportionate such that relative to body mass they were intermediate between the two single KOs. This once again reinforced the sparing of kidneys from the general overgrowth associated with loss of the maternal Grb10 allele.
Grb10Δ2-4 KO x Insr KO offspring e17.5 embryo and placentaWe next investigated the potential for interaction between Insr and Grb10 within the placenta by analysing weights of the whole embryo and placenta at e17.5 (Fig. 6). Similar to pups at PN1, compared to wild types (0.9245±0.0240g), Insr KO (0.8034±0.0569g) embryos were 13% smaller, though not significantly so, whereas Grb10Δ2-4 KO embryos (1.3010±0.0445g) and Grb10Δ2-4:Insr DKO embryos (1.2130±0.0741g) were larger, by 41% (p<0.0001) and 31%, respectively (Fig. 6A). This meant Grb10Δ2-4 KO (p<0.0001) and Grb10Δ2-4:Insr DKO (p<0.05) embryos were both significantly larger than Insr KO embryos but not different from each other.
Fig. 6
留言 (0)