
記住我
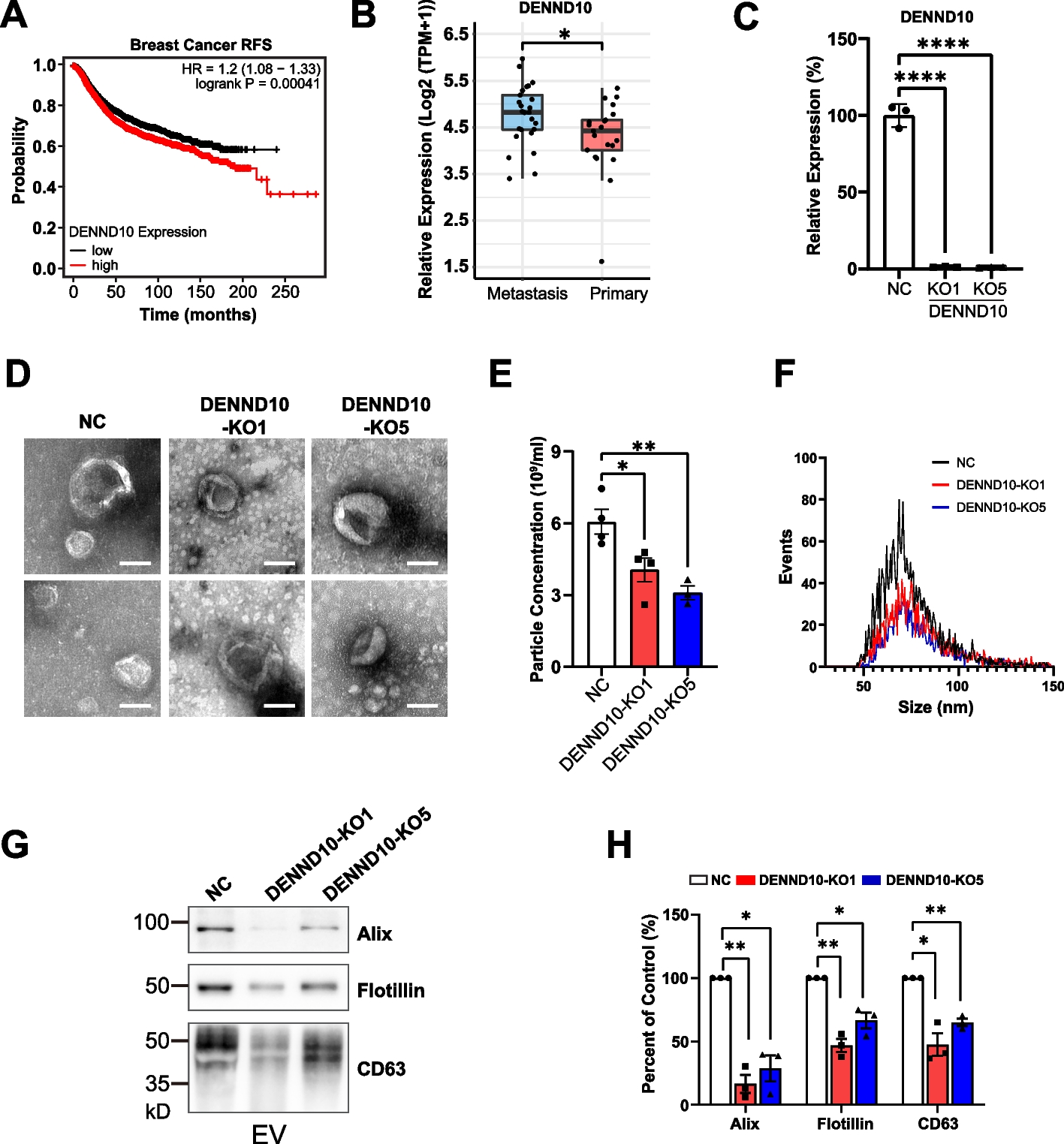


All groups of zQ175dn and Q175FDN mice and their respective WT controls completed a single one-hour open field trial (Fig. 1A) to assess naturalistic behaviour as mice explore a novel space. Manifest male and female zQ175dn mice showed a deficit in open field habituation, as indicated by a smaller reduction in distance traveled over time (Fig. 1B, C, E; Additional file 1: Fig S1A, B), while male manifest Q175FDN mice exhibited a unique pattern of behaviour in the open field, with significantly lower distance traveled in the first 10 min of the trial which then rose to WT levels for the final 50 min (Fig. 1D, E). Premanifest male and female zQ175dn mice exhibited similar distance traveled to WT littermates (Additional file 1: Figure S1C). Other measures such as percentage of time at center did not show a difference between genotypes for any groups (manifest zQ175dn: 17.0 ± 1.6 percent, WT littermates: 15.6 ± 1.6 percent, p = 0.5225; premanifest zQ175dn: 19.8 ± 2.2 percent, WT littermates: 18.1 ± 1.8 percent, p = 0.5415 2-way ANOVA; males and females pooled).
Fig. 1
Total distance traveled and percentage change in distance traveled per 5-min intervals during a single 60-min open field trial in manifest zQ175dn and Q175FDN compared to wild-type (WT) littermates. A) Example of mouse tracking of the base of the tail in the open field for 60 min. B) Total distance traveled per 5-min interval in the open field for manifest zQ175dn mice (males and females combined). C) Percentage change in distance traveled compared to the first 5-min interval in the open field for manifest zQ175dn mice (males and females combined). D) Total distance traveled per 5-min interval in the open field for male manifest Q175FDN mice. E) Distance traveled in the first 5 min (5 min) and last 5 min (60 min) for male manifest zQ175dn mice and WT littermates (left panel; genotype effect, last 5 min, p = 0.0141*) and Q175DFN mice and WT littermates (right panel; genotype effect, first 5 min, p = 0.0014**). One-way or two-way analysis of variance was used for all statistical analysis Asterisks (*) denote significance level. Individual values for groups with n < 6 are provided in Additional file 7: Individual values. ns = not significant. M = Male
We used the unsupervised machine learning algorithm B-SOiD [14] to further investigate how behaviour is affected in the open field. Using this method, we were able to reliably detect occurrences of locomotion (walking forward), turning left, turning right, sniffing, and rearing behaviours (Fig. 2, Additional file 2: Figure S2). Locomotion and turning behaviours were combined in our analysis as mice generally alternated between these behaviours. All groups spent the largest proportion of time in locomotion and turning behaviours (range of means for groups: 30 – 37% of the hour) and sniffing behaviours (range of means: 18 – 32% of the hour) with no genotype differences (Additional file 2: Figure S2A, B [top panels]). Rearing (away from the wall) accounted for a smaller proportion of total time (range of means: 3 – 8% of the hour), with male manifest zQ175dn mice showing increased rearing compared to WT littermates (Fig. 2A). Over time in the open field session, male manifest zQ175dn increased their total percentage of time rearing (Fig. 2B) and showed a longer average duration for each bout of rearing (Fig. 2C) compared to WT littermates.
Fig. 2
Engagement in rearing behaviour (away from the wall) during a single 60-min open field trial in Q175 mice compared to WT littermates. A Percentage of time spent rearing away from the wall (overall effect, p = 0.0117*; genotype effect for male manifest zQ175dn vs. WT littermates, p = 0.0031**; one-way ANOVA with multiple comparisons). B Percentage of time spent rearing in 5-min intervals over the 60-min trial in male manifest zQ175dn mice (mixed effects analysis). C Average duration of each bout of rearing per mouse in male manifest zQ175dn mice (genotype effect, p = 0.0154*; unpaired t-test). Asterisks (*) denote significance level. Individual values for groups with n < 6 are provided in Additional file 7: Individual values. ns = not significant. M = Male. F = Female
Although rearing in male manifest zQ175dn was the only behaviour to show a significant genotype difference when we analyzed total engagement in this behaviour, other differences appeared when we explored engagement in behaviours over time. Over the hour in the open field, WT littermates for all groups showed a reduction in locomotion and turning behaviours (Additional file 2: Figure S2A). Female manifest zQ175dn also showed a reduction, though smaller, in this behaviour over time, with a significant interaction between time and genotype for locomotion and turning, while males did not (Additional file 2: Figure S2A). Notably, male manifest Q175FDN mice engaged in locomotion and turning less during the first 10 min in the open field, and engaged in sniffing behaviours more, compared to WT littermates (Additional file 2: Figure S2A, B), which may indicate increased anxiety in these mice. Male manifest zQ175dn mice showed a decrease in sniffing over the hour while WT littermates showed a slight increase in sniffing; female manifest and both pre-manifest zQ175dn groups showed no difference in sniffing behaviour (Additional file 2: Figure S2B).
Altogether, our open field behavioural analysis revealed differences in behaviour patterns of Q175 mice that differ by sex and background strain.
Rotarod performance is affected by protocol and animal weightThe rotarod was used to assess motor learning and kinematics in zQ175dn mice. We tested two protocols for the accelerating rotarod task, which we called “Rotations Allowed” and “Standard” (described in methods) in the male manifest zQ175dn mice and their WT littermates. We found no difference in time on rotarod or time to first rotation between zQ175dn mice and WT on the Rotations Allowed protocol (Additional file 3: Figure S3A). Surprisingly, we also found no change in time on rotarod or time to first rotation over days of training for either genotype (Additional file 3: Figure S3A), indicating that animals were not improving with training on this version of the task. Consequently, we did not continue with the Rotations Allowed protocol for other groups. On the Standard protocol, all groups tested improved in time on rotarod over days of training, and interestingly, male manifest zQ175dn mice performed significantly better than their WT littermates, while female manifest zQ175dn mice performed equally well to littermates (Additional file 3: Figure S3C, D).
In light of these puzzling results, we turned our attention to animal weight and its effects on rotarod performance. Our male manifest zQ175dn mice were on average 10.48 ± 2.09 g (g) lighter than WT littermates (genotype: p < 0.0001****; zQ175dn range: 28.5—45.4 g; WT range: 30.9—51.7 g), and female manifest zQ175dn mice were 7.205 ± 2.17 g lighter than WT littermates (genotype: p = 0.0027**; zQ175dn range: 24.7—48.3 g; WT range: 33.6—45.4 g; Fig. 3A). Both male and female WT mice showed a significant inverse correlation between time on rotarod and weight on day 1 of training, while manifest zQ175dn mice of either sex did not show this correlation (Fig. 3A). After normalizing for weight (see methods), we found that male manifest zQ175dn mice showed a modest yet significant reduction in time on rotarod compared to WT littermates on the Rotations Allowed protocol (Additional file 3: Figure S3B). On the Standard protocol, male manifest zQ175dn performed equally well to WT for time on rotarod after weight normalization (Fig. 3B) in contrast to performing better than WT before weight normalization (Additional file 3: Figure S3C). Female manifest zQ175dn performed significantly worse than WT littermates for normalized time on rotarod on the Standard protocol (Fig. 3C). As expected, male and female premanifest zQ175dn mice did not show any deficit on rotarod and actually performed better than WT littermates (Standard protocol; Fig. 3D, E), and there was no difference in weight between genotypes at this age (p = 0.6456; unpaired t-test; males and females pooled).
Fig. 3
Rotarod performance and effects of weight in zQ175dn mice and wild-type (WT) littermates. A Left panel: Weight (g) on day 1 of rotarod training in male and female manifest zQ175dn mice compared to WT littermates (overall genotype effect, p < 0.0001****; genotype effect for male manifest zQ175dn vs. WT littermates (multiple comparisons), p < 0.0001****; genotype effect for female manifest zQ175dn vs. WT littermates, p = 0.0043**; two-way analysis of variance [ANOVA] with multiple comparisons). Right panels: Correlation between weight (g) and average time on rotarod on day 1 of training for manifest zQ175dn mice and WT littermates (Pearson’s r correlation test). B Time on rotarod normalized by weight (see methods for details) for male manifest zQ175dn mice and WT littermates that performed the Standard Protocol. C Time on rotarod normalized by weight for female manifest Q176/B6 mice and WT littermates. D Time on rotarod for male premanifest zQ175dn mice and WT littermates. E Time on rotarod for female premanifest zQ175dn mice and WT littermates. F Average number of paw slips per minute for male manifest zQ175dn mice and WT littermates. G Average number of paw slips per minute for female manifest zQ175dn mice and WT littermates. Two-way ANOVA used for all statistical analysis unless otherwise noted. Asterisks (*) denote significance level. ns = not significant. M = Male. F = Female
Male, but not female, manifest zQ175dn mice show altered paw kinematics on the rotarodPreviously, our group developed a method to examine paw kinematics on the rotarod, by tracking the vertical position of the paws relative to the rotarod [13]. Here, we found increased paw slip (when one or both paws dip below the bottom of the rotarod, not normalized by weight) frequency in the male manifest zQ175dn mice, particularly on day 4 of training (Fig. 3F). In contrast, female manifest zQ175dn mice did not show any difference in paw slip frequency (Fig. 3G), and both male and female premanifest mice showed similar paw slip frequencies to WT littermates (male premanifest zQ175dn: 8.08 ± 0.50 slips/minute, WT littermates: 7.54 ± 0.26 slips/minute, genotype effect, p = 0.5228; female premanifest zQ175dn: 2.95 ± 0.73 slips/minute, WT littermates: 2.90 ± 0.49 slips/minute, p = 0.9479; 2-way ANOVA).
Water T-maze performance is impaired in zQ175dn mice and affected by animal weightzQ175dn animals performed a water T-maze task (Fig. 4A) in which they located a hidden platform in one of two arms, to assess response learning and reversal learning. Interestingly, animal weight and time to platform on day 1 of the acquisition phase was positively correlated in the 9 – 11-month old male and female WT mice (males: r = 0.6239, p = 0.0129*; females: r = 0.8853, p = 0.0190*; Pearson’s r correlation), whereas this correlation was not present in the manifest zQ175dn mice (males: r = 0.08865, p = 0.8206; females: r = 0.3864, p = 0.1548; Pearson’s r correlation). Due to the lower weight of manifest zQ175dn mice compared to WT littermates (Fig. 3A), these mice may have an advantage that occludes behavioural deficits caused by the HD mutation. Thus, we normalized for weight in our analysis of time to platform (see methods; non-normalized time to platform data and data separated by sex shown in Additional file 4: Figure S4).
Fig. 4
Water T-maze performance and behavioural feature analysis in zQ175dn mice and WT littermates. A Schematic of the water T-maze with red box outlining the area defined as the “decision point” for the behavioural feature analysis. B Left panel: Time to platform normalized by weight (see methods for details) for manifest zQ175dn mice and WT littermates (males and females) during the acquisition phase. Right panel: Number of arm entries for manifest zQ175dn mice and WT littermates (males and females) during the acquisition phase. C Left panel: Time to platform normalized by weight for manifest zQ175dn mice and WT littermates during the reversal phase. Right panel: Number of arm entries for manifest zQ175dn mice and WT littermates during the reversal phase. D Time to platform during the reversal phase for male (left panel) and female (right panel) premanifest zQ175dn mice compared to WT littermates. E Correlation between percentage of time at decision point and time to platform (not normalized) for day 3 of acquisition (Pearson’s r correlation test) in male manifest zQ175dn mice and WT littermates. F Correlation between number of accelerations per trial and time to platform (not normalized) for day 3 of acquisition (Pearson’s r correlation test) in male manifest zQ175dn mice and WT littermates. Note: due to a flash drive error, 2 female manifest zQ175dn and 5 WT littermates were excluded from analysis for days 1 and 2 of acquisition for water T-maze experiments (therefore for these two days n = 22 for female manifest zQ175dn and n = 21 for WT littermates). See methods for details. Asterisks (*) denote significance level. Individual values for groups with n < 6 are provided in Additional file 7: Individual values. ns = not significant. M = Male. F = Female
In the acquisition phase, manifest zQ175dn mice had an increased normalized (weight-corrected) time to platform and a higher number of arm entries (Fig. 4B; Additional file 4: Figure S4A, B). Immediately following day 3 of the acquisition phase, all mice performed a probe trial to assess whether they predominantly used an egocentric (striatum-dependent) or allocentric (hippocampal-dependent) strategy [16] to locate the platform. We did not find a significant difference between WT littermates and manifest zQ175dn groups, but all groups showed a greater proportion of mice using an egocentric approach rather than allocentric (Additional file 5: Figure S5A). Of the mice that reached the reversal phase (male WT: 12/15; male manifest zQ175dn: 5/9; female WT: 10/11; female manifest zQ175dn: 14/17), we found a significantly increased normalized time to platform for zQ175dn males and females combined (Fig. 4C) compared to WT mice, and a trend for this in males and females separated (Additional file 4: Figure S4C), but no difference in number of arm entries (Fig. 4C; Additional file 4: Figure S4D).
Premanifest zQ175dn males did not show any difference in time to platform in the acquisition phase, while premanifest females were faster than WT littermates (Additional file 6: Figure S6A). We found a similar pattern for the number of arm entries, with premanifest males showing no genotype effect and premanifest females performing better than WT littermates (Additional file 6: Figure S6B). 60% or more of mice in all groups used an egocentric strategy in the probe trial, as opposed to an allocentric strategy (Additional file 5: Figure S5B). For the mice that reached the reversal phase (male WT: 8/10; male premanifest zQ175dn: 11/12; female WT: 5/9; female premanifest zQ175dn: 8/9), male premanifest zQ175dn mice had a slower time to platform compared to WT littermates but females did not (Fig. 4D), while number of arm entries did not show a genotype difference for either sex (Additional file 6: Figure S6C).
Time to platform on Water T-Maze is impacted by various behavioural featuresWe were interested in the finding that male manifest zQ175dn exhibited significantly increased normalized time to platform in the acquisition phase (and a trend for this in the reversal phase), yet these mice did not show any difference in number of arm entries (Additional file 4: Figure S4B). After testing, all mice underwent a swimming speed assessment. We did not find any difference in the swimming speed results between male and female manifest zQ175dn and their WT littermates (male manifest zQ175dn, 19.5 ± 1.25 cm/s, WT littermates, 19.6 ± 1.25 cm/s, p = 0.9469; female manifest zQ175dn, 18.5 ± 0.97 cm/s, WT littermates, 18.3 ± 1.69 cm/s, p = 0.9081; unpaired t-tests). Therefore, the difference in time to platform did not seem to be due to increased number of arm entries or slower swimming speed. We hypothesized that variability in the time spent at the “decision point”, the area between the arms where mice must turn right or left, could be contributing to differences in time to platform.
We used the software PyMouseTracks [15] to track mouse position in the maze and determine the amount of time spent at the decision point (Fig. 4A). Using this approach, we found a difference in male manifest zQ175dn mice, which spent a larger portion of the trials at the decision point on day 3 of the acquisition phase (portion per trial (%) at decision point: male manifest zQ175dn: 19.4 ± 2.5, WT littermates: 26.1 ± 4.17, interaction effect, p = 0.0259; 2-way ANOVA). For the reversal phase, we did not find any significant differences in time at the decision point (across 3-day reversal phase: interaction effect, p = 0.6891; 2-way ANOVA). However, if spending more time at the decision point is a contributing factor to longer time to reach the platform, we should find a positive correlation between these two measures. Indeed, we found that normalized time to reach the platform in the acquisition phase was positively correlated with the percentage of time at the decision point in both genotypes (male manifest zQ175dn, r = 0.2849, p = 0.0104*; WT littermates, r = 0.3672, p < 0.0001****) and this correlation was stronger when examining day 3 alone (Fig. 4E).
Another factor that could impact time to platform is the number of times an animal stops and restarts in the maze. We analyzed this feature for male manifest zQ175dn mice with PyMouseTracks by counting the number of accelerations per trial in the acquisition phase. We did not find any significant genotype differences in the number of accelerations per trial (male manifest zQ175dn: 6.9 ± 0.50, WT littermates: 7.3 ± 1.5; interaction effect, p = 0.2019; 2-way ANOVA). However, we did find a correlation between normalized time to platform and number of accelerations in male WT mice (r = 0.3372; p < 0.0001****) and the correlation between these two measures was significantly stronger in male manifest zQ175dn mice compared to WT (r = 0.5632; p < 0.0001****; Fisher’s z test, p = 0.022*). On day 3 alone, both genotypes had a strong correlation between number of accelerations and normalized time to platform (Fig. 4F). Altogether, these data show that percentage of time at the decision point and number of accelerations are two behavioural features contributing to time to reach the platform in this task.
Manifest zQ175dn mice have a significant learning deficit on a lever-pulling taskConsidering the diverse behavioural patterns discerned across our tests, we assessed animals' motor learning and performance on a task requiring more fine motor control. We employed a home-cage strategy—a non-invasive, hands-off approach that reduces experimenter bias. The Pipaw task, a motor control and motor learning assay, was previously employed to evaluate fine motor skills in the Q175FDN strain at 10 months of age (Woodard et al., 2021). Here, we used the same task to assess the zQ175dn mouse model. Group-housed mice had 24/7 access to the task chamber. A trial was deemed successful when a mouse pulled a lever within a designated goal range (Fig. 5A; see methods section). On average, mice attempted 200 to 400 trials daily.
Fig. 5
Lever pulling assay and slice electrophysiology in male manifest (10 – 12 month old) zQ175dn mice and wild-type (WT) littermates. A Schematic of the testing system of the PiPaw home-cage-based automated testing system used for the lever pulling assay. During the final testing phase, mice must pull the lever into the goal range of 12°—21° to earn a reward. B Success rate per training day in zQ175dn mice compared to WT littermates (mixed effects analysis). C Average number of trials performed per day of training (mixed effects analysis). D Total number of trials performed over the entire training period (p = 0.1920; unpaired t-test). E Spontaneous event frequency for striatal slices harvested from male manifest zQ175dn mice (n = 3 animals) and WT littermates (10 – 12 months of age; n = 3) previously engaged in the PiPaw task for 3–4 weeks (two-way analysis of variance [ANOVA] with multiple comparisons). Asterisks (*) denote significance level. Individual values for groups with n < 6 are provided in Additional file 7: Individual values. ns = not significant. M = Male
Our analysis, focused on the male manifest zQ175dn group, showed a significant learning deficit compared to WT littermates, as illustrated by a slower rise in success rate over days (Fig. 5B), even though they consistently performed an equal or higher number of daily trials (Fig. 5C). This delayed learning trajectory in the zQ175dn mice eventually reached a performance threshold comparable to that of WT littermates. The average daily trials for both WT and zQ175dn remained consistent over time, suggesting stable motivation to engage with the task (Fig. 5C). Notably, despite the zQ175dn mice engaging in more trials over time, when evaluating the entire learning course, no significant difference was found in the total number of trials between groups (Fig. 5D).
Reduced spontaneous excitatory activity in acute brain slices from male manifest zQ175dn miceBuilding on our behavioural observations, we sought to examine potential neural correlates in striatal activity. Specifically, we aimed to ascertain whether our findings would align with previously reported changes in the spontaneous excitatory postsynaptic currents (sEPSCs) mediated by cortical glutamate release, recorded from spiny projection neurons (SPNs) in acute corticostriatal brain slices from naive HD mice of a similar age [10, 17,18,19]. This exploration was important for ensuring consistency in our understanding of the disease's progression at the cellular level.
To probe the spontaneous activity of SPNs in the dorsolateral striatum, whole-cell voltage-clamping was employed on sagittal brain slices from male manifest zQ175dn and WT mice that had engaged in the Pipaw lever-pulling task. SPNs were held at -70 mV and frequency and amplitude of sEPSCs were recorded. As expected, zQ175dn mice exhibited lower sEPSC frequency than WT littermates (Fig. 5E). No difference was observed in the amplitude of sEPSCs (WT: 14.81 ± 0.48 pA, n = 9(3); zQ175dn: 13.86 ± 0.43 pA, n = 17(3)). Our findings corroborate previous observations [10, 19] indicative of consistent alterations in striatal inputs in the progression of HD.
留言 (0)