Gene therapy based on recombinant adeno-associated virus (rAAV) has revolutionized the clinical management of various genetic diseases, with approved AAV drugs either by the EMA, the FDA or both: Glybera [1] (lipoprotein lipase deficiency), Luxturna [2] (for Leber congenital amaurosis), Zolgensma [3] (for spinal muscular atrophy), Roctavian [4] (for Hemophilia B), Upstaza [5] (for severe aromatic L-amino acid decarboxylase deficiency) and Hemgenix [6] (for Hemophilia B). Over a hundred clinical trials using AAV drugs have been conducted to correct defective human genes, establishing AAV as the leading in vivo gene delivery system due to its low immunogenicity, customizable design, and safety profile. Despite its rapid advancements, certain challenges remain, including limited tissue tropism, suboptimal transduction efficacy in specific tissues, pre-existing neutralizing antibodies (NAbs), and the requirement for high vector dosages [7, 8]. Specifically, optimizing target-specific vectors, enhancing transduction efficiency, and ensuring immune evasion are key areas that require further advancement. Overcoming these hurdles will be instrumental in expanding the impact and effectiveness of AAV-based therapies in diverse clinical settings.
The target tissues and cell types for transgene expression are primarily determined by AAV capsid, and different wild-type AAV serotypes exhibit distinct tissue tropisms due to variable regions interacting with specific cell surface receptors [9, 10]. Therefore, the AAV capsid must engage with various host organism components to facilitate effective transduction of target cells. The binding of AAVs to cell surface glycans is believed to act as attaching factors that enhance internalization by increasing interactions with transmembrane receptor proteins responsible for entry. The presence of specific types of glycans can influence binding and internalization in a manner specific to each serotype, potentially affecting tropism. Heparan sulfate proteoglycan was identified as an essential component for cell binding and transduction for AAV2, establishing it as the first recognized receptor [11]. Subsequent research revealed specific glycans for other serotypes, such as sialic acid moieties for AAV5, -1, -6, and -4, and galactose for AAV9 [12,13,14,15]. Glycans are accepted as 'primary receptors' implying an early interaction, characterized by low-affinity, indiscriminate binding of these glycans, which lead to sequestration and concentration of AAV at the cell plasma membrane. Distinct glycan affinities could play a pivotal role in determining tissue and host compatibility. However, the specificity provided by the glycans seems relatively limited.
In 2016 AAV receptor (AAVR) was recognized as an essential receptor for the entry of various AAV serotypes [16]. While AAVR was initially the main focus, it's evident that more than 40 host genes influence AAV transduction acting as partners or engaging with AAV at different stages along an AAVR-mediated pathway playing a role in entry, trafficking, and uncoating processes. Recently GPR108 has been described as an AAV entry factor, predominantly found in the Golgi, suggesting its involvement in virus trafficking or escape mechanisms [17, 18]. Moreover, ATP2C1, that encodes a membrane calcium transport pump, is crucial for upholding cellular calcium balance and impacts on AAV transduction revealing that proper cellular calcium levels are necessary for effective intracellular trafficking and structural alterations in the AAV capsid, as it has been observed in other viruses [19]. These factors all adhere to specific locations on the capsid to function as transduction agents. Furthermore, while glycan-binding sites across different serotypes are typically situated similarly, they do not exhibit amino acid alignment in phylogenetic comparisons.
Consequently, AAV tropism remains broad and unspecific, limiting their application in certain tissues of interest. Additionally, pre-existing NAbs in the human population against specific AAV serotypes can impede effective transduction or render patients ineligible for treatment. To overcome these limitations, researchers worldwide have explored modifications to the AAV capsid to expand its potential applications. Various approaches have been investigated, ranging from genetic engineering to chemical modifications, in the development of newly engineered capsids [20]. Modified versions of AAVs encompass a spectrum of approaches, from genetic modifications of the viral cap gene to conjugation of proteins like streptavidin or antibodies to enhance interactions with specific cells. Previous studies have demonstrated that incorporating distinct genetic sequences into the VP2 gene can influence AAV9's targeting to various cells within bone tissue [8] and AAV6's transduction efficiency in diverse cancer cells [8]. For instance, Lee and Ahn found that AAV2 modified with a streptavidin-biotin complex linked to an anti-EpCAM antibody exhibited significant efficacy in targeting EpCAM-positive ovarian cancer cells [21].
Numerous research teams have utilized DNA shuffling and peptide display methods to create innovative AAV capsids with enhanced in vivo transduction capabilities. These novel AAV vectors have shown success in targeting various organs, such as the liver, brain, skeletal muscle, and eye [20]. Additionally, recent efforts have focused on synthetically modifying AAV capsids to improve properties like immune evasion and cell-specific receptor targeting. The presence of glycan binding sites of different serotypes in similar general locations, despite they do not share amino acids that align phylogenetically gave hope for retargeting gene therapy vectors. Consequently, modifications in heparin-binding and the integration of an AAV9-like galactose binding site into AAV2 have resulted in alterations of transduction characteristics, ultimately leading to improved transduction efficiency [22].
Ultimately, chemical modifications of the AAV capsid have unveiled novel pathways for enhancing the properties of recombinant AAVs. Introducing GalNac or Mannose to exposed tyrosines has notably enhanced transgene expression in the liver and retina [23]. Moreover, the impact can be serotype-dependent; for instance, binding N-ethyl Maleimide to the AAV9 capsid has augmented its affinity for murine bone marrow while diminishing transduction in liver tissue, unlike AAV2 and AAV8 which did not exhibit this effect [24]. However it is challenging to generate modifications highly specific and selective, targeting only the desired regions of the capsid.
Emerging developments in synthetic biology have opened doors to the incorporation of a reactive unnatural amino acid onto the surface of the AAV capsid, allowing for precise click chemistry conjugation with molecules at designated sites. For this purpose, sets of distinct prokaryotic tRNA/tRNA synthase pairs have been designed and introduced into mammalian cells, enabling the integration of unnatural amino acids into newly forming polypeptides [25]. This approach has been applied in AAV engineering, introducing an azido group onto the AAV capsid surface to facilitate click chemistry conjugation with molecules such as oligonucleotides [26], aptamers [27], and various chemicals like PEG and folic acid [27, 28]. However, modifications using azido-lysine at different positions across different AAV serotypes (AAV2, AAV8, AAV-DJ/8 and AAVLK03) have shown restricted transduction efficiency and lower titers compared to their respective unmodified wild-type AAV counterparts. Remarkably, these hybrid AAV constructs have exhibited novel attributes, such as the ability to be tailored for precise targeting of particular cancer cells and the capacity to interfere with immune responses by mitigating the generation of neutralizing antibodies and enhancing resistance to neutralization by Nabs [26,27,28].
Nonetheless, the modification of AAV capsid properties presents challenges that demand a thorough understanding of the essential virus components involved in recombinant virus production and entry interactions. Further investigation is necessary to comprehensively elucidate the precise chemistry of AAV capsids and their interactions with cells, thereby laying the foundation for enhanced and targeted gene therapies.
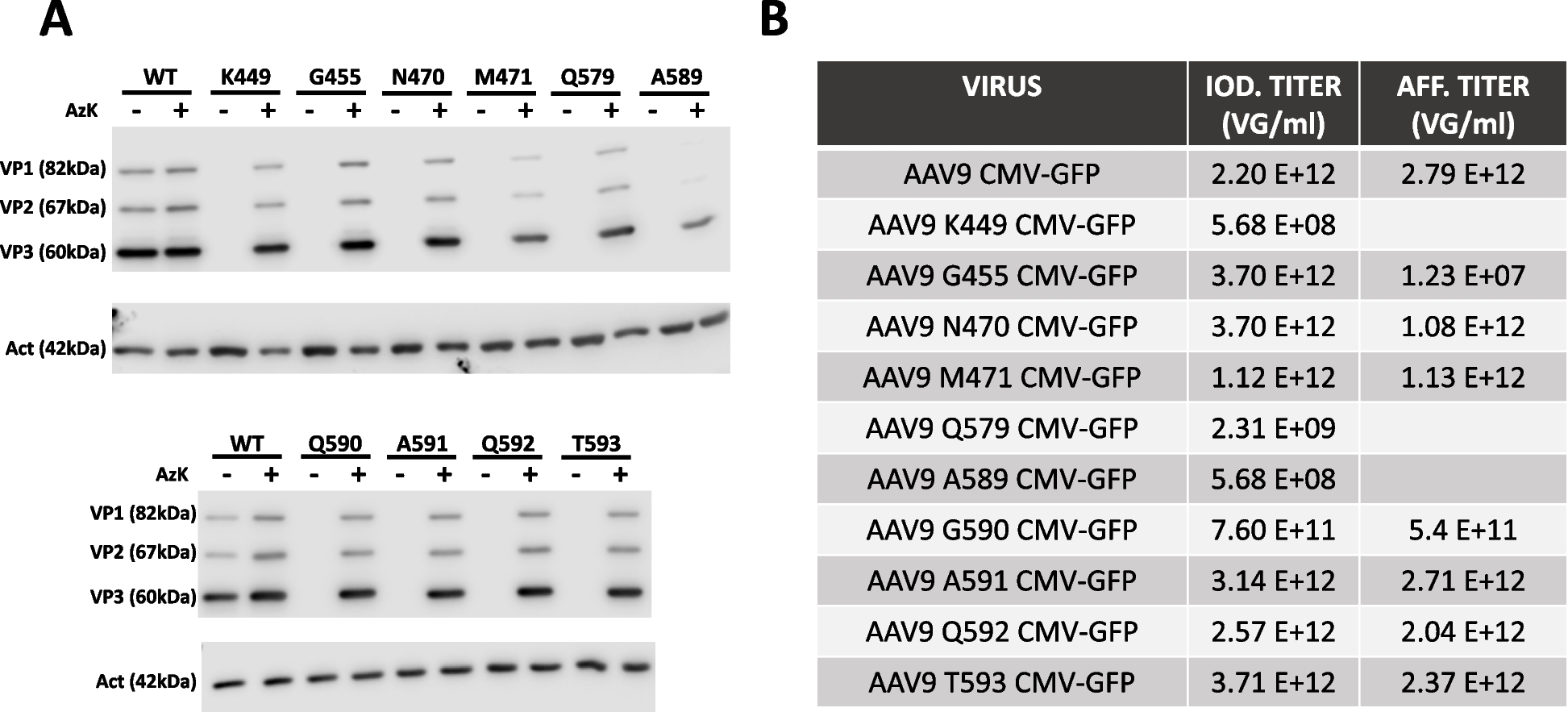
We have examined the potential for several specific residues within the AAV9 capsid to be replaced with azido-lysine, ensuring that it doesn't interfere with AAV production and maintains the ability to attach a Cy5.5 molecule to these altered residues. This assessment involved evaluating the infectivity of these modified versions of AAV9. From our investigation, we have identified five residues that can successfully be substituted with azido-lysine (AzK), resulting in titers comparable to the wild-type AAV9. However, these modified versions exhibit varying transduction capacities. Additionally, we have noticed that the attachment of the Cy5.5 molecule to the AAV9 capsid leads to differing outcomes, depending on the specific residue of attachment and the proportion of modified residues within the capsid.

留言 (0)