
記住我
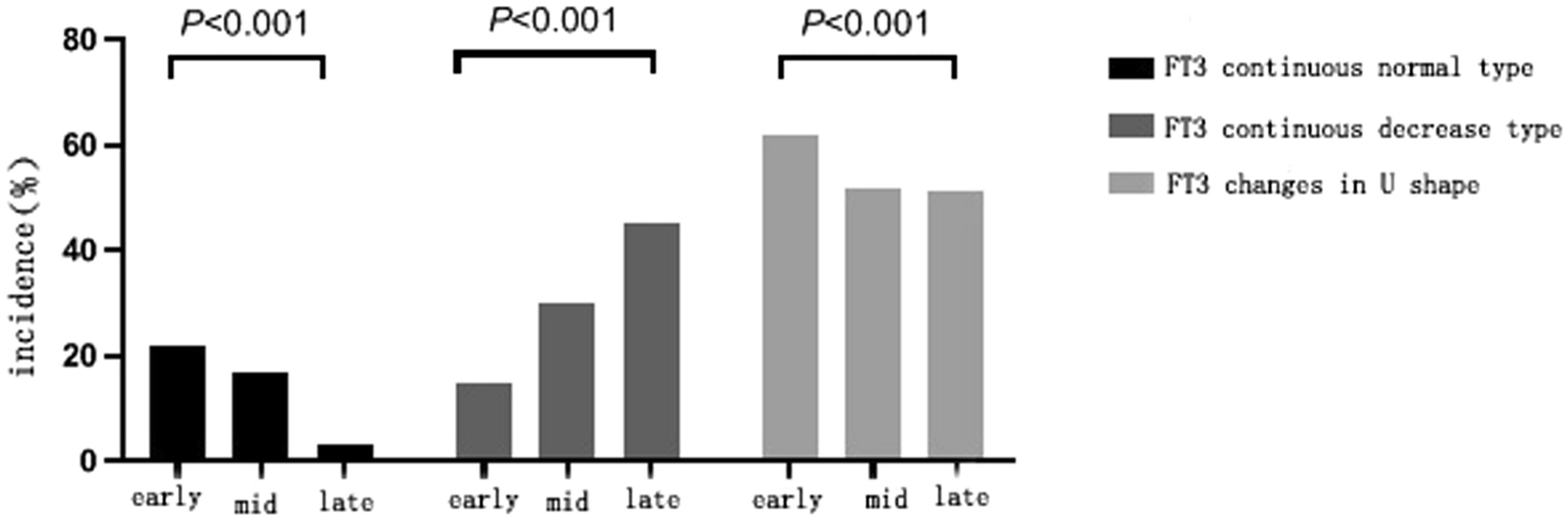

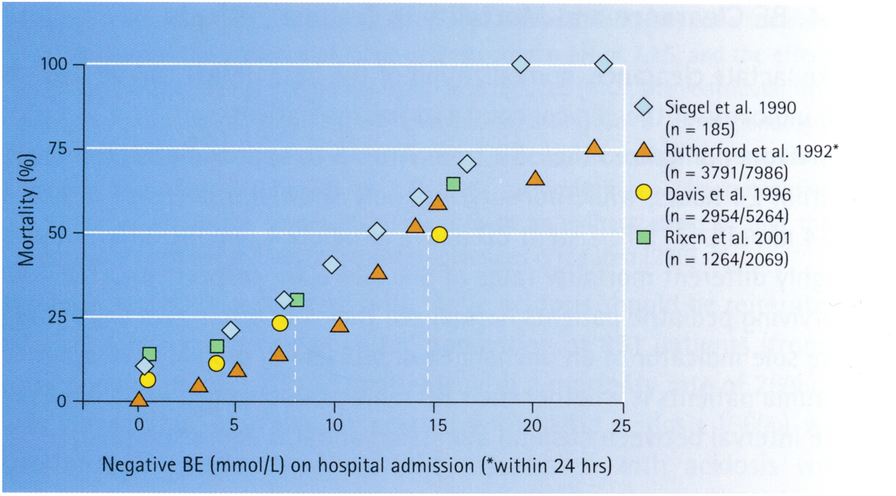



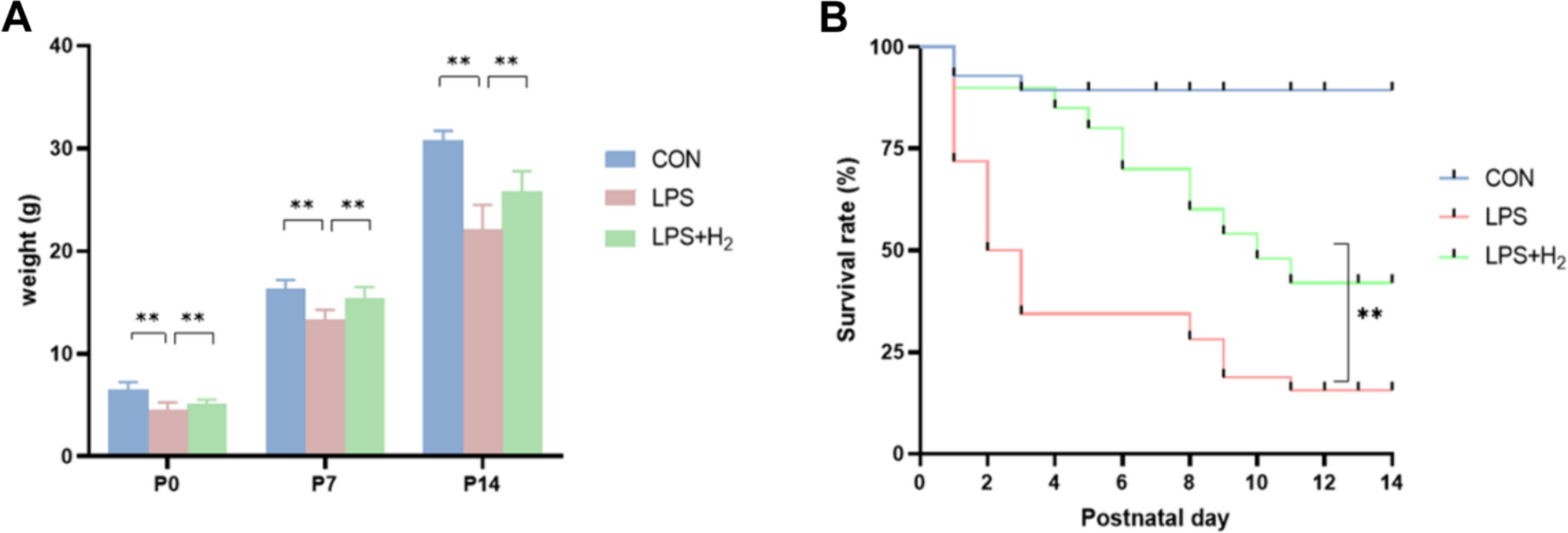

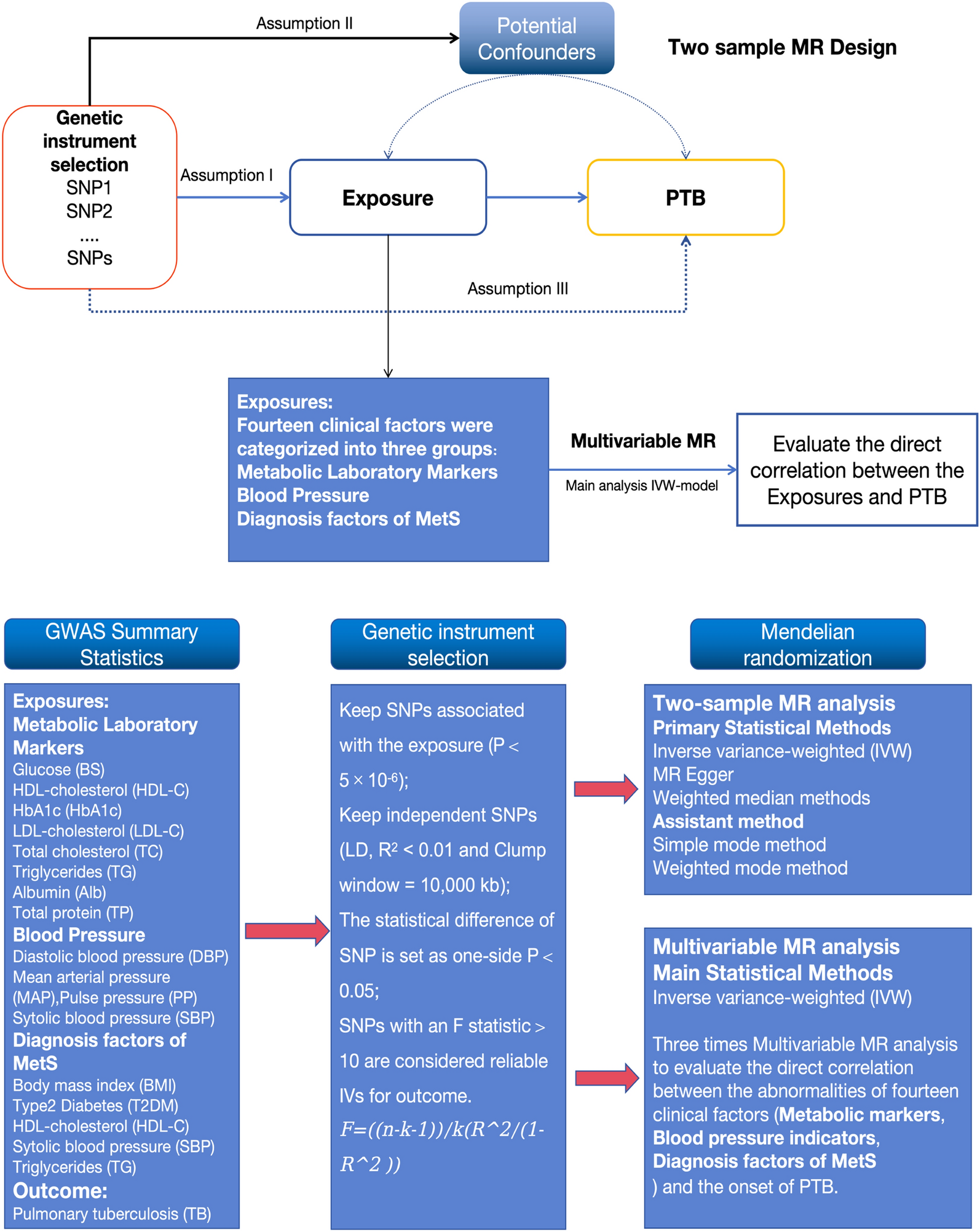

Among the 13 datasets merged in this study, six datasets (GPL96, GPL570, GPL6102, GPL14951, GSE19915_GPL3883, and TCGA_BLCA_mRNA) demonstrated that KCNK1 mRNA was significantly elevated in BC (p < 0.05; Fig. 2A). Pooling the datasets to calculate the composite SMD, KCNK1 mRNA was significantly increased in 1264 BC samples compared to 222 control samples (SMD = 0.58, 95% CI [0.05; 1.11]; Fig. 2B). In Fig. 2C, Egger’s test indicated that there was no publication bias in the results of SMD (p = 0.956). Considering that KCNK1 mRNA expression was significantly different in cancer and control samples, we plotted the ROC of each dataset to assess the distinguishing ability of KCNK1 mRNA. The results showed that KCNK1 mRNA expression had an extremely strong ability in distinguishing cancer and control samples (Additional file 2: Figure S2A), which was further validated by the results of the combined sROC (AUC = 0.82 [0.78–0.85]; Fig. 2D).
Fig. 2
Comparison of differences in KCNK1 mRNA expression levels in control and bladder cancer groups. A KCNK1 mRNA was differentially expressed in each dataset. B Forest plot of standardised mean difference of KCNK1 mRNA in bladder cancer tissues. C Egger’s test. D Summary receiver operator characteristic
KCNK1 protein levels in BC were detected by in-house IHCFurther, we performed IHC staining of in-house samples (n = 245) to verify KCNK1 expression in BC tissues at the protein level. According to Fig. 3A–E, KCNK1 protein stained weakly positive in control tissue samples under the microscope. Compared with the control samples, KCNK1 protein stained strongly positive in BC tissue samples by IHC (Fig. 3F–J), and the protein level was significantly elevated (p < 0.0001; Fig. 3K). The ROC curves indicated that at the protein level, highly expressed KCNK1 also possessed an excellent ability to distinguish between BC and control samples (AUC = 0.99 [1.00–0.98]; Fig. 3L).
Fig. 3
KCNK1 protein levels were assessed by immunohistochemistry (IHC) assay based on in-house bladder cancer samples. A–E IHC staining of control samples. F–J IHC staining of cancer samples. K Protein levels of KCNK1 in bladder cancer and control samples. L Receiver operator characteristic curves of in-house IHC. ns/NSp > 0.05, ∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001, ∗∗∗∗p < 0.0001
scRNA-seq revealed KCNK1 expression and cellular communicationImmediately following this, we further analysed the distribution of KCNK1 expression in cells of BC tissues and intercellular communication using scRNA-seq. This study filtered to obtain 9255 cells annotated as 8 common cell types (Fig. 4A). KCNK1 was abundantly expressed in BC epithelial cells (Fig. 4B, C), and there was a trend of elevated KCNK1 expression with cell differentiation (Fig. 4D, E). AUCell analysis showed that KCNK1 was mainly active in epithelial cells (Fig. 4F).
Fig. 4
Comprehensive analysis of single-cell RNA sequencing exploring KCNK1 expression. A Included cells were annotated into 8 cell types. B UMAP plot of KCNK1 expression in cells. C Violin plots of KCNK1 expression in different cells. D The cellular differentiation trajectory in pseudo-temporal analysis. E Expression levels of KCNK1 at cell differentiation. F Mapping of AUC values of each cell onto UMAP plots. G Number of cell interactions. H Cell interaction weights/strengths
Considering the large number and weight/intensity of different types of cellular interactions in BC tissues (Fig. 4G, H), we further analysed the specific mechanisms of cellular communication. First, among all ligand-receptor (L-R) pair interactions, the MDK-associated L-R pair was very active in BC epithelial cells (Fig. 5A). Since MDK was active in the MK signalling pathway, we launched a specific analysis of the MK signalling pathway. Figure 5B, C showed that the MK signalling pathway was mainly involved in cellular communication between epithelial cells and iPS cells, Smooth muscle cells and CD8 + T cells, and genes involved in the MK signalling pathway were also mainly expressed in the above cells. MK signalling pathway contributed the greatest intensity among all afferent and efferent signals (Fig. 5D, E). Among them, BC epithelial cells were the main sender and receiver, and CD8 + T cells were the main mediator (Fig. 5F). In addition, BC epithelial cells were identified from the global communication pattern to be mainly involved in Pattern 2. The major signalling to contributors included MK, ANNEXIN, GALECTIN, CCL, BAFF and IFN-II (Additional file 2: Figure S3).
Fig. 5
Cell communication analysis based on the ‘Cellchat’ package. A Ligand-receptor pair interactions in cellular communication. B Hierarchical structure of cell clusters involved in the MK signalling pathway. C Distribution of signalling genes involved in the MK signalling pathway. D Afferent signalling contributions of different cell clusters. E Different cell cluster efferent signalling contributions. F Signalling roles of different cell clusters in the MK signalling pathway
Potential epigenetic regulatory mechanisms of KCNK1 overexpressionIn the present study, we explored the potential regulatory mechanism of KCNK1 in depth to reveal the cause of elevated KCNK1 expression. We used Cistrome Data Browser to predict KCNK1 upstream TFs (Fig. 6A) and obtained two TFs (GRHL2 and FOXA1) after a series of screening. The ChIP-seq of these two TFs possessed peaks at 2 kb before and after the KCNK1 upstream TSS, suggesting that GRHL2 and FOXA1 might act on the KCNK1 TSS and regulate the upregulation of KCNK1 expression (Fig. 6B, C). The motif plots of GRHL2 and FOXA1 were shown in Fig. 6D, E. ATAC-seq suggested that the KCNK1 TSS region had an open chromatin structure and was in an active transcriptional state (Fig. 6B, C). Further, we collected histone modification ChIP-seq of labelled silencers (H3K27me3), promoters (H3K4me3) and enhancers (H3K4me1 and H3K27ac). The results suggested the existence of potentially active (promoters, enhancers) and repressive (silencers) genomic regulatory elements in the KCNK1 upstream TSS and the existence of a complex transcriptional regulatory network in KCNK1 (light red shaded areas in Fig. 6B, C). In addition, there were loops between the promoter of the KCNK1 TSS and the downstream super enhancer (SE) and the downstream typical enhancer (TE), suggesting that the promoter might be spatially engaged with the SE and TE to promote KCNK1 expression (light orange shaded region in Fig. 6B, C). GRHL2 and FOXA1 might also be spatially involved in the above transcriptional regulation (light blue shaded area in Fig. 6B, C). Among them, as a ChIP marker, RAD21 might help promoter-enhancer (P-E) interactions by regulating the spatial structure of chromatin to promote active transcription of genes.
Fig. 6
Epigenetic regulatory mechanisms of KCNK1 expression. A Cistrome Data Browser-based transcription factor screening process. B Multi-omics exploration of potential regulatory mechanisms of KCNK1 (before amplification). C Multi-omics exploration of potential regulatory mechanisms of KCNK1 (after zoom-in). D Motif map of transcription factor GRHL2. E Motif map of the transcription factor FOXA1
Potential pathway enrichment analysisIn this study, there were 1154 HECEGs that could be screened for the next step of analysis. KEGG analysis showed that KCNK1-associated HECEGs were mainly enriched in signalling pathways such as p53 signaling pathway, Amino sugar and nucleotide sugar metabolism, mRNA surveillance pathway and Cell cycle (Fig. 7A). Reactome analysis suggested that HECEGs were mainly involved in APC/C-mediated degradation of cell cycle proteins, Cell Cycle Checkpoints, Regulation of mitotic cell cycle, M Phase and APC/C:Cdc20 mediated degradation of mitotic proteins were significantly enriched (Fig. 7B). GO analysis similarly showed that KCNK1-associated HECEGs were mainly involved in cell cycle-related signalling pathways (Additional file 2: Figure S4).
Fig. 7
Pathway enrichment analysis of highly expressed co-expressed genes of KCNK1. A KEGG enrichment analysis. B Reactome enrichment analysis
Preliminary exploration of the molecular function of KCNK1 in BCIn addition, we tried to clarify the molecular mechanisms and biological pathways of KCNK1. Gene set enrichment analysis (GSEA) showed that KCNK1 was involved in multiple BC cell metabolism-related signalling pathways, especially glucose metabolism (Fig. 8A). Meanwhile, elevated KCNK1 expression could activate the activity of metabolic signalling pathways. Comparing with other cells, single-cell AUCell analysis showed that there were multiple active metabolic signalling pathways in BC epithelial cells, including fructose and mannose metabolism, Drug metabolism—other enzymes (Fig. 8B–I). The molecular mechanism of KCNK1 was further explored from protein interactions using PPI. As a hub gene of the interaction network, KCNK1 was involved in the formation of ion transmembrane transport proteins, especially potassium ions (Additional file 2: Figure S5).
Fig. 8
Preliminary validation of the molecular function of KCNK1. A Gene set enrichment analysis of KCNK1 in bladder cancer. B–I UMAP plots of single-cell metabolic activity scores
Correlation of KCNK1 expression with BC TMETME has an essential role in immunosurveillance, immune escape, immunosuppression and immunotherapy [31]. Figure 9A demonstrated the composition of 22 immune-infiltrating cells in the high KCNK1 expression and low KCNK1 expression groups in BC TME. Comparing with the low KCNK1 expression group, the high KCNK1 expression group had higher levels of infiltration of Dendritic cells activated (p < 0.0001), Dendritic cells resting (p < 0.0001) and T cells follicular helper (p < 0.0001), and lower levels of infiltration of B cells naive (p < 0.0001), Macrophages M2 (p < 0.01) and T cells regulatory (Tregs) (p < 0.0001) (Fig. 9B). In addition to the two immune cells (B cell and Neutrophil), KCNK1 expression was negatively correlated with cellular levels of T cell CD4 (r = − 0.19; p = 1.4e-4), T cell CD8 (r = − 0.16; p = 1.1e-3), DC (r = − 0.10; p = 0.05), and Macrophage (r = − 0.34; p = 1.8e-12) (Fig. 9C). Further, TME was assessed as a whole, and we found that KCNK1 expression was significantly negatively correlated with TME stromal score (r = − 0.22; p = 7.3e-6), immune score (r = − 0.17; p = 5.3e-4) and ESTIMATE score (r = − 0.21; p = 2.1e-5), and positively correlated with tumour purity (r = 0.22; p = 3.9e-6) (Fig. 9D). TME scores were significantly different in the high and low KCNK1 expression groups (p < 0.05; Fig. 9E).
Fig. 9
Significance of KCNK1 expression in the tumour microenvironment (TME). A TME composition of the high KCNK1 expression group and the low KCNK1 expression group. B KCNK1 expression was significantly correlated with the level of multiple immune cell infiltration. C Correlation of KCNK1 expression with multiple immune cells. D Correlation of KCNK1 expression with TME scores. E Significant differences in TME scores between the high KCNK1 expression group and the low KCNK1 expression group. ns/NSp > 0.05, ∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001, ∗∗∗∗p < 0.0001
Clinical value of KCNK1 expression in BC patientsWe also explored the clinical significance of KCNK1 overexpression in BC. Based on the clinicopathological characterisation information of the in-house samples, we found that KCNK1 expression was significantly associated with age, pathology grade and pathologic_M in BC patients (Table 2). Among them, KCNK1 expression in BC was significantly different in low-grade (Pathology Grade I) versus high-grade (Pathology Grade II and III&IV) (p < 0.05; Additional file 2: Figure S2B). Immune-targeted therapy suggested that patients in the high KCNK1 expression group had better outcomes when treated with the combination of anti-CTLA-4 and anti-PD-1 (p = 0.019; Fig. 10A). Meanwhile, the high KCNK1 expression group had lower TIDE scores, suggesting a low tumour immune escape potential and a better outcome of immunotherapy for BC patients (p = 5.7e-7; Fig. 10B). Further predicting potential therapeutic drugs for KCNK1, we found that the drug half maximal inhibitory concentration (IC50) of docetaxel, paclitaxel, and vinblastine was low in the high KCNK1 expression group (p < 0.05; Fig. 10C). Molecular docking showed that KCNK1 protein had a high molecular affinity for docetaxel (affinity = − 7.6 kcal/mol), paclitaxel (affinity = − 7.0 kcal/mol) and vinblastine (affinity = − 7.3 kcal/mol) and possessed better drug efficacy (Fig. 10D).
Table 2 Study of the relationship between KCNK1 mRNA expression and clinical characteristics of KCNK1 patients from in-house data Fig. 10
Clinical treatment and molecular docking of KCNK1 expression. A Immune-targeted therapy scores in the high KCNK1 expression group and low KCNK1 expression group. B Tumour immune escape scores in the high KCNK1 expression group and low KCNK1 expression group. C Assessment of the pharmacological therapeutic potential of KCNK1 expression. D Molecular docking to assess the affinity of KCNK1 protein to drugs
留言 (0)