
記住我



MEFs91 (received from the Eriksson lab, Åbo Akademi University) were cultured in DMEM (Sigma-Aldrich, D5671), supplemented with 10% FCS (Sigma-Aldrich, F7524), 2 mM l-glutamine (Sigma-Aldrich, G7513) and 100 μg ml–1 penicillin–streptomycin (Sigma-Aldrich, P0781), at 37 °C and 5% CO2 in a humidified incubator.
3D-SIM imagingSub-confluent cultures of MEFs growing on no. 1.5 glass coverslips were fixed with 4% paraformaldehyde for 10 min at room temperature (RT). The fixed cells were permeabilized with 0.1% Triton X-100 for 10 min at RT and then were incubated with chicken anti-vimentin (1:200, 919101, Biolegend) for 30 min in phosphate-buffered saline (PBS) containing 5% normal goat serum. This was followed by staining with Alexa-Fluor-488-conjugated goat anti-chicken antibody (1:400, A-11039, Invitrogen) and DAPI in PBS for 30 min. The stained cells were mounted with ProLong Glass Antifade Mountant (Life Technologies).
Moreover, MEFs were seeded on no. 1.5 glass coverslips; the next day, they were washed with PBS and 2 mM MgCl2 for 5 s, followed by incubation with PBS containing 0.1% Triton X-100, 10 mM MgCl2, 0.6 M KCl and protease inhibitors for 25 s at RT. The extracted cells were rinsed with PBS and 2 mM MgCl2 for 10 s and subsequently incubated with 2.5 units µl–1 benzonase for 30 min at RT. After rinsing with PBS and 2 mM MgCl2, the cells were fixed with 4% paraformaldehyde for 5 min at RT. The fixed cells were then stained with chicken anti-vimentin (1:200, 919101, Biolegend) and rabbit anti-lamin-A/C (to determine the location of the nucleus; 1:100, sc-376248, Santa Cruz Biotechnology) antibodies in PBS containing 5% normal goat serum for 30 min at RT, followed by incubation with goat anti-chicken and anti-rabbit secondary antibodies (1:400, A-11039, A-11011, Invitrogen) for 30 min at RT. After being washed in PBS, stained cells were mounted with Prolong Glass Antifade Mountant (Life Technologies).
3D-SIM imaging was done using an N-SIM Structured Illumination Super-Resolution Microscope system (Nikon) using an oil immersion objective lens (SR Apo TIRF100X, 1.49 numerical aperture (NA), Nikon). For 3D-SIM, 26 optical sections were imaged at 100-nm intervals. The raw SIM images were reconstructed with the N-SIM module of Nikon Elements Advanced Research with the following parameters: illumination contrast, 1.00; high-resolution noise suppression, 0.75; and out-of-focus blur suppression, 0.25. Brightness and contrast were adjusted for image presentation.
Cryo-FIB and cryo-ET in situ analysis of VIFsMEFs were seeded onto glow-discharged, carbon-coated, gold EM grids (Quantifoil holey carbon R 2/1, Au 200) in standard medium overnight. After a single wash in 1× PBS (Fisher Bioreagents, BP399-1), the grids were vitrified in liquid ethane using a manual plunge freezing device.
The grids were coated with 5–10 nm platinum on carbon with a Leica BAF060 system cooled to −160 °C, prior to cryo-FIB milling. Next, the grids were transferred to the focused ion beam scanning electron microscope (FIB-SEM) (Zeiss Auriga 40 Crossbeam), equipped with a Leica cryo-stage. After transfer, the grids were coated with two 5-s flashes of organometallic platinum, using the internal gas-injection system, set at 28 °C. The cells were milled with a focused gallium ion beam at a constant voltage of 30 kV and a current between 10 and 240 pA at a stage angle of 18°. The process was controlled by the Nano Patterning and Visualization Engine (Zeiss) and monitored with the SEM at 5 kV. The resulting FIB milled lamellae were 100–200 nm thick (Extended Data Fig. 2b).
The cryo-FIB milled grids were transferred to a Titan Krios 300 kV cryo-TEM (Thermo Fisher Scientific) equipped with a K2 summit direct electron detector and a Quantum energy filter (Gatan). The tilt series were acquired using SerialEM92 with a 100-µm objective aperture at a magnification of ×64,000, a pixel size of 2.21 Å and −4-µm defocus; a dose-symmetric tilt scheme was used93, from −60° to +60° in 3° increments starting from 0°. Overall, 102 tilt series were acquired, each with a total dose of ~160 e–/A2.
The tomograms were reconstructed and binned four times (pixel size, 8.84 Å) with weighted back-projection (WBP) using IMOD94 or AreTomo95. If platinum remnants were present in the tilt series, IMOD was used for tomogram reconstruction. The platinum particles were used as fiducial markers for tilt-series alignment. AreTomo was used for marker-free tilt series alignment.
Platinum particles were typically highly concentrated at the front edges of the lamellae and served as fiducial markers for precise tilt-series alignment (Extended Data Fig. 1b, blue dashed rectangle). However, in the tomograms, these high-contrast particles introduce a high density of distorting back-projection rays in the tomographic volume above and below the front edge of the lamellae (Extended Data Fig. 1b, red dashed rectangle). These regions were excluded from subsequent data analysis. Therefore, the processed volume of a tomogram was cut out from the original tomogram (Extended Data Fig. 1b, yellow dashed rectangle, and Supplementary Video 1).
Subtomogram averaging of VIF segmentsFirst, to determine the native protofibril stoichiometry of VIFs, we applied a subtomogram averaging approach. From the dataset, we selected seven representative tomograms with precise tilt-series alignment (residual error, ~3 Å) and a Thon ring spectrum featuring up to seven non-astigmatic Thon rings visible at 0° tilt. Subsequently, we trained a crYOLO network to detect the VIFs. We localized 14,273 subtomograms (35 × 35 × 35 nm3) containing VIF segments, which were distributed over 1,114 filaments.
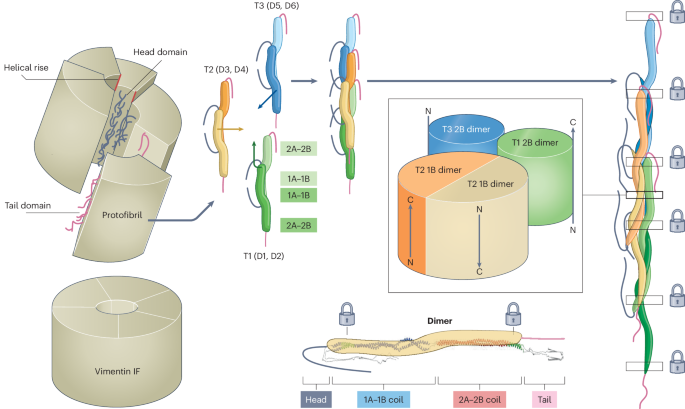
We then applied the Actin Polarity Toolbox (APT) pipeline47,69,70. Here, the subtomograms were projected with a projection thickness of 16 nm. False positive (for example, actin filaments, microtubule walls or vesicle membranes) and low-quality VIF segments were excluded by multiple rounds of unsupervised 2D classification in RELION50,56. This procedure yielded a particle stack of 2,371 good VIF segments. On the basis of the geometrical orientation of the picked filaments in the tomograms, crYOLO calculates initial priors for the in-plane rotation and the tilt angle for each subtomogram (referred to as psi and tilt priors in the RELION convention). The third Euler angle (rot angle) remains a priori unknown. Therefore, we assigned to each VIF segment a random rot prior, and used these priors to reconstruct a rotationally symmetric, tube-like VIF 3D template4. Next, we exported the VIF segments and their priors from RELION to cryoSPARC96 and performed a 3D reconstruction, using the previously generated rotational symmetric structure as initial reference. Without applying any assumptions on the symmetry of the assembly, the average converged to a resolution of ~20 Å and clearly shows the five-protofibril architecture of VIFs (Extended Data Fig. 1c).
Assuming that VIFs are assemblies with helical symmetry, an initial hypothesis about the helical-symmetry parameter of VIFs can be derived from this result, because a five-protofibril architecture would directly translate to a helical twist angle of 360° / 5 = 72°. Second, a characteristic feature of IFs is their ~210-Å axial periodicity1,30,52,53,54. This would directly translate to a helical rise of 210 Å / 5 = 42 Å. We then repeated the previous 3D reconstruction in cryoSPARC, but with application of helical symmetrization, with a helical rise of 42 Å and a helical twist of 72° (Fig. 1d, Extended Data Fig. 1d and Supplementary Fig. 1).
Cross-section analysis of VIFsAn alternative method to determine the native protofibril stoichiometry of VIFs is to directly evaluate their tomographic cross-sections4,49. However, most of the VIFs in the dataset are oriented in the xy plane of the tomograms, with a maximal out-of-plane tilt of ~15°, as measured previously by the tilt-angle distribution of the filaments during crYOLO picking and subtomogram averaging of VIF segments. Therefore, their cross-sections are contained mainly in the xz planes of the tomograms and cannot be directly analyzed, because they are distorted by the missing-wedge artifact68.
To overcome this problem, we used IsoNet71 to compensate for the missing-wedge-induced resolution anisotropy in VIF cross-sections. IsoNet is a convolutional neural network that can be trained to learn missing-wedge deconvolution. We used the seven tomograms that we had already selected as input for IsoNet. The network was trained on the full tomograms containing the edge of the lamellae with a high number of platinum particles, which were previously used as fiducial markers for tilt-series alignment. In this case, they serve as high contrast, point-like features that clearly reproduce the missing-wedge convoluted point spread function in the tomograms. Therefore, they could have an advantageous effect on the convergence of the deconvolution network, but at the very least clearly show reduction of the missing wedge after missing-wedge deconvolution (Extended Data Fig. 2b, blue dashed rectangle). After missing-wedge deconvolution, the tomograms were cut to the same volumes as before (Extended Data Fig. 2b, yellow dashed rectangle, and Supplementary Video 2) to exclude regions with a high concentration of back-projection rays caused by the large number of platinum particles at the edge of the lamellae (Extended Data Fig. 2b, red dashed rectangle). The following parameters were used during the IsoNet workflow: for contrast transfer function (CTF) correction, a signal-to-noise ratio fall off parameter of 0.7 was used. For automatic mask creation, a density percentage of 50% and s.d. percentage of 50% were chosen. Training of the missing-wedge deconvolution neural network was based on randomly extracted 2,000 subtomograms (box size, 643 voxels) per tomogram within the automatically created mask, with the following training parameters: 30 iterations; learning rate, 0.0004; dropout rate, 0.5.
The VIF cross-sections (Extended Data Fig. 2c) were extracted from one of the missing-wedge-corrected tomograms. These cross-sections confirm the five-protofibril architecture of VIFs. However, to extract a meaningful statistical conclusion from the cross-section data, we developed the following procedure. We used the 3D coordinates of the previously established 2,371 good VIF segments and extracted their related subtomograms from the missing-wedge-corrected tomograms. Then, the initial crYOLO priors (in-plane rotation and tilt angle) and random rot angles were used to align the VIF segments. An average of these subvolumes yields a rotational symmetric, tube-like 3D structure of VIFs, similar to that used before, as an initial reference for 3D reconstruction. However, in this case, the subvolumes were extracted from the missing-wedge-corrected tomograms. We next extracted the central cross-section from each of the aligned subvolumes, and applied a 2D classification in RELION on these images without alignment. Because the cross-sections are missing-wedge corrected, they can be classified without using a metric that explicitly models the missing-wedge effect97. Because the alignment was fixed in this 2D classification, the cross-sections were combined into classes that are similar, under the application of a random rot angle. In this manner, we identified a subset of 444 aligned cross-sections. Finally, these cross-sections were extracted from the original WBP subvolumes and averaged. This cross-section average (Extended Data Fig. 2d) also confirms the five-protofibril architecture of VIFs.
By contrast, a cross-section located in the xy plane of a tomogram exhibits isotropic resolution and can be evaluated directly. Therefore, we identified a tomogram that contained a vimentin bundle oriented approximately parallel to the electron beam (Extended Data Fig. 3a,b, left column). For contrast enhancement, the tomogram was binned four times and denoised using Topaz-Denoise98. To ensure that the denoising operation did not create artificial density, we compared the denoised tomogram with its original version (WBP reconstruction in AreTomo), low-pass filtered to 30 Å (Extended Data Fig. 3a,b, right column). In total, we extracted 150 cross-section images (35 × 35 nm2, 8.84-Å thick) that were located in the xy planes of the denoised tomogram. The xy cross-sections also show the five-protofibril architecture of VIFs (Fig. 1e and Extended Data Fig. 3c).
Cryo-ET of detergent-treated MEFsMEFs were grown to ~80% confluency on glow-discharged holey carbon EM grids (R2/1, Au 200 mesh; Quantifoil) prior to preparation for cryo-ET analysis. Grids with a relatively homogeneous distribution of cells were selected using fine tweezers and then were washed in PBS and 2 mM MgCl2 for 5 s. The grids were treated for 20–40 s in pre-permeabilization buffer (PBS containing 0.1% Triton X-100, 10 mM MgCl2, 600 mM KCl and protease inhibitors) and then rinsed in PBS and 2 mM MgCl2 for 10 s. Next, the grids were incubated with benzonase (2.5 units µl–1 in PBS and 2 mM MgCl2; Millipore, Benzonase Nuclease HC, purity >99%) for 30 min at RT. After the grids were washed with PBS and 2 mM MgCl2, a 3-µl drop of 10 nm fiducial gold markers (Aurion) was applied to the grids. For vitrification, the grids were manually blotted for ~3 s from the reverse side and plunge frozen in liquid ethane.
Tilt-series acquisition was conducted using a Titan Krios transmission electron microscope equipped with a K2 Summit direct electron detector and Quantum energy filter. The microscope was operated at 300 keV with a 100-µm objective aperture. In total, 225 tilt series were collected at a nominal magnification of ×42,000, and the slit width of the energy filter was set to 20 eV. Super-resolution videos were recorded within a tilt range from −60° to +60° with 2° increments using SerialEM92. The image stacks were acquired at a frame rate of 5 frames per second, with an electron flux of ~2.5 e–/pixel/s. The tilt series were recorded with a total electron dosage of ~125 e–/Å2 and within a nominal defocus range of −2 μm to −6 μm. The super-resolution image stacks were drift-corrected and binned twice using MotionCorr99, resulting in a pixel size of 3.44 Å for the tilt series. For each projection, the defocus was measured, and the CTF was corrected by phase-flipping. Then, from each tilt series, an overview tomogram that had been binned four times was reconstructed (Supplementary Fig. 2b and Supplementary Video 3). CTF correction and tomogram reconstruction were performed using MATLAB scripts (MathWorks), derived from the TOM toolbox100,101.
Algorithms from EMAN2 (ref. 102) were used to train a convolutional neural network that could segment VIFs in the overview tomograms. The segmentations were manually checked and cleaned from obvious false-positive VIF detections in Chimera103. On the basis of scripts derived from APT47, two sets of segment coordinates were extracted from the segmentations. In the first set (values for the second set are in parentheses), the picking distance along the VIFs was set to 165 Å (55 Å), resulting in 390,297 (1,148,072) segment coordinates. Next, on the basis of the segment coordinates, two stacks of subtomograms were reconstructed from the CTF-corrected tilt series with the TOM toolbox. The dimensions of the subtomograms were 65 × 65 × 65 nm3 and 38 × 38 × 38 nm3, respectively, and the voxel size was 3.44 Å.
Subsequently, APT scripts were applied to project the subtomograms, using a projection thickness of 331 Å for the first set and 220 Å for the second set. The size of the subtomogram projections derived from the first and second coordinate sets were 65 × 65 nm2 and 38 × 38 nm2, respectively.
Initial estimate of helical symmetryThe VIF segments were subjected to multiple rounds of unsupervised 2D classifications in RELION50,56. Two-dimensional classes in which single VIFs were not combined (for example, segments containing multiple VIFs running parallel or crossing on top of each other) or those containing false-positive VIF detections (for example, actin filaments or vesicle membranes), determined by visual inspection, were excluded. Consequently, the first particle set was concentrated to 133,780 (and the second to 615,106) VIF segments.
On the basis of the resulting 2D class averages from the first set of particles (Fig. 1f), 2D autocorrelation functions of the class averages were calculated using the TOM toolbox function tom_corr, which were then displayed as profile plots and averaged (Supplementary Fig. 2c), to measure the distance between similar features observed in the class averages (Fig. 1f, yellow asterisks). The resulting 2D class averages from the first particle set were also used to calculate an averaged power spectrum with the TOM toolbox function tom_ps, to prove that the VIFs had helical symmetry and to obtain an estimate of their helical symmetry parameter (Fig. 1g). This initial estimate was 72° for the helical twist angle and 37 Å for the helical rise.
Computational filament assemblyThe assembly of extended stretches of VIFs (computationally assembled VIFs, ca-VIFs) was based on the 2D classification of the second particle set (Supplementary Fig. 3a). For this purpose, the 2D transformation calculated for each segment (namely its in-plane rotation angle and xy translation) was inverted and applied to the respective class averages, so that the inversely transformed class averages matched the position and orientation of the segments in the tomogram image frame4,104. As a result of this operation, the ca-VIFs are represented by a series of class averages (Supplementary Fig. 3b,c), which significantly improves their signal-to-noise ratio compared with that of the raw filaments. Additionally, the ca-VIFs were unbent (Supplementary Fig. 3d), determined using a MATLAB algorithm derived from the ImageJ105 straighten function1,106.
In total, 5,205 ca-VIFs of different lengths were assembled (Extended Data Fig. 4a). We selected a subgroup of 389 ca-VIFs that were ≥353 nm long. These were boxed to a uniform length of 353 nm and were measured using autocorrelation (Extended Data Fig. 4b), to determine whether the previously detected periodicity (Fig. 1f, yellow asterisks) persists over much longer distances. This result showed a long-range periodic pattern in 2D projections of VIFs that repeats every 186.5 Å ± 26.0 Å, supporting the previous measurements taken from single class averages.
Determination of helical symmetryNext, we calculated a combined power spectrum of the ca-VIFs (Extended Data Fig. 4c). Owing to the increased resolution, the previously detected layer lines (Fig. 1g) are split into fine layer lines, and a dense spectrum of layer lines is revealed. For determination of the helical symmetry of VIFs on the basis of the power spectrum of the computationally assembled filaments, the following procedure was developed in MATLAB.
Firstly, 1-pixel-wide rows in the interval between 1/30 Å and 1/69 Å were sequentially extracted from the power spectrum and compared by cross-correlation with a zero-order Bessel function (assuming a filament radius of 55 Å). The similarity of the extracted rows with a meridional reflection was measured, indicative of the helical rise of the underlying helical assembly51. As a result, the layer line at 1/42.5 Å was identified as being related to the helical rise of VIFs (Extended Data Fig. 4d,f,g).
In the next step, the sequence of the layer lines (1/207.4 Å, 1/195.9 Å, 1/185.6 Å, 1/176.3 Å; shown in Extended Data Fig. 4c, inset), which are organized around the layer line that reflects the long-range periodic pattern found previously (Extended Data Fig. 4b), were related to a similar sequence of layer lines found around the meridional reflection (1/42.5 Å, 1/40.1 Å, 1/37.9 Å, 1/36.3 Å; shown in Extended Data Fig. 4d). To this end, the relationship n = P / hr was applied, which connects the number of asymmetric units (n) with the helical pitch (P) and helical rise (hr) of a helical assembly. In particular, we searched for the optimal n value that relates the two given sequences of layer lines by P = n × hr, interpreting the first sequence as layer lines associated with the helical pitch of VIFs and the second sequence as layer lines associated with the helical rise of VIFs. The result of this calculation was n = 4.8824 (Extended Data Fig. 4e). A helical twist angle of 73.7° (that is 360° / n) and a helical rise of 42.5 Å were determined on the basis of the power spectrum of ca-VIFs.
However, owing to the high density and complexity of the layer line spectrum, we could not index the Bessel peaks51, so the uniqueness of this solution needed to be proved by other means. Therefore, we conducted extensive helical 3D classifications and helical-symmetry searches on the basis of single-particle data, which converged independently to a numerically identical solution (see below).
Cryo-EM of human VIFs polymerized in vitroHuman full-length vimentin was expressed in a transformed BL21 Escherichia coli strain and isolated from inclusion bodies, as described previously107. The protein was stored in 8 M urea, 5 mM Tris HCl (pH 7.5), 1 mM EDTA, 10 mM methylamine hydrochloride (Merck) and approximately 0.3–0.5 mM KCl at −80 °C.
To reassemble human vimentin, the protein was subjected to a stepwise dialysis in phosphate buffer at pH 7.5. A dialysis tube with a 12–14-kDa cutoff (Serva) was rinsed three times with dialysis buffer 1 (6 M urea, 2 mM phosphate buffer). Next, 200 µl of purified vimentin (0.2 mg ml–1) in 8 M urea was added to the dialysis tube, which was dialyzed for 1 h at RT against dialysis buffer 1 under gentle magnetic mixing. The buffer was exchanged three times in 1-h intervals with fresh dialysis buffer, with decreasing concentrations of urea (dialysis buffer 2: 4 M urea, 2 mM phosphate buffer; dialysis buffer 3: 2 M urea, 2 mM phosphate buffer; dialysis buffer 4: 1 M urea, 2 mM phosphate buffer). In the last step, the buffer was exchanged with 2 mM phosphate buffer, and vimentin was incubated for 2 h at RT.
VIF formation was initiated by dialyzing vimentin with 2 mM phosphate buffer containing 100 mM KCl at pH 7.5 at 37 °C overnight. To prepare samples for cryo-EM, 4 µl of filamentous vimentin at a concentration of 0.1 mg ml–1 was applied to glow-discharged, carbon-coated copper grids (Cu R2/1, 200 mesh, Quantifoil) and vitrified in liquid ethane using a manual plunge freezing device.
The VIF grids were imaged using a 300 kV Titan Krios G3i cryo-electron microscope (Thermo Fisher Scientific) equipped with a K3 direct electron detection camera (Gatan) mounted on a Bio Quantum Energy Filter (Gatan). Dose-fractionated micrographs were acquired with a 100-µm objective aperture and zero-loss energy filtering using a 20-eV slit width, at a magnification of ×130,000, with a pixel size of 0.34 Å in super-resolution mode using EPU (Thermo Fisher Scientific). A defocus range of −0.8 to −2.8 μm was chosen. The frame exposure time was set to 0.013 s, with a total exposure time of 1 s per frame (75 frames in total), resulting in a total electron dosage per dose-fractioned micrograph of ~62 e–/Å2. The micrographs were corrected for beam-induced motion with RELION56 and binned by Fourier cropping, resulting in 12,160 micrographs (Supplementary Fig. 4) with a pixel size of 0.68 Å. For CTF estimation, Gctf108 was used. The estimated maximal resolution per micrograph histogram was optimized as a function of the amplitude contrast fraction parameter. This procedure yielded an improved CTF model to describe the data with an amplitude contrast fraction of 0.2, as compared with the default value of 0.1.
Single-particle reconstruction of VIFsFilament picking was performed with crYOLO109. We used a neural network trained on in-vitro-polymerized keratin K5 and K14 IFs4, which were acquired using identical cryo-EM microscopy parameters to those used to acquire the VIFs, and we applied this neural network to the VIF micrographs. In the prediction step, the filament mode was activated, with an initial box distance between the VIF segments of 27.2 Å. Directional method convolution was used, and a search range factor of 1.41 and a filament and mask width of 14 nm were specified. As a result, 130,094 VIFs were picked from the micrographs, which were subdivided in 1,462,717 VIF segments with a particle box size of 38 × 38 nm2.
For initial particle sorting and cleaning, the segments were extracted with a pixel size of 6.8 Å and subjected to multiple rounds of unsupervised 2D classification in RELION56,57. In between rounds of 2D classification, the 2D classes combining segments of low quality (for example, segments containing overlapping or unraveling VIFs or carbon edges), determined by visual inspection, were excluded, therefore reducing the number of particles gradually to 801,585 good VIF segments. At this stage, the 2D classes (Supplementary Fig. 5a) already showed the characteristic pattern described above, in which one filament wall appears more pronounced in projection than does its counterpart (Fig. 1f and Supplementary Figs. 2d and 3a). However, at the same time, the 2D classes showed that VIFs exhibit pronounced variability in diameter, similar to what we previously observed for keratin K5 and K14 IFs4.
Because the diameter of a helical filament is coupled to its helical symmetry, multiple rounds of helical 3D classifications were performed in RELION56,57 with the aim of reducing heterogeneity in the particle set. As initial 3D template for helical 3D classifications a rotational symmetric filament tube was reconstructed from the VIF segments using 90° as approximation for the tilt angle, the refined psi angle from the previous 2D classification step and a random angle as the rot angle4. As a starting point for the helical symmetry search, an initial twist angle of 72° and an initial helical rise of 37 Å were set (Fig. 1g), and this helical symmetry was also imposed on the initial 3D template. The actual helical-symmetry search was performed in an interval between 50° to 100° for the helical twist and 30 Å to 57 Å for the helical rise, capturing possible helical assemblies based on four or six protofibrils.
In between the rounds of helical 3D classification, the 3D classes that converged to the borders of the search interval were removed, therefore reducing the number of particles gradually to 520,902 good VIF segments. In the final round of helical 3D classification (Supplementary Fig. 5b,c), the mean helical symmetry calculated from all the VIF 3D classes was 73.4° for the helical twist angle and 42.1 Å for the helical rise. These parameters were used as the initial twist and rise values for subsequent local helical symmetry searches during 3D refinement.
The processing of the particles was iteratively switched in RELION between 3D auto-refine and 3D classification jobs56,57. VIF segments at this stage were extracted with a pixel size of 1.0 Å. For 3D auto-refine jobs, the angular sampling was reduced to 1.8°, with local angular searches starting from 0.9°. Local helical symmetry search was switched on. Subsequently, a 3D classification job without image alignment was conducted. The 3D classes that were carried over to the next 3D auto-refine job were selected on the basis of visual inspection. Additionally, particles converging to a distance of <12 Å between neighboring segments were removed. The number of particles was gradually reduced to a final number of 236,920 good VIF segments. The final local helical-symmetry search converged to a helical twist angle of 73.7° and a helical rise of 42.5 Å, which is numerically identical to the helical-symmetry values previously determined by power spectrum analysis (Extended Data Fig. 4).
To improve the resolution of the VIF structure, a reference mask was applied during subsequent 3D auto-refine runs (Supplementary Fig. 6c). This mask was based on an intermediate VIF structure that was low-pass-filtered to 30 Å. In this structure, the luminal fiber was removed using UCSF Chimera103, with the aim of reducing the structural heterogeneity in the averaging volume. The RELION command relion_mask_create was used to extend the binary reference mask by 6 voxels and add a soft edge of 12 voxels. The result of this masked 3D auto-refine job with applied helical symmetry (helical twist 73.7°, helical rise 42.5 Å) was sharpened with LocalDeblur110. The LocalDeblur parameter ʎ and K were set to 1.0 and 0.025, respectively, and the local-resolution map was calculated using ResMap111.
For the final 3D auto-refine job, the reference mask described above was kept constant and the sharpened structure described above was used as initial reference. However, in contrast to previous 3D auto-refine jobs, helical symmetry was relaxed and not applied during final 3D refinement. The resulting VIF 3D structure reached a resolution of 7.2 Å (Supplementary Fig. 6f).
To visualize the increased homogeneity of the VIF segments that were used in the final 3D auto-refine job, the segments were combined with a 2D classification job in RELION. Here, the 2D alignment of the particles was fixed to the translations and in-plane rotations that were calculated in the final 3D auto-refine job (Supplementary Fig. 5d).
The final VIF 3D structure (Fig. 2 and Supplementary Fig. 6) was sharpened using LocalDeblur (ʎ = 1.0, and K = 0.025), helical symmetry was imposed (helical twist 73.7°, helical rise 42.5 Å), and the structure was low-pass-filtered to 7.2 Å. The local-resolution distribution of the map was calculated with ResMap (the mean resolution of all analyzed voxels was 5.2 Å) and interpreted as a measure for local structural plasticity (Fig. 2e and Supplementary Fig. 6e).
Cryo-EM of in-vitro-polymerized human VIFs-ΔTThe gene encoding human vimentin ΔT (amino acids 1–410) was cloned in pEt24d(+), and the protein was expressed in Rosetta2 pLysS as inclusion bodies107. The protein was purified in multiple steps, consisting of isolation of the inclusion bodies, guanidinium chloride-based solubilization and size-exclusion chromatography of the recombinant ΔT vimentin. In brief, the cell extract obtained after sonication of the frozen bacteria pellet in a buffer containing Tris HCl 50 mM pH 8, NaCl 200 mM, glycerol 25%, EDTA 1 mM, lysozyme 10 mg ml–1, MgCl2 20 mM, DNase 1 8 µg ml–1, RNase A 40 µg ml–1, NP40 1%, deoxycholic acid sodium 1% and one cOmplete Protease Inhibitor Cocktail tablet (Roche) was centrifuged for 30 min at 12,000g and 4 °C. The inclusion-bodies pellet was washed three times (or more if dark material persisted) using cycles of resuspension and centrifugation (30 min at 12,000g, at 4 °C) and a buffer containing Tris HCl 10 mM, pH 8, Triton X-100 0.5 %, EDTA 5 mM, DTT 1.5 mM and one cOmplete Protease Inhibitor Cocktail tablet. As a last step, the inclusion-bodies pellet was washed in a buffer containing Tris HCl, pH 8 10 mM, EDTA 1 mM, DTT 1.5 mM and one cOmplete Protease Inhibitor Cocktail tablet. The vimentin ΔT protein was then solubilized with 6 M guanidine hydrochloride in 10 mM Tris HCl pH 7.5, and clarified for 30 min at 10,000g at 4 °C. The vimentin-containing supernatant was collected. A size-exclusion chromatography using Superdex 200 Increase 10/300 GL (Cytiva) was performed. Protein purity was checked using SDS–PAGE.
The protein concentration was adjusted to 0.2–0.4 mg ml–1, and VIF-ΔT filaments were reconstituted by serial dialysis of 30 min at 22 °C using buffers of decreasing concentration of guanidine hydrochloride, followed by an overnight dialysis step at 4 °C in guanidine-hydrochloride-free buffer. The dialysis buffers were composed of 5 mM Tris HCl pH 7.5, with 1 mM EDTA, 0.1 mM EGTA and 1 mM DTT, containing 4, 2 and 0 M guanidine hydrochloride. The dialyzed protein solution was used for VIF-ΔT assembly by dialysis using a high-salt buffer containing 10 mM Tris HCl pH 7.5 with 100 mM KCl.
To prepare samples for cryo-EM, 3 µl of the VIF-ΔT solution was applied onto glow-discharged holey carbon EM grids (Cu R2/1, 200 mesh, Quantifoil), which were subsequently blotted manually and plunge frozen in liquid ethane.
For imaging, the VIF-ΔT grids the same EM setup were used as before, with the only difference being that the dose-fractionated micrographs were recorded in counting mode. The micrographs were corrected for beam-induced motion with MotionCor2 (ref.
留言 (0)