
記住我

To define the boundaries of the regulatory element in the BbOhmm promoter involved in response to oxidative stress, the entire BbOhmm open reading frame (ORF) along with different lengths of upstream sequences (1008–2500 bp) were cloned and reintroduced into the (previously constructed) ΔBbOhmm strain [2], with functionality of the upstream regulatory region tested by complementation of the ΔBbOhmm oxidative stress phenotype. Tolerance of the ΔBbOhmm strain to hydrogen peroxide (H2O2) was restored to wild type levels using at least 1100 bp of upstream promoter (from ATG starts site) sequence but not by sequences containing 1000 bp or less (Additional file 1: Fig S1). To further define the regulatory element, upstream regions corresponding to 1094, 1088, 1082, 1062, and 1008 bp (+ the BbOhmm ORF) were transformed into the ΔBbOhmm strain. BbOhmm with the 1094 bp and 1088 bp constructs, but not with any of the others (e.g., 1082 bp, 1062 bp, and 1008 bp) were able to complement the H2O2 phenotype of the ΔBbOhmm strain to wild type, thus defining the regulatory element to − 1087 to − 1083 bp of sequence upstream of coding region, ‘ATATC’. To determine the contribution of the “ATATC” element, the sequence was mutated to “CCCTC”, resulting in loss of the ability of the promoter to be properly activated and complement the H2O2 phenotype of the ΔBbOhmm strain (Additional file 1: Fig S1).
To isolate potential transcription factors (TFs) that could bind to the BbOhmm promoter in response to oxidative stress, yeast-one hybrid screening was performed using as the sequence target, a three-copy tandem repeat of the element “ATATC” fused to the aureobasidin A (AbA) ORF. No background binding to this element in the yeast host strain was observed since the resultant yeast strain could grow on uracil-free SD agar but not on SD agar containing 100–200 ng/ mL AbA. The resultant yeast strain was then transformed and screened with a B. bassiana cDNA library. A total of 132 clones with inserts > 200 bp were sequenced, with sequence analysis revealing 7 clones encoding four proteins containing DNA-binding domains (Additional file 1: Fig S2A). Further yeast-one hybrid screening indicated only 3 of the constructs (proteins products) showed binding to the tandem repeat element (Additional file 1: Fig S2B) and could activate transcription (Additional file 1: Fig S2C). Bioinformatic analysis indicated that these proteins were annotated as GAL4-like Zn2Cys6 proteins (BBA_01499, BBA_04239, and BBA_01981) in the B. bassiana genome, consisting of 702, 764, and 1010 amino acids that were designed as BbOsrR1, BbOsrR2, and BbOsrR3 (Oxidative stress response Regulators 1, 2, and 3), respectively. BbOsrR1 and BbOsrR3 contained GAL4-like Zn2Cys6 binuclear cluster DNA-binding domains and fungal transcription factor regulatory middle homology region (one each), whereas BbOsrR2 consisted of a large integument protein UL36-like region (PHA03247) containing a GAL4-like Zn2Cys6 binuclear cluster (Additional file 1: Fig S2D). Transcription levels of BbOsrR1 were reduced by 54.1 or 77.9% after exposure of B. bassiana to 4 mM H2O2 or 60 μM menadione (MND) for 30 min as compared to those in the normal culture (¼ SDY), respectively. However, H2O2 or MND stress resulted in significant increase in BbOsrR2 expression level (73.5 or 579.3%). BbOsrR3 expression was slightly decreased after exposure of the fungus to H2O2 stress but not to MND (Additional file 1: Fig S2E). Construction of green fluorescent protein (eGFP)-tagged fusion proteins for each TF, in which the eGFP protein was fused to the C-terminal of each protein and expressed in the B. bassiana wild type strain. The GFP signals in each strain localized in the nucleus of conidia, germlings without or with exposure to oxidative stress (4 mM H2O2 or 60 μM MND for 30 min), and in vivo blastospores (derived from infected insect) (Additional file 1: Fig S3).
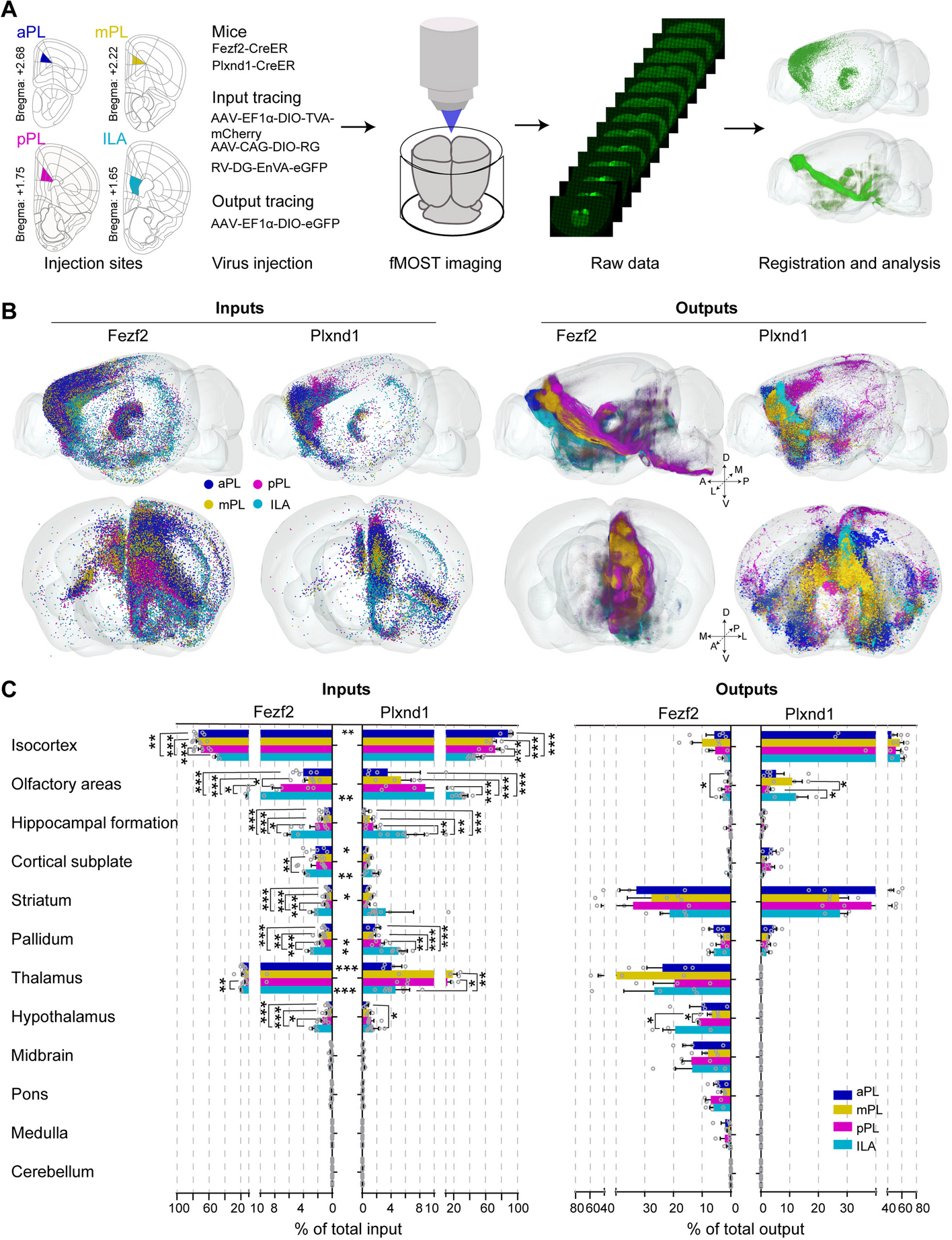
To examine the abilities of the three TFs to bind to the promoter region of BbOhmm, electrophoretic mobility shift assays (EMSA) and chromatin immunoprecipitation and quantitative PCR (ChIP-qPCR) assays were performed. EMSA tests indicated that E. coli-expressed DNA-binding domains of BbOsrR1, BbOsrR2, and BbOsrR3 could bind to the “ATATC” motif-containing promotor sequence (524 bp) of BbOhmm (Fig. 1A). To expand upon the identity of the binding sites for BbOsrR1, 2, and 3, a scanning mutation approach in which sets of the “C” and “G” nucleotides in the sequences surrounding the “ATATC” region were point mutated to (C to T) and (G to A) as follows: (i) Mutation A, − 1091 bp to − 1072 bp upstream of the coding region, 7 mutations in a 20-bp region, (ii) Mutation B, − 1105 bp to − 1090 bp and − 1073 bp to − 1055 bp upstream of “ATG” consisting of 10 changes out of 15 bp at 5′-end and 14 out of 18 bp at the 3′-flanking end, and (iii) Mutation C, containing the point mutations made in A + B in one sequence. EMSA tests were performed using the E. coli-expressed DNA-binding domains of BbOsrR1, 2 and 3 with the mutated sequences-contained promoter sequences (524 bp) of BbOhmm. All the TF DNA-binding domains could bind to the Mutation A sequence, but only BbOsrR2 bound to the Mutation B sequence, in which a putative Zn2Cys6 binding sequences “GATATCAACTGG” was found. No binding band was seen between any of the TF binding domains with the Mutation C sequence (Fig. 1A). These results indicated (i) flexibility in the binding of the BbOsrR TFs to the ATATC site (i.e., binding to ATATT sequences) and (ii) contributions of flanking sequences and / or shifted binding sites for the BbOsrR1 and BbOsrR2 TFs.
Fig. 1
Binding of BbOsrR1, 2, and 3 to BbOhmm promoter elements and BbOhmm expression in the wild type and mutant strains. A EMSA analysis of BbOsrR1, 2, and 3 binding to BbOhmm promotor regions. Wild type and three scanning mutant elements (Mutations A, B, and C) were constructed as detailed in the “Methods” section. (B-C) ChIP-qPCR analysis of BbOsrR1, 2, and 3 binding to the BbOhmm promotor region. Fungal cells were grown under no-stress and H2O2 (4 mM) or menadione (MND, 60 μM)-stress conditions over the indicated time course. D,E RT-qPCR analysis of BbOhmm expression in the wild type and ΔBbOsrR1, 2, and 3 mutant strains grown under no-stress and H2O2 (4 mM) or menadione (MND, 60 μM)-stress conditions over the indicated time course. Standard deviations (± SD) in B–E derived from triplicate experiments. Columns with the same lowercase letters above them are not significantly different (LSD test, P < 0.05)
ChIP-qPCR analysis using Myc-labeled TFs (fusion proteins) in B. bassiana revealed low enrichment of BbOsrR1 and BbOsrR3 at the BbOhmm promoter, which was not affected by addition of oxidant H2O2 (4 mM) or MND (60 μM) to the growth media. However, BbOsrR2 was highly enriched in the BbOhmm promoter, with increased signal seen over a time course of exposure of cells to H2O2 or MND peaking at 30–60 min post-stress (Fig. 1B, C). To examine functions of BbOsrR1, BbOsrR2, and BbOsrR3, targeted gene knockout (ΔBbOsrR1, ΔBbOsrR2, and ΔBbOsrR3) and their corresponding complemented strains were constructed (Additional file 1: Fig S4). RT-qPCR analysis revealed that similar patterns of BbOhmm expression were seen between the ΔBbOsrR1 and wild type strains; however, overall expression in the mutant strain was significantly lower than those in the wild type strain when grown in standard media (normal). Expression of BbOhmm was slightly induced by H2O2 but not influenced by MND stress in the ΔBbOsrR1 strain (Fig. 1D, E). No difference in BbOhmm expression was seen between the ΔBbOsrR2 and wild type strains in standard media; however, BbOhmm expression was significantly elevated when ΔBbOsrR2 cells were stressed by H2O2 (for 30 min) or MND (increasing from 30 to 120 min) as compared to the wild type strain (Fig. 1D, E), which correlated to the data of BbOsrR2 binding to the BbOhmm promoter region in response to both H2O2 and MND. Expression of BbOhmm was almost completely repressed in the ΔBbOsrR3 mutant under normal and H2O2 / MND stress conditions (Fig. 1D, E). These data suggest that BbOsrR2 acts as a (strong) negative regulator of BbOhmm in response to both H2O2 and MND, whereas BbOsrR3 acts as a positive regulator of BbOhmm expression.
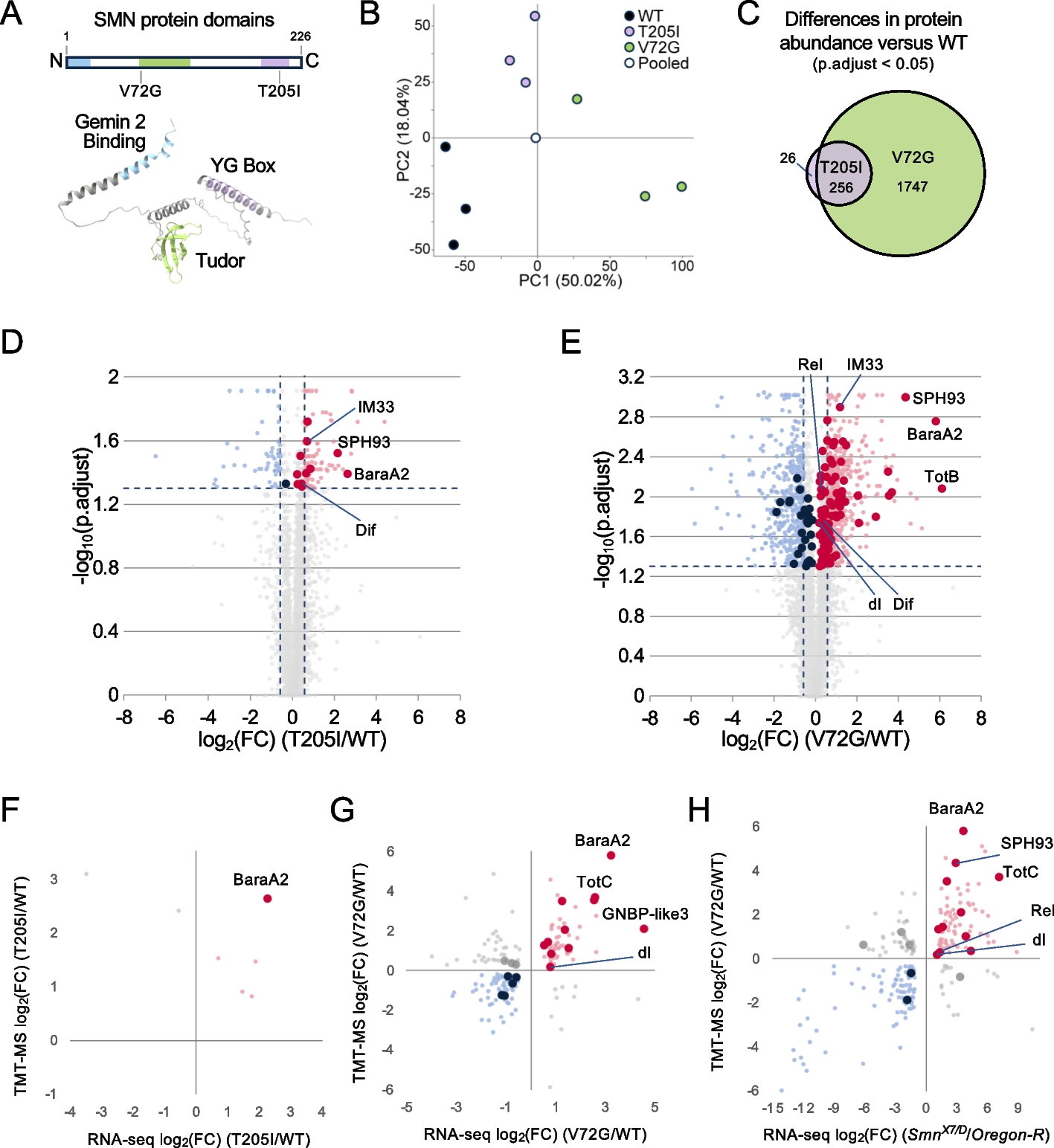
BbOsrR1, 2, and 3 differentially contribute to oxidative stress responses and virulenceIn terms of their cellular phenotypes, disruption of BbOsrR1, BbOsrR2, or BbOsrR3 did not cause any alteration in vegetative growth on the various media tested, including CZA, PDA, and ¼ SDAY, although overall conidial yield was reduced by 18.5–65.2% and 48.0–60.5% in the ΔBbOsrR1 and ΔBbOsrR2 strains, respectively, but increased by 38.0–39.4% in the ΔBbOsrR3 strain (P < 0.01). The BbOsrR2 mutation also showed delayed conidial germination, resulting in an increased median germination time (for 50% of conidial germination, GT50) from 11.8 h for the wild type strain to 13.3 h for the ΔBbOsrR2 strain (P < 0.01). However, disruption of BbOsrR3 promoted conidial germination, resulting in a decreased GT50 of 10.6 h (for ΔBbOsrR3) versus 11.8 h for the wild type strain (P < 0.01) (Additional file 1: Fig S5A-D). Oxidative stress responses of the three mutants were examined on ¼ SDAY. Inactivation of BbOsrR1 resulted in an increased tolerance to H2O2 (4 mM), reducing the relative growth inhibition (RGI) in the presence of H2O2 by 36.4% as compared to the wild type strain (P < 0.01), although sensitivity to MND remained unaffected (Fig. 2A, B). In contrast, the ΔBbOsrR2 strain displayed decreased resistance to both H2O2 and MND (RGI values increased by 146.8 and 49.0% as compared to the wild type strain, respectively, P < 0.01) (Fig. 2A, B). With respect to BbOsrR3, gene disruption led to a significantly increase in tolerances to both H2O2 and MND (RGI reduced by 31.4 and 34.1%, respectively, P < 0.01) (Fig. 2A, B). Moreover, no obvious difference in conidial germination was detected between ΔBbOsrR1, ΔBbOsrR3, and wild type strains on ¼ SDAY containing different doses of H2O2 or MND for 24 h. However, ΔBbOsrR2 conidia displayed dramatically decreased germination when stressed by the two oxidant agents, generating reduced median inhibiting concentrations (IC50) from 5.9 mM H2O2 and 69.7 μM MND for wild type strain to 3.3 mM H2O2 and 36.4 μM MND for ΔBbOsrR2 strain, respectively (Fig. 2C, D). Similar results were also examined on CZA containing H2O2 or MND (Additional file 1: Fig S6A and B), suggesting oxidative stress responses of the three mutant strains were independent of culture media.
Fig. 2
Sensitivities of B. bassiana wild type and the mutant strains to H2O2 and menadione (MND). A Vegetative growth of indicated B. bassiana strains on ¼ SDAY and ¼ SDAY containing 4 mM H2O2 or 60 μM MND for 7 days. B Calculated relative growth inhibition (RGI) of B. bassiana strains challenged with oxidative stressors for 7 days. RGI was calculated as described in “Methods” section. C Conidial germination on ¼ SDAY and ¼ SDAY containing 4 mM H2O2 or 60 μM MND for 24 h. Scale bar = 10 μm. D The dose for inhibiting 50% conidial viability (IC50) of H2O2 and menadione. IC50 was calculated using the probit regression model and the SPSS 17.0 program. Standard deviations (± SD) of data derived from triplicate experiments are shown in B and D. The asterisks (**) in the column charts denote P < 0.01 for the indicated fungal strains versus the WT (t-test), respectively
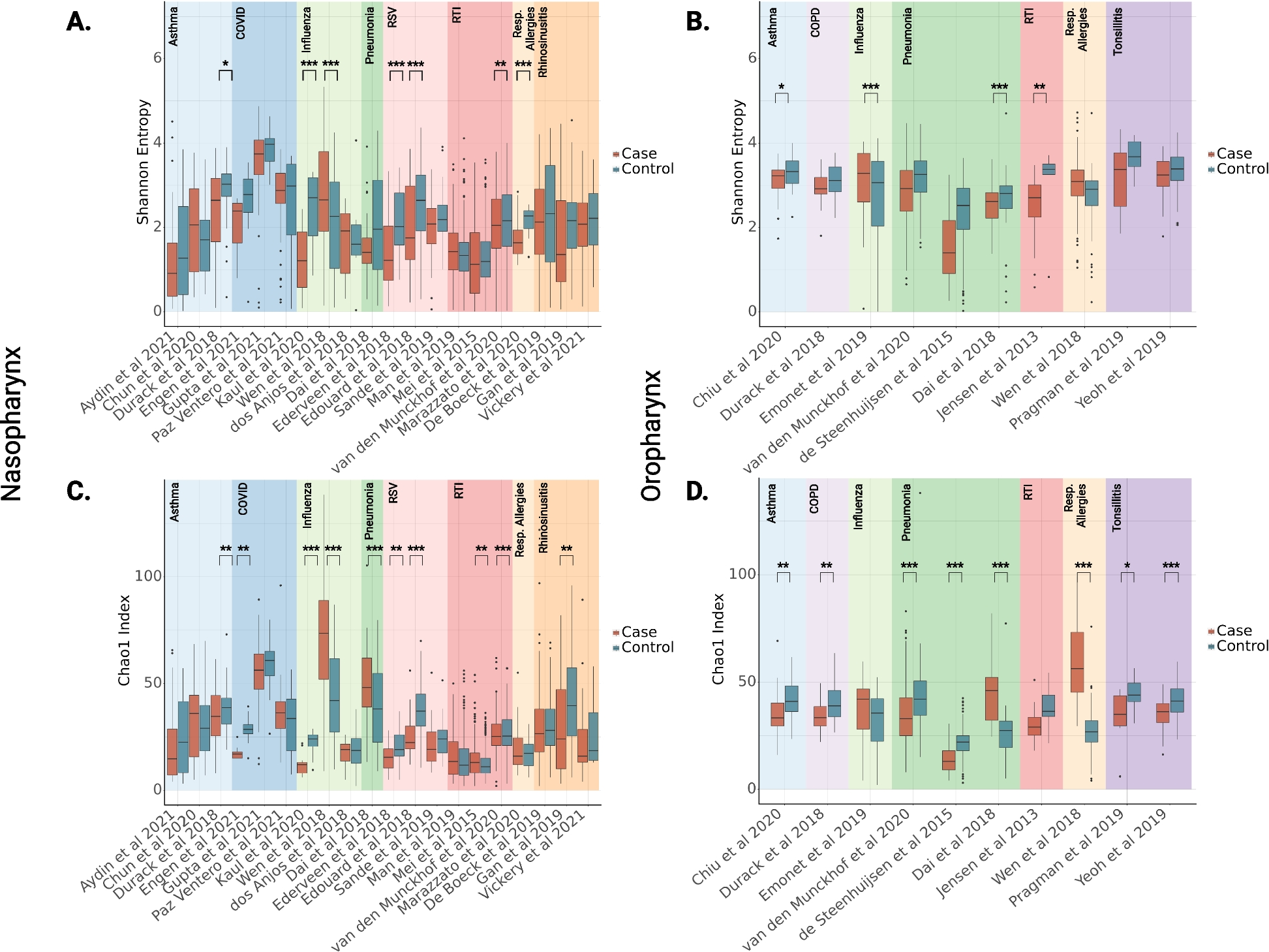
Insect bioassays using Galleria mellonella larvae as the host, revealed that loss of BbOsrR1 did not change B. bassiana virulence, but that the ΔBbOsrR2 mutant was impaired in its ability to infect and kill hosts, resulting in an increased mean lethal time to kill 50% of host insects (LT50) from 134.0 ± 2.7 h for wild type strain to 142.2 ± 2.9 h for ΔBbOsrR2 strain in topical bioassays that reflect the “nature” route of infection (5 × 106 conidia/mL) (P < 0.05), and from 67.5 ± 1.1 h for wild type strain to 74.1 ± 1.4 h for ΔBbOsrR2 strain in intrahemocoel injection assays that bypass the need for cuticle penetration to directly interact with the host (innate) immune system (2 μL 1 × 106 conidia/ mL) (P < 0.01). In contrast, the ΔBbOsrR3 was more virulent than the wild type strain, yielding an LT50 = 124.2 ± 2.9 h as compared to wild type strain (134.0 ± 2.7 h) in topical bioassays (P < 0.01), and an LT50 = 59.1 ± 1.4 h as compared to the wild type strain (67.5 ± 1.1 h) in intrahemocoel injection assays (P < 0.01) (Fig. 3). No obvious differences were seen between all the different complement strains and the wild type strain in the phenotypes examined, and unless otherwise noted, all the complemented strains were identical to the wild type strain (Fig. 3).
Fig. 3
Survival of Galleria mellonella larvae with the median lethal time (LT50) following topical application by spray of 1 mL conidial suspension (1 × 107 spores/mL) (A) and injection into the second proleg with 2 μL of 5 × 106 conidia/mL (B). All controls were treated with water. The experiments were repeated twice at different time. The survival data were plotted as Kaplan–Meyer curves and difference of the gene disruption mutant strains or reverse complementation strains from the wild type strain was analyzed using a log-rank test. LT50 was calculated using SPSS 17.0 program. The asterisks (*) and (**) in the column charts denote P < 0.05 and P < 0.01 for the indicated fungal strains versus the WT (t-test), respectively
BbOsrR1 regulates antioxidation and transcription-involved genesWe then employed RNA sequencing (RNA-seq) and chromatin immunoprecipitation sequencing (ChIP-seq) to identify target genes of BbOsrR1 in response to H2O2 stress. Global RNA-seq analysis of the ΔBbOsrR1 / wild type comparison revealed 1213 differentially expressed genes (DEGs, 695 showing increased expression and 518 showing decreased expression, fold change > 2, P < 0.001) in standard media (¼ SDY) and 1442 (688 increased / 754 decreased) in ¼ SDY + H2O2 (4 mM for 30 min) (Additional file 2: Table S2-S3). The number of upregulated and downregulated DEGs under non-stress conditions and H2O2-stress conditions are summarized in Table 1 with GO enrichment analysis of DEG datasets shown in Additional file 1: Fig S7. RT-qPCR analysis showed that transcription levels of three fungal central development pathway (CDP) genes, BrlA (BBA_07544), AbaA (BBA_00300), and VosA (BBA_01023), which are present in the downregulated ¼ SDY DEG dataset, were significantly decreased at conidiation stage (5 days on ¼ SDY agar, ¼ SDAY). Another CDP gene, WetA (BBA_06126), was also significantly downregulated in the ΔBbOsrR1 strain at this stage (Additional file 1: Fig S5E). To evaluate expression patterns of antioxidant / detoxification DEGs from RNA-seq datasets in minimal medium CZB, five genes were selected and analyzed using RT-qPCR. Expression levels of all the genes were significantly upregulated in the CZB-cultured ΔBbOsrR1 cells either under no-stress or H2O2 (4 mM for 30 min)-stress condition as compared to wild type strain (Additional file 1: Fig S6C), which were consistent with their expression patterns in the nutrient-rich ¼ SDY-derived RNA-seq data (Additional file 2: Table S2-S3).
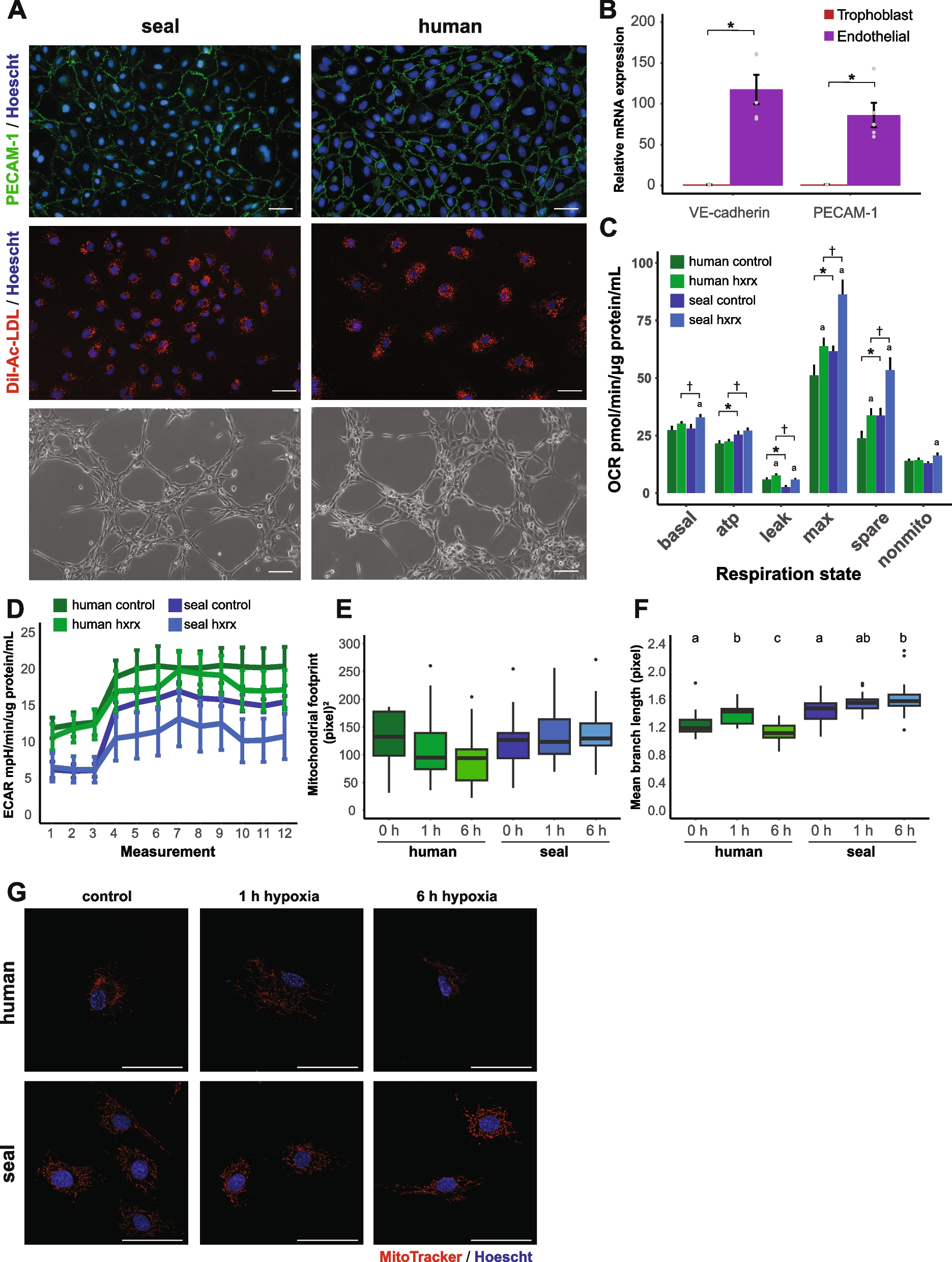
Table 1 Datasets summary of RNA-seq and ChIP-seqTo probe targets of BbOsrR1, ChIP-seq using a B. bassiana strain expressing Myc-tagged BbOsrR1 (probed with an anti-Myc antibody) was performed to compare growth of the ΔBbOsrR1 mutant in ¼ SDY to growth in ¼ SDY + H2O2 stress. These analyses revealed a total of 460 unique ChIP-seq peaks that mapped to 340 different target genes (within 1.5-kb sequences upstream of an ORF start codon) in ¼ SDY dataset, whereas only 40 target genes of BbOsrR1 were identified within 65 ChIP-seq peaks in promoter regions in ¼ SDY + H2O2 dataset (Fig. 4A, Additional file 2: Table S13-S14). MEME analysis revealed an identifiable binding sequence for BbOsrR1 in ChIP-seq peaks, as BGGYGRCGGHGGCGG with highest E value (2.2e − 4) and frequency (205/525) (Fig. 4B and Table 1). Cross analysis of the ChIP-seq with the RNA-seq datasets revealed 60 co-identified BbOsrR1 target genes (17.6%) in the standard media dataset and 6 BbOsrR1 target genes (15%) in the H2O2 stress dataset (Fig. 4C and Table 1). With respect to the former (60), target genes were enriched in antioxidant and detoxification-associated function (e.g., glutathione S-transferase, catalase (catA), oxidoreductase family protein, trafficking protein particle complex, AMP-binding enzyme, peroxisomal membrane protein 24, and nitric oxide dioxygenase), as well as transcripts involved in cell cycle and transcription. Many of these targets involved in antioxidant / detoxification were significantly upregulated in the ΔBbOsrR1 strain, supporting its role as a repressor of oxidative response pathway components. Expression of BbOsrR2 was identified as a potential direct target of BbOsrR1 (Fig. 4C); however, overall expression of BbOsrR2 was not significantly affected in the ΔBbOsrR1 strain (a slight 1.45-fold decrease that did not meet the cut-off for significance, Additional file 2: Table S13). Of the six putative direct target genes of BbOsrR1 under H2O2 stress, all were significantly upregulated in the ΔBbOsrR1 strain, which encoded the Clp1 cyclin-like protein (BBA_07582, termed BbClp1), transcription factor TFIID (BBA_05384), tyrosinase (BBA_07585), siderophore iron transporter 1 (BBA_07619), and two hypothetical proteins (BBA_09893 and BBA_10000) (Additional file 2: Table S14). Binding of purified BbOsrR1 to the promoter regions of three identified putative target genes, including CatA, BbClp1, and BbOsrR2, was verified by EMSA using respective promoter fragments from each gene and the purified protein. These bindings were not affected by addition of the mutated promoter regions of each target, in which the binding motifs “CCCGTCGCGTGGCGG” of CatA (at − 672 to − 657 bp), “CCGATTGGGTCGGCGG” of BbOsrR2 (at − 586 to − 570 bp), and “CATTACTCGAGGCGG” of BbClp1 (at − 1077 to − 1062 bp) promoters were mutated to “TTTATTATATAATAA,” “TTAATTAAATTAATAA,” and “TATTATTTAAAATAA,” respectively (Fig. 4D).
Fig. 4
ChIP-seq and RNA-seq analysis identify BbOsrR1 target genes. A Identification of BbOsrR1 (gene promoter) targets by comparison of RNA-seq and ChIP-seq under the normal and H2O2 (4 mM)-stressed conditions for 30 min. B MEME analysis indicating the likely binding sequence of BbOsrR1. C Partial annotation of identified BbOsrR1 gene targets and their expression patterns. D EMSA verification of three identified gene targets of BbOsrR1: CatA, BbOsrR2, and BbClp1 with indicated BbOsrR1 protein concentrations. The promoter region of each target harboring mutated binding motif was used as the scrambled DNA sequence control (Mutated promoter) as detailed in the “Methods” section
The BbOsrR1 target gene, BbClp1, negatively mediates oxidative stress responseBioinformatic analysis indicated that BbClp1 encodes a protein containing a cyclin box domain. The gene expression was not affected by oxidative stress as compared to that of the normal cultures from ¼ SDY (Additional file 1: Fig S2E). Construction and expression of a BbClp1::eGFP fusion reporter indicated the expected nuclear localization of BbClp1 in all the cells, including conidia, germlings without or with exposure to oxidative stress (4 mM H2O2 or 60 μM MND for 30 min), and in vivo blastospores (Additional file 1: Fig S3). To further investigate the role of BbClp1 in oxidative stress response, we constructed a target gene disruption strain ΔBbClp1 and complemented strain ΔBbClp1RC (Additional file 1: Fig S4). No significant effects in terms of vegetative growth or virulence were seen in the ΔBbClp1 mutant as compared to the wild type strain (Additional file 1: Fig S5A and B, and Fig S8). However, the strain displayed decreased conidial yield (by 21.8–22%, P < 0.01) and delayed conidial germination, with the GT50 increasing from 11.8 h for the wild type strain to 12.7 h for the ΔBbClp1 strain (P < 0.01) (Additional file 1: Fig S5C and D). The ΔBbClp1 strain also showed increased tolerances to H2O2 (4 mM) and MND (60 μM) (RGI decreased by 44.4 and 30.4% as compared to the wild type strain, respectively, P < 0.01, Fig. 2A, B). However, no obvious difference in conidial germination was examined between ΔBbClp1 and wild type strains on ¼ SDAY containing different doses of H2O2 or MND for 24 h (Fig. 2C, D). To examine the underlying mechanism of BbClp1 mediated oxidative responses, RNA-seq was used to compare gene expression patterns between the ΔBbClp1 mutant and wild type strains under the normal (¼ SDY), H2O2 (4 mM), and MND (60 μM) stress conditions (30 min). Totals of 1478, 1540, and 2059 DEGs (fold change > 2, P < 0.001) were found in comparisons between ΔBbClp1¼SDY/wild type¼SDY, ΔBbClp1H2O2/wild typeH2O2, and ΔBbClp1MND/wild typeMND, respectively (Additional file 2: Table S4-S6). The DEGs identified by comparing the ΔBbClp1 mutant grown in ¼ SDY to stress conditions indicated 461 (229 upregulated) and 1118 (614 upregulated) DEGs in the H2O2 and MND stress condition datasets, respectively. A total of 164 DEGs (92 upregulated and 72 downregulated) DEGs were shared between the ΔBbClp1 H2O2 and MND datasets (Fig. 5A). A total of 140 upregulated DEGs were found unique to H2O2 stress, and 482 unique to MND stress. Major categories of those H2O2 and MND DEGs included ribosome biogenesis, RNA processing and metabolism, and cellular component biogenesis (Fig. 5B). A total of 157 downregulated DEGs uniquely found in the H2O2-stress dataset were enriched in sugar (e.g., glucose, hexose, and monosaccharide) catabolism, protein catabolism and function, and cellular process (Fig. 5B), whereas a total of 542 downregulated DEGs uniquely found in the MND dataset were associated with protein catabolism, protein modification and function, and macromolecule catabolism (Fig. 5B). It was noticed that BbOhmm was significantly upregulated in the ΔBbClp1 strain (53-, 22-, and 15-fold in ¼ SDY, ¼ SDY + H2O2, and ¼ SDY + MND, respectively) (Additional file 2: Table S4-S6). RT-qPCR analysis revealed that transcription levels of two CDP genes, WetA and VosA, in which the latter present in the ¼ SDY DEG dataset (Additional file 2: Table S4), were significantly decreased in the ΔBbClp1 strain at conidiation stage (5 days on ¼ SDAY) (Additional file 1: Fig S5E). Moreover, expression levels of several antioxidant / detoxification genes were significantly upregulated in the ΔBbClp1 cells cultured in CZB or CZB containing H2O2 (4 mM) or MND (60 μM) for 30 min as compared to those in wild type strain (Additional file 1: Fig S6C), which were in line with their expression patterns in the ¼ SDY-derived RNA-seq data (Additional file 2: Table S4-S6).
Fig. 5
Transcriptomic analysis of the differently expressed genes (DEGs) in the ΔBbClp1 strain versus the wild type strain under the normal and oxidative stress (4 mM H2O2 or 60 μM menadione, MND) conditions for 30 min. A Venn diagram of DEGs under the normal and oxidative stress conditions. B GO enrichment of H2O2 and MND-induced DEGs as compared to those DEGs under the normal condition
BbOsrR2 controls cell detoxification genes and negatively regulates the Fus3 MAPK pathwaySimilar to experiments probing targets of BbOsrR1, ChIP-seq, and RNA-seq were used to isolate targets of BbOsrR2 regulation. RNA-seq analysis revealed that loss of BbOsrR2 led to total of 960 (616 downregulated), 971 (472 downregulated), and 1051 (572 downregulated) DEGs when compared to the wild type strain (fold change > 2, P < 0.001) under no-stress (¼ SDY), H2O2, and MND-stress conditions, respectively (Additional file 2: Table S7-S9). The number of upregulated / downregulated DEGs under no-stress condition and specifically induced by H2O2- and MND stress are summarized in Table 1 and GO enrichment analysis of those DEG datasets are shown in Additional file 1: Fig S7. RT-qPCR analysis revealed that transcription levels of three CDP genes, AbaA, WetA, and VosA, in which WetA present in the downregulated ¼ SDY DEG dataset (Additional file 2: Table S7), were significantly decreased in the ΔBbOsrR2 strain at conidiation stage (5 days on ¼ SDAY) (Additional file 1: Fig S5E). In addition, expression of five antioxidant / detoxification genes were dramatically repressed in the ΔBbOsrR2 cells cultured in CZB or CZB containing H2O2 (4 mM) or MND (60 μM) for 30 min as compared to those in wild type strain (Additional file 1: Fig S6C), which were in line with their expression patterns in the ¼ SDY-derived RNA-seq data (Additional file 2: Table S4-S6).
To identify direct promoters / target genes of BbOsrR2 in response to oxidative stress, BbOsrR2 binding sites were detected using a global genome-wide ChIP-seq screen with a B. bassiana strain expressing a Myc-tagged-BbOsrR2 fusion protein. This screening yielded a total of 1389, 1106, and 503 ChIP-seq peaks (within 1.5 kb nucleotides upstream of an ORF, P < 0.001) in cells grown in SDY, ¼ SDY + H2O2, and ¼ SDY + MND, which were mapped to 1109, 926, and 285 different genes (Additional file 2: Table S15-S17), respectively. Comparison of the ChIP-seq to the RNA-seq data revealed 133 (12%), 146 (15.8%), and 64 (22.5%) target genes found in all datasets for the ¼ SDY, ¼ SDY + H2O2, and ¼ SDY + MND conditions, respectively. Of these, 94 / 39, 84 / 62, and 10 / 54 targets were significantly upregulated / downregulated in the ΔBbOsrR2 strain under the three conditions, respectively (Fig. 6A and Table 1). MEME analysis revealed a putative identifiable binding sequence for BbOsrR2 from the ChIP-seq data to be CKYCGBSGYCG with highest E value (3.7e − 5) and frequency (267 / 2958) (Fig. 6B and Table 1). Target genes under the no-stress (¼ SDY) condition were mainly associated with transporter activity, oxidoreductase activity, and organic acid metabolic processes, in which ~ 83% (29 / 35) were significantly upregulated in the ΔBbOsrR2 strain (Fig. 6C). One target was the dual specificity phosphatase gene, Msg5 (BBA_09275), that was significantly upregulated in the ΔBbOsrR2 mutant (Fig. 6C and Additional file 2: Table S15). Yeast Msg5 specifically dephosphorylates threonine and tyrosine residues in the Mpk1 and Fus3/Kss1 MAP kinases to regulate cell wall integrity (CWI) and pheromone responses, respectively [45]. Examination of the RNA-seq dataset revealed that the Fus3 pathway genes, Ste4 (BBA_03738), Fus3 (BBA_01244), and Cdc28 (BBA_02861), were significantly upregulated in the ΔBbOsrR2 strain, suggesting participation of BbOsrR2 in the Fus3 MAPK pathway as a negative regulator (Fig. 6E). When fungal cells were stressed by H2O2, most BbOsrR2 targets (25 / 28) were distributed in terms corresponding to membrane functioning and organization, with 15 and 10 genes significantly upregulated and downregulated, respectively, in the ΔBbOsrR2 strain, that mainly included transporter and detoxification genes. In addition, three signaling pathway-involved genes were identified, including two upregulated genes, encoding a G-protein-coupled receptor (BBA_02917) and a G-patch domain-containing protein (BBA_07914), respectively, and one downregulated gene, namely, an integral membrane Pth11-like (BBA_06906) gene. BbOhmm was also identified as a target that was significantly upregulated in the ΔBbOsrR2 strain (Fig. 6C and Additional file 2: Table S16). Under the MND-stress condition, targets were enriched in membrane activity and functioning (transporter activity), and ATP binding activity, in which 15 of 17 targets were found to be downregulated in the ΔBbOsrR2 mutant (Fig. 6C and Additional file 2: Table S17). These data were consistent with the decreased tolerance of ΔBbOsrR2 strain to oxidative stress. One pheromone-regulated membrane protein (BBA_03054) and one SRF-type transcription Mcm1 (BBA_06763) genes were significantly upregulated, in which the former is involved in the Fus3-MAPK pathway, while the latter functions in cell cycle and stress response [46]. Binding of purified BbOsrR2 to the promoter regions of three identified putative target genes, including Msg5, a hypothetical protein gene (BBA_06338), and Mcm1 was verified by EMSA using respective promoter fragments from each gene and the purified protein. These bindings were not affected by addition of the mutated promoter regions of each target, in which the motifs “CCGCCTCTTCCC” of Msg5 (at − 893 to − 881 bp), “GCAAGGCCGTGTG” of Mcm1 (at -439 bp to -426 bp), and “GTCTCGCCGACTGT” of BBA_06338 (at − 1972 bp to − 1958 bp) promoters were mutated to “TTATTTTTTTTT,” “ATAAAATTATATA,” and “ATTTTATTAATTAT,” respectively (Fig. 6D).
Fig. 6
ChIP-seq and RNA-seq analysis identify BbOsrR2 target genes. A Identification of BbOsrR2 (gene promoter) targets by comparison of RNA-seq and ChIP-seq under the normal, H2O2 (4 mM), and menadione (MND, 60 μM)-stressed conditions for 30 min. B MEME analysis indicating the likely binding sequence of BbOsrR2. C Partial annotation of identified BbOsrR2 gene targets and their expression patterns. D EMSA verification of three BbOsrR2 targets: Msg5, a hypothetical protein gene (BBA_06338) and Mcm1 with indicated BbOsrR2 protein concentrations. The promoter region of each target harboring mutated binding motif was used as the scrambled DNA sequence control (Mutated promoter) as detailed in the “Methods” section. E Model of BbOsrR2 regulation of the Fus3-MAP kinase pathway via (negative) targeting of Msg5 and Mcm1, affecting expression of the Ste4, Fus3, and Cdc28 genes. Upregulated genes in the ΔBbOsrR2 mutant are shadowed in red
BbOsrR3 regulates oxidative response genes and the Fus3-MAPK pathwaySimilar to the experiments performed for BbOsrR1 and BbOsrR2, RNA-seq, and ChIP-seq were used to identify BbOsrR3 target genes. RNA-seq experiments revealed 1799 (1016 / 783, upregulated / downregulated), 1725 (871 / 854), and 2620 (1243 / 1377) DEGs (fold change > 2, P < 0.001) in the ΔBbOsrR3 mutant when compared to the wild type strain during growth in ¼ SDY, ¼ SDY + H2O2, and ¼ SDY + MND, respectively (Fig. 7A). The number of upregulated and downregulated DEGs under no-stress condition, specifically induced by H2O2 stress and MND stress, are summarized in Table 1 with GO enrichment analysis of these DEGs shown in Additional file 1: Fig S7. BbOhmm was significantly downregulated in the ΔBbOsrR3 strain (30, 21, and 13-fold under ¼ SDY, ¼ SDY + H2O2, and ¼ SDY + MND conditions, respectively (Additional file 2: Table S10-S12). RT-qPCR analysis revealed that transcription levels of CDP gene, AbaA, were significantly increased but VosA was decreased in the ΔBbOsrR3 strain at conidiation stage (5 days on ¼ SDAY) (Additional file 1: Fig S5E), which was in line with their transcription patterns in the ¼ SDY DEG dataset (Additional file 2: Table S10). Moreover, expression levels of several antioxidant / detoxification genes were significantly upregulated in the ΔBbOsrR3 cells cultured in CZB or CZB containing H2O2 (4 mM) or MND (60 μM) for 30 min as compared to those in wild type strain (Additional file 1: Fig S6C), which were consistent with their expression patterns in the ¼ SDY-derived RNA-seq data (Additional file 2: Table S10-S12).
Fig. 7
ChIP-seq and RNA-seq analysis identify BbOsrR3 target genes. A Identification of BbOsrR3 (gene promoter) targets by comparison of RNA-seq and ChIP-seq under the normal, H2O2 (4 mM), and menadione (MND, 60 μM)-stressed conditions for 30 min. B MEME analysis indicating the likely binding sequence of BbOsrR3. C Partial annotation of identified BbOsrR3 gene targets and their expression patterns. D EMSA verification of two identified BbOsrR3 targets: Fus3 and a hypothetical protein gene (BBA_06338) with indicated BbOsrR3 protein concentrations. The promoter region of each target harboring mutated binding motif was used as the scrambled DNA sequence control (Mutated promoter) as detailed in the “Methods” section. E Model of BbOsrR3 regulation of the Fus3-MAP kinase pathway via (negatively) targeting Fus3 and affecting expression of the Ste2, Sst2, and Msg5 genes. Upregulated genes in the ΔBbOsrR3 mutant are shadowed in red
BbOsrR3 binding sites were identified via ChIP-seq using a B. bassiana strain expressing Myc-tagged BbOsrR3, yielding 47 (that could be mapped to 43 different genes), 44 (mapped to 36), and 75 (mapped to 56) ChIP-seq peaks (within 1.5 kb nucleotides upstream of an ORF, P < 0.001) when cells were grown in ¼ SDY, ¼ SDY + H2O2, and ¼ SDY + MND, respectively (Additional file 2: Table S18-S20). Comparison of the ChIP-seq with the RNA-seq datasets indicated expression of 10 (8 upregulated / 2 downregulated, 23.3% of the total ChIP sets), 7 (5 / 2, 19.4%), and 14 (10 / 4, 25.0%) genes as BbOsrR3 dependent under the three conditions, respectively (Fig. 7A and summarized in Table 1). MEME analysis indicated the putative DNA-binding sequence for BbOsrR3 to be CCYCSSYRAC with highest E value (8.8e − 3) and frequency (53/166) (Fig. 7B and Table 1). Many upregulated targets were involved in signal transduction and regulator (e.g., Fus3 MAPK, PTH11 integral membrane protein, F-box domain-containing protein, protein kinase protein, bZIP transcription factor, and LMBR1 domain-containing protein), lipid metabolism (sphingolipid long chain-base responsive protein, fatty acid hydroxylase superfamily protein, and phosphotransferase enzyme family protein), cell resistance and detoxification (LPG synthase protein, major facilitator superfamily transporter and metalloreductase), xylose / nitrogen source utilization and virulence (hexose transportor-like protein, small secreted protein, PAP superfamily protein and subtilase-like protein), and vascular remodeling (ring 10 protein) (Fig. 7C and Additional
留言 (0)