
記住我



Previously, we identified the CSN complex in M. oryzae and found that its component Csn6 is involved in fungal development, autophagy, ubiquitination, and pathogenicity [26]. To further investigate the biological function of the CSN complex in M. oryzae, high-throughput target gene deletion strategies were used to knock out MoCSN1, MoCSN2, MoCSN3, MoCSN4, MoCSN5 and MoCSN7a in M. oryzae 70-15 (wild type) (Figure S1A). At least four positive knockout transformants with similar phenotypes were obtained for each CSN gene, and single-copy validation was performed (Figure S1B and C). To determine the roles of various CSN subunits in the growth of rice blast fungus, the obtained CSN mutants and 70-15 were inoculated on complete medium (CM) or minimum medium (MM) for 9 days. As shown in Figure S2A and B, compared with 70-15, the destruction of any CSN subunit led to a sharp decrease in mycelial growth. After the corresponding CSN gene was replenished, the mycelial growth rates of all replenished strains (Mocsn1-C, Mocsn2-C, Mocsn3-C, Mocsn4-C, Mocsn5-C and Mocsn7a-C) were restored to those of the wild type (WT) strain (Figure S2A and B). Further phenotypic analysis indicated that all subunits of the CSN are necessary for spore production and complete pathogenicity (Figure S2C and D), indicating that the CSN complex is a key regulator of nutritional growth, spore production, and pathogenicity in rice blast fungus.
Although previous studies have shown that the CSN complex is closely related to the pathogenicity of phytopathogenic fungi, the specific regulatory mechanisms involved are still unclear. MoCsn6 is involved in the regulation of autophagy in M. oryzae [26]. Therefore, we used yeast two-hybrid methods to preliminarily identify interactions between CSN subunits and autophagy-related proteins. Yeast two-hybrid experiments showed that MoCsn1, MoCsn3, and MoCsn4 all interacted with MoAtg5, MoAtg6, MoAtg14, MoAtg16, and MoAtg17, that MoCsn4 interacted with MoAtg5, MoAtg12, and MoAtg18, that MoCsn5 interacted with MoAtg6 and MoAtg14, and that MoCsn7a interacted with MoAtg5 and MoAtg6 (Figure S3A). These results suggest that the CSN complex may mediate the pathogenic ability of M. oryzae by regulating autophagy, but the exact mechanism still needs to be explored.
Identification of the Csn5 protein in rice blast fungusIn this study, we focused on MoCsn5, the key subunit of the CSN complex, to explore its function in M. oryzae. The amino acid sequence (344 aa) encoded by MGG_05274 was compared in the NCBI database. The results showed that this protein had 53.31% homology with Homo sapiens Csn5 and 66.08% homology with F. graminearum Csn5 (Figure S4A); therefore, we named this protein MoCsn5. A phylogenetic tree was constructed based on the Csn5 amino acid sequences of M. oryzae (XP_003712833.1), F. graminearum (XP_011318525.1), Mus musculus (NP_038743.1), Caenorhabditis elegans (NP_500841.1), Aspergillus fumigatus (XP_755961.2), H. sapiens (NP_006828.2), and Neurospora crassa (XP_956786.1) (Figure S4B). The phylogenetic tree showed that the Csn5 of M. oryzae had greater homology with the Csn5 of F. graminearum than with those of C. elegans, M. musculus, and H. sapiens (Figure S4B). Csn5 is the fifth subunit of the CSN, which is highly conserved in eukaryotes and contains a conserved MPN domain (Figure S4C).
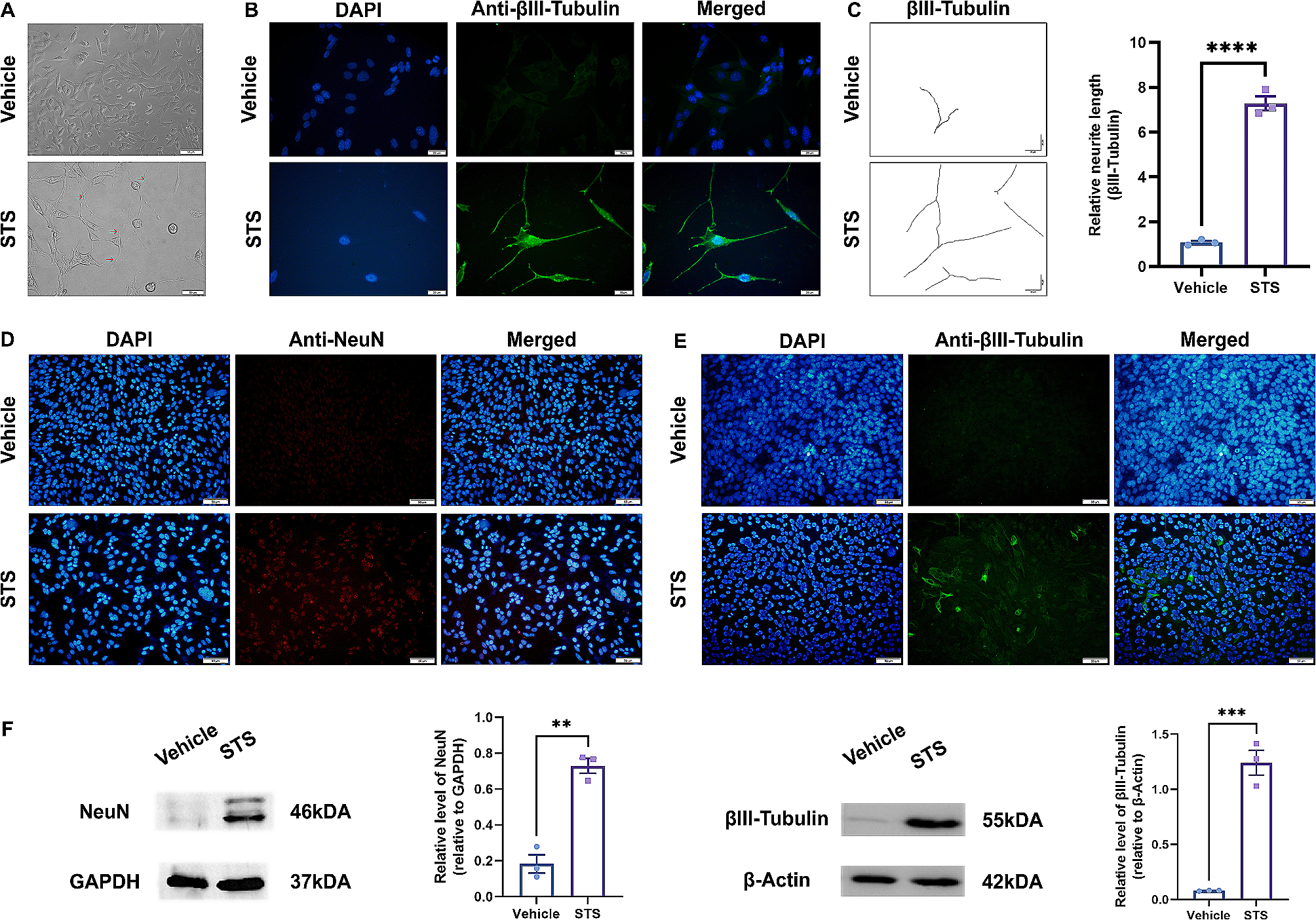
ΔMocsn5 is deficient in vegetative growth and sporulationTo further explore the basic biological functions of MoCsn5, growth and spore production assays were performed. The ΔMocsn5 strain showed a significantly slower growth rate on both CM and MM (Fig. 1A). As shown in Fig. 1B, the aerial hyphae of ΔMocsn5 were thinner than those of 70-15 and Mocsn5-C. Compared with those of 70-15 and Mocsn5-C, the colony diameter of ΔMocsn5 on CM was decreased by 22.10%, and that on MM was decreased by 51.38% (Fig. 1C). The conidium pedicels of 70-15 and Mocsn5-C show typical branching structures and bear more conidium, whereas ΔMocsn5 has few fascicular conidial pedicels, and most of them are single-branching structures with few spores (Fig. 1D). Statistical analysis revealed that the spore production of ΔMocsn5 was significantly decreased to only 0.73% of that of 70-15 (Fig. 1E). In summary, MoCsn5 was essential for the vegetative growth and sporulation of rice blast fungus.
Fig. 1
MoCsn5 is required for vegetative growth and conidiation. A Colony morphology of 70-15, ΔMocsn5 and Mocsn5-C. The strains were grown on CM and MM plates for 9 days. B Hyphal growth of 70-15, ΔMocsn5 and Mocsn5-C on solid CM. C Statistical analysis of the colony growth diameter. The data were analyzed using GraphPad Prism 8.0 software. The error bars represent the standard deviations. ***P < 0.001. D Conidiophores of 70-15, ΔMocsn5 and Mocsn5-C. The strains were cultivated in an incubator at 25 ℃ for 9 days and observed under an optical microscope. E Statistical analysis of conidia production. The data were analyzed with GraphPad Prism 8.0 software. ***P < 0.001. F Localization of MoCsn5 in the conidium, appressorium and invasive hypha. The MoH2B-mCherry vector was transformed into the complementation strain with the MoCsn5-GFP label. Conidia were harvested from colonies that were cultivated in solid CM for 9 days. The conidial suspension (5 × 104 ml−1) was dripped onto a hydrophobic film and incubated in a humid chamber at 22 °C for 24 h. The red fluorescence and green fluorescence in the spores and appressoria were observed under a fluorescence microscope. Bar: 10 μm. G The fluorescence densities of MoH2B-mCherry and MoCsn5-GFP were analyzed using ImageJ software
Then, we investigated the subcellular localization of MoCsn5. The MoCsn5-GFP (green fluorescent protein) vector was constructed and transformed into the ΔMocsn5 mutant. As shown in Fig. 1F, the green fluorescence of MoCsn5-GFP was observed in the conidium, appressorium and invasive hypha. To confirm whether MoCsn5-GFP is localized in the nucleus and cytoplasm, the nuclear location marker protein MoH2B-mCherry was transformed into the complemented strain with the MoCsn5-GFP label through an Agrobacterium tumefaciens-mediated transformation (ATMT) strategy. The green fluorescence of MoCsn5-GFP overlapped with that of the nucleus labeled with MoH2B-mCherry (Fig. 1G). These data indicate that MoCsn5 is localized in the cytoplasm and nucleus of M. oryzae.
MoCsn5 is involved in conidial germination, appressorium formation, and pathogenicity in M. oryzaeWe then performed mycelial plug pathogenicity experiments on two susceptible hosts (barley and rice). As expected, the disease spots were smaller in the ΔMocsn5 mutant than in the 70-15 and complemented strains (Fig. 2A and B). In addition, we inoculated wounded rice and barley leaves with mycelial plugs of 70-15, ΔMocsn5 and Mocsn5-C for 3 days. Moreover, 70-15 and Mocsn5-C caused severe lesions, whereas ΔMocsn5 caused smaller disease lesions than 70-15 and Mocsn5-C (Fig. 2C and D). As the infectious weapon of rice blast fungus, conidia germinate to form appressoria with sufficient penetration turgor to infect the host and thus play a vital role in virulence. To investigate whether MoCsn5 regulates the development of appressoria in M. oryzae, we induced appressoria on an artificially hydrophobic surface and observed their morphology under a microscope. As shown in Fig. 2F, the conidial morphology was not affected by MoCSN5 deletion, and typical three-celled spores remained. Compared with those of 70-15 and Mocsn5-C, the germ tube germination rate and appressorium formation rate of ΔMocsn5 were significantly decreased at 4 h postinoculation (hpi) and 24 hpi (Fig. 2G-J). At 4 hpi, the germination rate of ΔMocsn5 was only 25.3%, and the appressorium formation rate was only 6.3%, whereas the germination rate of 70-15 was 90.0%, and its appressorium formation rate was 84.3% (F ig. 2I and J). Although the observation time was extended to 24 h, the tube germination rate (37.3%) and appressorium formation rate (16.7%) of ΔMocsn5 were still significantly lower than those of 70-15 (97.3%/93%) and Mocsn5-C (96.3%/94.3%) (Fig. 2I and J). Pmk1-MAPK pathways have been reported to regulate appressorium formation in M. oryzae. The decreased phosphorylation levels of MoPmk1 in ΔMocsn5 indicate that MoCsn5 is involved in appressorium formation through the MAPK pathway (Fig. 2K).
Fig. 2
MoCsn5 is involved in conidial germination, appressorium formation, and pathogenicity in M. oryzae. A Disease spots of detached barley leaves inoculated with mycelial plugs from the 70-15, ΔMocsn5 and Mocsn5-C strains. Leaves were cultured at 25 ℃ for 4 days after inoculation. B Mycelial plugs from the 70-15, ΔMocsn5 and Mocsn5-C strains were inoculated on detached rice leaves. C Disease symptoms on wounded leaves of rice inoculated with mycelial plugs of the 70-15, ΔMocsn5 mutant and Mocsn5-C strains. D Disease symptoms on wounded leaves of barley inoculated with mycelial plugs of 70-15, ΔMocsn5 mutant and Mocsn5-C strains. E Disease spots of detached barley leaves inoculated with conidial suspensions of the 70-15, ΔMocsn5 and Mocsn5-C strains. Leaves were cultured at 25 ℃ for 4 days after inoculation. F Conidial morphology of 70-15, ΔMocsn5 and Mocsn5-C. Bar: 10 μm. G Germ tube germination assays and appressorium formation assays on hydrophobic surfaces at 4 hpi. Bar: 10 μm. H Germ tube germination assays and appressorium formation assays on hydrophobic surfaces at 24 hpi. Bar: 10 μm. I Statistical analysis of germ tube germination rates of 70-15, ΔMocsn5 and Mocsn5-C when conidia were dropped onto hydrophobic surfaces at 4 hpi and 24 hpi. The data were analyzed with GraphPad Prism 8.0. ***P < 0.001. J Statistical analysis of the appressorium formation rates of 70-15, ΔMocsn5 and Mocsn5-C when conidia were dropped onto hydrophobic surfaces at 4 hpi and 24 hpi. The data were analyzed using GraphPad Prism 8.0. ***P < 0.001. K Phosphorylation analysis of Pmk1 in 70-15 and ΔMocsn5. Pmk1 phosphorylation and Pmk1 levels were detected with phospho-Pmk1 and Pmk1 antibodies, respectively. The protein actin was used as a loading control. L The conidial suspension (5 × 104 ml−1) was dripped onto isolated leaves and incubated in a humid chamber at 25 °C. After 3 days, the leaves were decolorized with methanol and observed under an optical microscope. Bar: 10 μm. M The infection rate was quantified and statistically analyzed using GraphPad Prism 8.0 software
Then, we tested the pathogenicity of ΔMocsn5 on barley leaves using conidia. As expected, only some nonexpanding small necrotic lesions were observed on the leaves inoculated with ΔMocsn5, while the leaves inoculated with 70-15 and Mocsn5-C produced a large number of connected brown necrotic lesions (Fig. 2E). Finally, we carried out infection experiments with isolated barley leaves to further explore the progression of infection in the ΔMocsn5 mutant. Seventy-two hours after spore infection, the leaves were observed under a microscope. The main growth morphology of infected hyphae in leaves infected with 70-15, ΔMocsn5 and Mocsn5-C is shown in Fig. 2L. The infected hyphae of 70-15 and Mocsn5-C expand into multiple adjacent plant cells, whereas the appressoria of ΔMocsn5 are not infected, or the infected hyphae are limited to a single plant cell (Fig. 2L). At 72 hpi, nearly 86% of the infection structures were detected in the 70-15 strains, and 76% had expanded to neighboring cells. However, with ΔMocsn5, only approximately 12% of the infected structures formed, and 9% were still limited to the initially infected cells (Fig. 2M). These results indicate that MoCsn5 is essential for germ tube germination, appressorium development, and pathogenicity in M. oryzae.
Tolerance to external stresses is critical for the survival, appressorium formation, and invasion of rice blast fungus [39,40,41]. We found that ΔMocsn5 was more sensitive to hyperosmotic stress (KCl, NaCl, and sorbitol), oxidative stress (menadione and H2O2), and amphotericin B (an antifungal agent that disrupts membrane permeability by binding to sterols on the fungal cell membrane) than 70-15 and was not sensitive to myriocin (a sphingolipid synthesis inhibitor) (Fig. 3A-F). Fungi have been reported to respond to hyperosmotic stress through two mechanisms: the Osm1-MAPK signaling pathway and the TORC2-Ypk1 signaling pathway [42,43,44]. We further determined the phosphorylation levels of Osm1 and Ypk1 under 0.6 M NaCl treatment. The ΔMocsn5 mutant and 70-15 strains showed a consistent trend in Ypk1 phosphorylation (Fig. 3G). However, the level of Osm1 phosphorylation was lower in the ΔMocsn5 mutant than in the 70-15 strain (Fig. 3H). The phosphorylation level of Osm1 in the ΔMocsn5 mutant was also significantly lower than 70-15 after treatment with 2 μM amphotericin B (Fig. 3I), which disrupts cell membrane permeability. The above-described experimental results indicate that MoCsn5 participates in the response to hyperosmotic stress by participating in the Osm1-MAPK signaling pathway.
Fig. 3
ΔMocsn5 is defective in the maintenance of external stresses. A and B Colony morphology and relative growth rate of 70-15, ΔMocsn5 and Mocsn5-C on CM supplemented with 2 mM hydrogen peroxide (H2O2) and 100 μM menadione (VK3). ***P < 0.001. C and D Colony morphology and relative growth rate of 70-15, ΔMocsn5 and Mocsn5-C on CM supplemented with 2 μM amphotericin B (AmB) and 1.5 μM myriocin. ***P < 0.001. E and F Colony morphology and relative growth rate of 70-15, ΔMocsn5 and Mocsn5-C on CM supplemented with 0.4 M KCl, 0.6 M NaCl, and 0.8 M sorbitol. Pictures were taken at 9 days. The data were analyzed with GraphPad Prism 8.0 software. ***P < 0.001, **P < 0.01. (G) Phosphorylation level of MoYpk1 in 70-15 and ΔMocsn5. The strains were cultured in CM supplemented with 0.6 M NaCl before analysis. H Phosphorylation level of MoOsm1 in 70-15 and ΔMocsn5 strains. The strains were cultured in CM supplemented with 0.6 M NaCl before analysis. I Phosphorylation level of MoOsm1 in 70-15 and ΔMocsn5 strains. The strains were cultured in CM or treated with 2 μM amphotericin B before analysis
MoCsn5 regulates the ubiquitin‒proteasome pathway in rice blast fungusOur previous experimental results showed that MoCsn5 is a subunit of the CSN complex and participates in the assembly of the CSN complex through interaction with other CSN subunits, mediating the ubiquitin‒proteasome pathway. To further investigate the role of MoCsn5 in ubiquitination, we detected the ubiquitination levels of 70-15 and ∆Mocsn5 using an anti-ubiquitin antibody. The lower level of ubiquitination in ∆Mocsn5 suggests that MoCsn5 promotes ubiquitination (Fig. 4A). This result was further confirmed by the recovery of the ubiquitination levels in the complemented strain Mocsn5-C (Fig. 4A). MG132 is a proteasome inhibitor that inhibits the degradation of ubiquitinated proteins. After induction with MG132 for 4 h, the levels of ubiquitination of 70-15 and ∆Mocsn5 significantly increased due to the accumulation of ubiquitinated proteins (Fig. 4A). However, regardless of MG132 induction, the ubiquitination protein level of ∆Mocsn5 was significantly lower than that of 70-15 and Mocsn5-C. This finding indicates that the ability of MoCsn5 to promote ubiquitination in rice blast fungus was sustained.
Fig. 4
MoCsn5 regulates the ubiquitin‒proteasome pathway in rice blast fungus. A Ubiquitination levels of 70-15, ΔMocsn5 and Mocsn5-C at 0 h and 3 h after MG132 induction. Hyphae were grown in liquid CM for 36 h and then shifted to CM supplemented with 20 μM MG132 (a proteasome inhibitor, a tool for studying cellular degradation of the ubiquitin‒proteasome pathway) for 0 h and 3 h. B Transcript levels of MoSKP1 (MGG_04978), MoCULLIN1 (MGG_07145), MoCULLIN3 (MGG_07731) and MoCULLIN4b (MGG_14763) in 70-15 and ΔMocsn5. The error bars represent the standard deviations. Tukey’s test was used to determine significance. **P < 0.01. C The interaction between MoCsn5 and MoCullin3 was detected by yeast two-hybrid assays. pGADT7-T and pGBKT7-53 were used as positive controls. pGADT7-T and MoCsn5-BD and pGBKT7 and MoCullin3-AD served as two pairs of negative controls. D The relationship between MoCsn5 and MoCullin3 in vivo was examined by coimmunoprecipitation assays. The MoCullin3-Flag bands were detected following MoCsn5-GFP immunoprecipitation
To explore the mechanism by which MoCsn5 promotes ubiquitination, we examined the mRNA levels of ubiquitin-associated proteins in M. oryzae. The qPCR results showed that the expression levels of MoCULLIN1, MoCULLIN3, MoCULLIN4b, and MoSKP1 were significantly decreased by the elimination of MoCsn5 (Fig. 4B). This finding explains the reduced level of ubiquitination in ∆Mocsn5. In addition, the results of the interaction experiment indicated that MoCsn5 directly interacts with MoCullin3 in vivo and in vitro to regulate CRLs (Fig. 4C and D). In summary, we conclude that MoCsn5 promotes the ubiquitination of M. oryzae by regulating the components of CRLs.
MoCsn5 negatively regulates autophagyThe ubiquitin‒proteasome system and autophagy‒lysosome pathway, which are responsible for the degradation of cellular proteins, are crucial for various cellular processes, such as the growth and development of organisms [8]. Due to the important function of CSN in the ubiquitin‒proteasome system and the interaction between MoCsn5 and autophagy-related proteins, we examined the autophagy levels in 70-15 and ΔMocsn5. The GFP-MoAtg8 fusion protein was used to determine whether there was a connection between MoCsn5 and autophagy. During the fusion of the autophagosome with the lysosome, MoAtg8 on the inner membrane of the autophagosome is delivered to the vacuole for degradation, where it produces free GFP (in the vacuole); therefore, the [(GFP)/(GFP + GFP-MoAtg8)] ratio is widely used to indicate the level of autophagy. First, the subcellular localization of GFP-MoAtg8 in 70-15 and ΔMocsn5 was observed. As shown in Fig. 5A, under nutritional conditions, GFP-MoAtg8 in 70-15 was positioned around the vacuole in a spot-like manner, whereas the vacuoles in ΔMocsn5 appeared bright and uniform green without obvious dot-like localization. After 3 h of starvation induction, the vacuoles in the 70-15 strain were uniformly green overall, and the same trend was observed in the ΔMocsn5 strain (Fig. 5A). For further confirmation, a western blot analysis was conducted to detect full-length GFP-MoAtg8 and free GFP in 70–15 and ΔMocsn5. As shown in Fig. 5B, the autophagic flux in the ΔMocsn5 mutant was greater than that in the wild type.
Fig. 5
MoCsn5 negatively regulates autophagy. A Subcellular localization of GFP-MoAtg8 in 70-15 and ΔMocsn5 under nutrient and starvation conditions. Bar: 10 μm. Mycelia were stained with CMAC to label the vacuoles. B Autophagic flux analysis of GFP-MoAtg8 in 70-15 and ΔMocsn5. Total GFP-MoAtg8 and free GFP were detected by western blot. The gray values of the protein bands were analyzed with ImageJ software. The degradation rate was expressed as [GFP/(GFP + GFP-MoAtg8)]. The protein content of actin was used as a control. C Analysis of MoAtg8/MoAtg8-PE turnover in 70-15 and ΔMocsn5. The ratio of MoAtg8-PE to actin was calculated to assess the rate of MoAtg8 lipidation to yield MoAtg8-PE
Next, we evaluated the turnover rate of endogenous MoAtg8/MoAtg8-phosphatidylethanolamine (MoAtg8-PE). The conversion of microtubule-associated protein 1 light chain 3 (LC3-I, Atg8 homology) to a lapidated form (LC3-II, Atg8-PE) is a committed step in autophagosome formation. Accordingly, the LC3-II protein level is commonly used as a marker of the autophagosome number [45]. As shown in Fig. 5C, under nutritional conditions, the MoAtg8 band was stronger, whereas the MoAtg8-PE band obtained with both 70-15 and ΔMocsn5 was weaker. After 3 and 6 h of starvation induction, the MoAtg8-PE and MoAtg8 band strengths were enhanced in both the 70-15 and ΔMocsn5 strains, indicating that starvation stimulated the formation of autophagosomes (Fig. 5C). However, it is worth noting that compared to 70-15, ΔMocsn5 has stronger MoAtg8-PE and MoAtg8 bands. These results indicated that the total conversion of MoAtg8 to MoAtg8-PE was increased in the ΔMocsn5 strain, and the number of autophagosomes and autophagic flux were greater in the ΔMocsn5 strain than in the 70-15 strain.
Because MoCsn5 and MoCsn6 are involved in the regulation of autophagy and other subunits also interact with autophagy-related proteins, we are very interested in whether other subunits of the CSN complex generally have regulatory functions in autophagy. Therefore, we transfected GFP-MoAtg8 into the corresponding mutants to detect the autophagy levels. Several transfection experiments were performed, and GFP-MoAtg8 was not transfected into the ΔMocsn2 and ΔMocsn3 mutants. Compared with 70-15, the ΔMocsn1, ΔMocsn4, and ΔMocsn7a strains all showed increased levels of autophagy (Figure S3B and C). Thus, other subunits of the CSN complex also regulate autophagy in M. oryzae.
MoCsn5 inhibits autophagy by promoting K48-ubiquitination of MoAtg6Figure 5 shows increased autophagosome formation and accelerated autophagosome transport in ΔMocsn5. Yeast two-hybrid and pull-down assays revealed that MoCsn5 interacts with the autophagy-related proteins MoAtg6 and MoAtg14 in vitro (Fig. 6A-C). Co-IP experiments also confirmed that MoCsn5 interacts with MoAtg6 and MoAtg14 in vivo (Fig. 6D and E). These results indicate that Csn5 may participate in the autophagy regulation pathway in M. oryzae by regulating autophagy-related proteins.
Fig. 6
MoCsn5 inhibits autophagy by promoting the K48-ubiquitination of MoAtg6. A The interaction between MoCsn5 and MoAtg6 was detected by yeast two-hybrid assays. pGADT7-T and pGBKT7-53 were used as positive controls. pGADT7-T and MoCsn5-BD and pGBKT7 and MoAtg6-AD served as two pairs of negative controls. B The interaction between MoCsn5 and MoAtg14 was detected by yeast two-hybrid assays. pGADT7-T and pGBKT7-53 were used as positive controls, and pGADT7-T and MoCsn5-BD and pGBKT7 and MoAtg14-AD were used as two pairs of negative controls. C Pulldown assays to detect the interaction between MoCsn5 and MoAtg14 in vitro. GST-MoAtg14 and His-MoCn5, empty GST, and His-MoCsn5 were incubated sequentially with glutathione agarose gel beads with GST labels for 2 h. The final eluents were detected by western blot assays. D The relationship between MoCsn5 and MoAtg14 was examined by in vivo coimmunoprecipitation assays. The MoCsn5-Flag bands were detected following MoAtg14-GFP immunoprecipitation. E The relationship between MoCsn5 and MoAtg6 was examined by in vivo coimmunoprecipitation assays. The MoCsn5-Flag bands were detected following MoAtg6-GFP immunoprecipitation. F Interaction intensity of MoAtg6 and MoAtg14 in 70-15 and ΔMocsn5. MoCsn5 knockout was performed in 70-15 strain expressing MoAtg6-GFP and MoAtg14-Flag to ensure consistent MoAtg6 and MoAtg14 contents. The strains were lysed and immunoprecipitated with anti-GFP beads and then subjected to immunoblotting with anti-GFP and anti-Flag antibodies. The formula MoAtg14-Flag/MoAtg6-GFP was used to calculate the interaction intensity of MoAtg6 and MoAtg14 in the different strains. G Interaction intensity of MoAtg6 and MoAtg14 in 70-15 and ΔMocsn5. MoCsn5 knockout was performed in 70-15 strain expressing MoAtg6-GFP and MoAtg14-Flag to ensure consistent MoAtg6 and MoAtg14 contents. The strains were lysed and immunoprecipitated with anti-Flag beads and then subjected to immunoblotting with anti-GFP and anti-Flag antibodies. The formula MoAtg6-GFP/MoAtg14-Flag was used to calculate the interaction intensity of MoAtg6 and MoAtg14 in the different strains. H Ubiquitination levels of MoAtg14 in 70-15 and ΔMocsn5. Gene knockout of MoCSN5 was performed in 70-15 expressing MoAtg14-Flag to ensure consistent transcript levels of MoATG14. The strain was immunoprecipitated with anti-Flag beads after lysis, and a western blot analysis was then performed with an anti-Flag antibody and an anti-ubiquitin antibody. The formula ubiquitin/MoAtg14-Flag was used to calculate the ubiquitination levels of MoAtg14-Flag. I Deletion of MoCSN5 inhibited the ubiquitination and K48-ubiquitination of MoAtg6. Gene knockout of MoCSN5 was performed in 70-15 expressing MoAtg6-GFP to ensure consistent transcript levels of MoATG6. The strain was immunoprecipitated with anti-GFP beads after lysis, and a western blot analysis was then performed with anti-GFP antibody, anti-ubiquitin antibody, and anti-K48 antibody. The ubiquitination and K48-ubiquitination levels of MoAtg6-GFP were calculated as ubiquitin/MoAtg6-GFP and K48-ubiquitin/MoAtg6-GFP, respectively. J Ubiquitination levels and K48-ubiquitination levels of MoAtg6 in 70-15, ΔMocsn5 and Mocsn5-C. K The protein levels of MoAtg6 in the 70-15, ΔMocsn5 and Mocsn5-C strains were measured with an anti-Beclin1 antibody. The strains were cultured in liquid CM at 25 ℃ for 36 h. The protein content of actin was used as a control. L Transcript levels of MoATG6 in 70-15 and ΔMocsn5. The error bars represent the standard deviations
The PI3KC3-I complex is formed by Atg6/Vps30/Beclin1, Vps34, Vps15, and Atg14 at the phagophore assembly site (PAS) and is essential for proper PAS targeting of other autophagy-related proteins, such as the Atg8, Atg18, and Atg12–Atg5-Atg16 complex [46, 47]. The localization of PI3KC3-I to the PAS is largely dependent on the binding of Atg6 to Atg14 [48]. To investigate the effect of ubiquitination on the PI3KC3-I complex, we examined the ability of Atg14 to target Atg6. Interaction strength tests showed that the amount of MoAtg14 bound to MoAtg6 in ΔMocsn5 was comparable to that bound in 70-15. MoCsn5 did not affect the ability of MoAtg14 to target MoAtg6 to the PAS (Fig. 6F and G). Subsequent analyses revealed that the ubiquitination level of MoAtg14 in ΔMocsn5 was unaffected but that the ubiquitination level of MoAtg6 was lower in ΔMocsn5 than in 70-15, suggesting that deficient ubiquitination modulates the level of MoAtg6 (Fig. 6H and I).
Atg6 is a core protein in the autophagy pathway, and previous studies have shown a positive correlation between its protein expression and autophagic flux [49]. Subsequent experiments showed that the increased MoAtg6 content in ΔMocsn5 resulted in increased autophagy activity (Fig. 6K). However, the mRNA levels of MoATG6 were not significantly different between ΔMocsn5 and 70-15 (Fig. 6L). These results suggested that the increased MoAtg6 content in ΔMocsn5 was caused by decreased degradation rather than increased expression. Different types of anti-ubiquitin antibodies were used in western blotting assays, and we ultimately detected a reduced K48-ubiquitination level of MoAtg6 in ΔMocsn5 (Fig. 6I). Measurements of the ubiquitination and K48-ubiquitination levels of MoAtg6 in 70-15, ΔMocsn5, and Mocsn5-C reconfirmed that the knockout of MoCSN5 decreased the K48-ubiquitination level of MoAtg6 (Fig. 6J), which means that MoCsn5 promotes the K48-ubiquitination of MoAtg6 in M. oryzae.
Previous studies have shown that the K48-ubiquitination of Beclin1 (a homologous protein of Atg6) is a negative pathway for autophagy regulation [50, 51]. K48-ubiquitination promotes proteasome-dependent Beclin1 degradation, thereby effectively downregulating autophagy. Our research indicated that MoCsn5 promotes the K48-ubiquitination of MoAtg6, which promotes MoAtg6 degradation and thus downregulates autophagy in rice blast fungus.
MoCsn5 inhibits autophagy by suppressing the K48-ubiquitination of MoTorAutophagosome formation is regulated by the TORC1 complex, and active TOR signaling inhibits autophagy through the phosphorylation of Atg proteins [52, 53]. To test whether MoCsn5 regulates MoTor activity, we used rapamycin to validate the relationship between MoCsn5 and MoTor kinase. Compared with 70-15 and the complemented strain Mocsn5-C, the ΔMocsn5 mutant was more sensitive to rapamycin on CM at 25 °C for 9 days (Fig. 7A and B). This finding suggested that the activity of the TORC1 complex in ΔMocsn5 was altered. To verify the regulatory effect of MoCsn5 on MoTor activity, we examined the phosphorylation of the TORC1 activity marker MoRps6. Compared with those in the 70-15 strain, the phosphorylation levels of MoRps6 in the ΔMocsn5 mutant strain were significantly lower after treatment with rapamycin, indicating reduced TORC1 complex activity (Fig. 7C).
Fig. 7
MoCsn5 inhibits autophagy by suppressing the K48-ubiquitination of MoTor. A and B Colony morphology and relative growth rate of 70-15, ΔMocsn5 and Mocsn5-C on CM supplemented with 100 ng/mL rapamycin for 9 days. *p < 0.05. C The phosphorylation levels of MoRps6 in the 70-15 and ΔMocsn5 strains were detected by anti-phospho-Rps6 and anti-Rps6 antibodies. The phosphorylation levels of MoRps6 were calculated as phospho-MoRps6/MoRps6. The protein content of actin was used as a control. D Protein levels of MoTor in 70-15, ΔMocsn5 and Mocsn5-C. The strains were cultured in liquid CM at 25 ℃ for 36 h. The protein content of actin was used as a control. Gene knockout and complementation of MoCSN5 were performed in 70-15 expressing MoTor-Flag to ensure consistent transcript levels of MoTor. E Transcript levels of MoTor in 70-15 and ΔMocsn5. The error bars represent the standard deviations. F Ubiquitination levels of MoTor in 70-15, ΔMocsn5 and Mocsn5-C. After lysis, the strains were immunoprecipitated with anti-Flag beads, and a western blot analysis was subsequently performed with an anti-Flag antibody and an anti-ubiquitin antibody. The level of ubiquitinated MoTor was calculated as ubiquitin/MoTor-Flag. G K48-ubiquitination levels of MoTor in 70-15, ΔMocsn5 and Mocsn5-C. After lysis, the strains were immunoprecipitated with anti-Flag beads, and a western blot analysis was subsequently performed with an anti-Flag antibody and an anti-K48-ubiquitin antibody. The K48-ubiquitination levels of MoTor were calculated as K48-ubiquitin/MoTor-Flag
Our results showed that the reduction in the MoTor content in the ΔMocsn5 mutant induced a decrease in TORC1 complex activity (Fig. 7D), which in turn led to increased autophagy. However, no significant difference in the mRNA levels of MoTOR were found between the ΔMocsn5 and 70-15 strains (Fig. 7E). These results suggest that the decrease in the MoTor content in ΔMocsn5 is due to enhanced degradation rather than decreased expression.
Previous studies have shown that the protein content of Tor is regulated by ubiquitination [54]. The ubiquitination level of MoTor in ΔMocsn5 was significantly greater than that in the WT 70-15 (Fig. 7F), suggesting that MoCsn5-mediated ubiquitination regulates the protein level of MoTor. Measurements of the ubiquitination and K48-ubiquitination levels of MoTor in 70-15, ΔMocsn5, and the complemented strain Mocsn5-C also confirmed that the knockout of MoCSN5 increased the K48-ubiquitination level of MoTor (Fig. 7G). These results showed that MoCsn5 stabilized the MoTor content through specific K48-ubiquitination, thereby inhibiting autophagy.
留言 (0)