
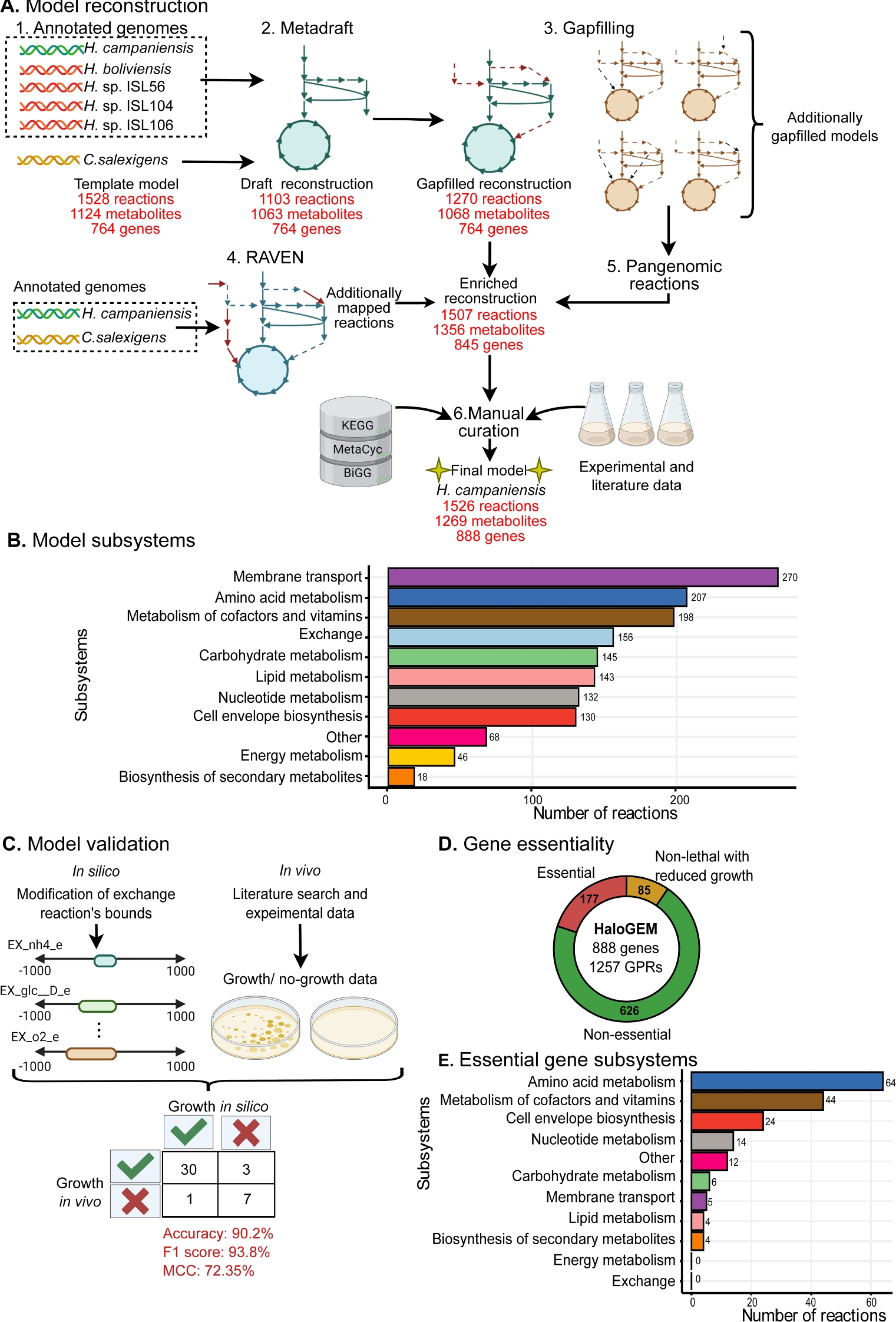
記住我



The success of many synthetic biology and biotechnology applications depends on the presence of a robust microbial chassis. We wanted to test if and how the presence of OLs in E. coli would affect its stress resistance and its performance as a host for synthetic biology applications.
First, we constructed a plasmid-borne synthetic operon containing the genes olsF and olsC required for the synthesis of unmodified OLs (uOLs) and hydroxylated OLs (OLs-OH) (Fig. 1a, b). From now on, we will use the abbreviation OLs when referring to both unmodified ornithine lipids (uOLs) and hydroxylated ornithine lipids (OLs-OH) (Rojas-Jiménez et al. 2005; Escobedo-Hinojosa et al. 2015). This plasmid called polsFC and the empty control plasmid pSEVA631 (pSEVA) (Silva-Rocha et al. 2013) were transformed into four different E. coli strains: the wild-type strain E. coli K12 (BW25113), the triple mutants PFC (phoB, fhlC, and cueR) and PYC (phoB, yedW, cusR), which have a reduced cellular proteome (Lastiri-Pancardo et al. 2020), and a phoB-deficient strain (Baba et al. 2006). Second, we wanted to determine if the E. coli strains expressing the synthetic operon formed uOLs and OLs-OH. The strains were grown in MOPS medium without phosphate limitation, [14C] acetate was added for lipid labeling, and total lipids were extracted and analyzed by thin-layer chromatography (TLC). In all strains, the glycerophospholipids PE and lipids PG and CL (which comigrate as a single band) were detected by TLC (Fig. 1c). In addition to these lipids, the E. coli strains expressing the synthetic operon olsFC produced both uOLs and OLs-OH (Fig. 1a–c). A few lipids of minor abundance that were not identified were observed in the TLC (labeled with asterisks). Interestingly, in E. coli strains accumulating uOLs and OLs-OH, the formation of PE was reduced relative to the control strains with the empty plasmid (Fig. 1c).
OL (uOLs and OLs-OH) production increases biomass yield under phosphate limited growth conditionsIn bacteria such as S. proteamaculans or V. cholerae, OLs are not formed under phosphate-replete conditions, and their synthesis is induced by phosphate-limited growth conditions (Barbosa et al. 2018). It is thought that the zwitterionic OLs replace the zwitterionic phospholipid PE under conditions of phosphate limitation, reducing the requirement for phosphate for the membrane and thereby allowing an alternative use for this nutrient. We observed a decrease in PE formation when uOLs and OLs-OH were accumulated in E. coli strains expressing the synthetic operon (Fig. 1c). Therefore, we hypothesized that an E. coli strain able to replace a fraction of its phospholipids with OLs could have a growth advantage under phosphate-limited conditions because phosphate normally used for membrane synthesis would be liberated for other cellular processes.
We performed growth experiments of all E. coli strains in MOPS/glucose medium with phosphate-limited concentrations (0.4 mM) and phosphate-replete concentrations (2.6 mM). When using 0.4 mM phosphate, we observed a substantial reduction in biomass yield, whereas the growth rates of the evaluated strains were not severely affected (Fig. S1a and b right). In this set of experiments, we included host strains and their derivatives carrying empty plasmids or expressing the synthetic operon olsFC. We determined the effect of the presence of OLs on the biomass production of E. coli grown under phosphate-limited (0.4 mM) and phosphate-replete (2.6 mM) conditions. In MOPS/glucose medium with 2.6 mM phosphate, no difference in biomass formation was observed between the different strains analyzed (Fig. S1a). In contrast, in phosphate-limited MOPS/glucose medium, biomass production in the strains that synthesized OLs increased by 23 to 41% compared with the strains carrying the empty plasmid (Fig. 2a). In all cases, the biomass formation of the OL-forming strains was higher than that of the wild-type strains lacking the plasmid polsFC (Fig. 2a, S1a and S2). These results suggest that the synthesis of OLs provides E. coli with an advantage when growing under growth-limited phosphate concentrations.
Fig. 2
The presence of ornithine lipids (uOLs and OLs-OH) in E. coli strains improves their growth phenotype under phosphate limitation and at pH 5.8. a Dry weight concentration (mgDW/mL) of cultures after 24-h growth in MOPS/glucose medium with low phosphate concentration (0.4 mM). b Growth rates of the different E. coli strains were calculated from the growth curves in MES/glucose medium with 2.6 mM phosphate at pH 5.8. The data represent the average of three independent experiments. Error bars indicate the standard deviation using two-tailed unpaired Student’s t-test. p values *p ≤ 0.05 and ****p ≤ 0.0001
OL (uOLs and OLs-OH) production improves growth phenotype at low pHWe wondered if the presence of OLs (uOLs and OLs-OH) would improve growth in MES/glucose medium without phosphate limitation at pH 5.8 in our engineered E. coli strains. As above experiments, we evaluated the four host strains and their derivatives carrying either the pSEVA631olsFC plasmid or the empty vector. The presence of OLs significantly enhanced the maximum growth rate of E. coli strains at pH 5.8, resulting in an approximately 35% increase compared with the vector control strains in three out of the four genetic backgrounds examined (Fig. 2b). When strains were grown under phosphate-replete conditions, no differences in growth were observed (Fig. S1a and S1b). On the other hand, during the first 8 h of the growth kinetic, all strains (with or without OLs) exhibited similar biomass. However, a noticeable difference emerged when comparing the plasmid-harboring strains in the subsequent hours, and after 24 h, all four OLs-OH-forming strains accumulated more biomass than the vector control strains. The OL-forming strains showed a shortened lag phase compared with the four wild-type strains lacking plasmids (Fig. S3).
Transcriptional response to OLs production at pH 7.4 and pH 5.8Abiotic stresses, including acidic pH, have been reported to affect the transcriptional response in E. coli (Aquino et al. 2017). Therefore, we wanted to determine if the positive phenotypic effects at low pH caused by the presence of OLs in E. coli could be explained by transcriptional changes. Given that PYC and PFC strains lack transcription factor genes that rewire their transcriptional regulatory network (Lastiri-Pancardo et al. 2020), we used the wild-type K12 strain to evaluate the specific effect of the presence of OLs on the transcriptome of E. coli grown at both pH 7.4 and pH 5.8 without phosphate limitation.
Total RNA was extracted from exponentially growing cultures (11 h) as, at this time point, a clear difference in growth rate was observed between the strains forming OLs and those lacking them (Fig. 2a). We conducted RNA-Seq analysis on two independent biological samples and sequenced the libraries to approximately 10 million read depths. We then compared the expression profiles of these samples based on their response to different perturbations: (A) response to OL production at neutral pH, (B) response to OL production at low pH, (C) response of the control strain to low pH compared with neutral pH, (D) response of the OL-producing strain to low pH compared with neutral pH, and (E) differential response to low pH and OL formation (Table 1).
Table 1 Transcriptional response to OL (uOLs and OLs-OH) formation at neutral and low pH. Perturbations and descriptionsFor the transcriptomic analyses, we used the iModulon classification of the differentially expressed genes (DEGs) data, which consists of the clustering of genes sharing an independently modulated signal. iModulons were obtained by independent component analysis (ICA) of 815 sample transcriptional profiles from 422 conditions for E. coli. For further analyses, we considered only those iModulons that covered 80% or more of the DEGs with a p value < 0.05 and log2 fold change of ± 1; that is, from the 204 classified iModulons (Rychel et al. 2021), 48 passed the filter (Fig. 3a and Fig. S4). Overall, the results show a mild transcriptional response to OL production (perturbations A and B) and a large transcriptional response to low pH conditions (C and D) (Fig. 3b).
Fig. 3
Low pH conditions cause a large transcriptional response, whereas OLs formation causes a mild transcriptional response. a 48 iModulons with coverage greater than 80% in at least one perturbation were used for this analysis. The functional categories are denoted according to color, yellow: metabolism, black: stress response, white: other functions. b The percent of genes that are over ( +) or under ( −) expressed in each iModulon. c Log2 fold change of individual genes involved in arginine metabolism, membrane and lipid synthesis, and stress response to low pH not classified by iModulons (value = DEGup − DEGdown); blue color, reduced expression, and red color, induced expression
The presence of OLs in E. coli grown at pH 7.4 (perturbation A) affected the gene expression level of 25 iModulons, 13 of which were upregulated, including those related to stress response (Curli, PspF, Hydrogenase 1, ProVWX, BlurR, asr, ZntR, and Fur 1–2) and metabolism (GlcC, BirA, AHAS I, and PurR). Some of the iModulons with reduced expression in perturbation A are associated with the maintenance of membrane and proton motive force (PMF) (pspF) and the integrity of the outer membrane under moderate acid stress (ars) (Fig. 3b). In perturbation B (presence of OLs at pH 5.8), five and eight iModulons showed reduced and increased expression, respectively. Of note, the expression of genes clustered in the pspF iModulon was also decreased as in perturbation A (Fig. 3b). For perturbations C and D (effect of pH 5.8 in the absence and presence of OLs, respectively), a similar tendency was observed in the class of iModulons affected (stress and metabolism). However, in perturbation D, the nutrition limitation-associated pyrR iModulon was upregulated while it did not change in perturbation C (Fig. 3b).
To better understand the specific transcriptional responses that benefit the growth phenotype of the OL-producing strains under low pH, we compared the expression profile of the control strain at neutral pH with that of the OL-producing strain at low pH (perturbation E). Two of the 48 filtered iModulons showed changes in their activity in this perturbation. The differential response to low pH in the presence of OLs showed a few key differences compared with the response of the control strain at neutral pH (Fig. 3b). For instance, reduced expression of the pspF iModulon was detected in perturbation E (as in A and B), suggesting that the presence of OLs specifically reduces the expression of genes related to the maintenance of PMF. In addition, expression of the kdpE iModulon (potassium two-component system) was reduced in the presence of OLs (perturbations, B, C, D, and E). In particular, a reduction in the expression of the pdhR iModulon, which contains genes associated with the formation of the pyruvate dehydrogenase (PDH) multienzyme complex and respiratory electron transport system (Ogasawara et al. 2007), was only detected by the analysis of perturbation E (Fig. 3b).
An increase in the activity of the curli iModulon in perturbations A, B, and E indicates that OL production positively affects the expression of genes related to curli fiber synthesis and biofilm formation in E. coli. In addition, the proVWX iModulon (transport system for the osmoprotectant glycine betaine) was also induced in these perturbations (Fig. 3b).
Besides the above-mentioned analyses by iModulons, we also analyzed individual genes that are related to ornithine synthesis, the synthesis and turnover of membrane lipids, and the response to acid stress in the four defined perturbations (Fig. 3c). Genes adiY and adiC, which encode an arginine/agmatine antiporter of the arginine-dependent extreme acid resistance (XAR), were induced at low pH (Fig. 3c; perturbations C, D, and E). Genes involved in the acid stress response such as gadA, hdeD, and yhiM were induced by the formation of OLs (perturbations A, B, and E). In contrast, the expression of asr that encodes a periplasmic intrinsically disordered protein induced under acid shock conditions was downregulated in perturbations A, C, D, and E (Fig. 3c). Collectively, these results showed that modifications of the membrane composition by OLs change the regulatory response to low pH in E. coli.
OL-producing strains are more tolerant to the proton motive force uncoupler CCCPThe consistently reduced expression of the pspF iModulon in strains producing OLs (Fig. 3; perturbations A, B, and E) suggests that changes in membrane lipid composition could affect how engineered strains manage proton membrane permeability, potentially providing a mechanism for their increased tolerance to low pH. This iModulon comprises genes known as Phage Shock Proteins, which are membrane proteins that play a critical role in regulating the membrane permeability to protons. Recent research by Wang et al. (2021) indicated that pspA mutants have a decreased ability to maintain the PMF during periods of starvation as well as its participation in long-term stationary phase survival was shown (Takano et al. 2023). To investigate this, we determined the effect of the presence of OLs on the biomass production of E. coli growing in the presence of 10 µM of the ionophore carbonyl cyanide m-chlorophenyl hydrazine (CCCP), a compound that dissipates the proton gradient across the inner membrane (Kane et al. 2018). The strain producing OLs exhibited better growth than the E. coli K12 strain carrying the empty vector (Fig. 4a). Furthermore, the OL-producing strain showed enhanced tolerance to CCCP, as it grew 35% faster than the control strain (Fig. 4b). These results indicate that the presence of OLs in the E. coli membranes makes the cell membrane less proton permeable, providing a possible mechanism for the low pH resistance of the engineered strains evaluated in this work.
Fig. 4
The presence of OLs in E. coli increases resistance to the ionophore CCCP (10 µM). E. coli strains harboring the empty control plasmid or the plasmid containing olsFC were grown at pH 7.4 in MOPS/glucose medium without phosphate limitation. The growth curves are shown in (a), and the maximum specific growth rates are shown in (b). The data represent the average of three independent experiments. Error bars indicate the standard deviation using two-tailed unpaired Student’s t-test. p values *p ≤ 0.05
OL production (uOLs and OLs-OH) increases heterologous violacein productionWe have shown that the presence of OLs in E. coli benefits its growth under both phosphate-limited and acidic pH growth conditions. These improved properties could enhance the performance of E. coli strains during heterologous expression, for example, during industrial applications. Previously, we reported that the proteome-reduced E. coli PFC strain produced higher levels of violacein than the wild-type strain, a pigment of industrial interest naturally synthesized by Chromobacterium violaceum (Lastiri-Pancardo et al. 2020). Therefore, and as a proof-of-concept experiment, we tested the ability of OL-producing engineered strains to synthesize violacein. The biosynthetic operon vioABCDE was cloned into the plasmid pAJM336 under the control of an IPTG-inducible promoter. The resulting plasmid pAJM336:vioABCDE (Fig. 5a) was transformed into OL-producing derivatives of the proteome-reduced mutants PFC and PYC. First, we determined that these strains transformed with polsFC produced OLs (Fig. S5). Then, we quantified violacein production in the evaluated strains after 24 h at 30 °C in MOPS/glucose medium (pH 7.4) in phosphate-limited conditions (0.4 mM phosphate) and MES/glucose medium (pH 5.8) under phosphate-replete conditions (2.6 mM phosphate). At pH 7.4 and phosphate-limited conditions in the absence of OLs, the triple mutant PFC accumulated more violacein than the triple mutant PYC (Fig. 5b). Both strains, PFC and PYC, produced more than twice the violacein levels under phosphate limitation when synthesizing OLs than the control strains (Fig. 5b). At pH 5.8 with 2.6 mM phosphate, again comparing the strains harboring the empty plasmid with the strains expressing olsFC, violacein production was more than tenfold higher in strains forming OLs (Fig. 5c). These results demonstrate that our membrane engineering approach is beneficial for producing costly metabolites via a heterologous pathway in a metabolic engineering application.
Fig. 5
E. coli strains forming OLs (uOLs and OLs-OH) show an increase in total violacein production. All strains harbored the plasmid pJAM336:vioABCDE (a) in addition to pSEVA or polsFC. Strains were grown in MOPS/glucose medium at pH 7.4 with 0.4 mM phosphate (b) and in MES/glucose medium at pH 5.8 with 2.6 mM phosphate (c), supplemented with 2 g/L tryptophan, 800 µM IPTG, and 0.01% casamino acids (mean + / − s.d., n = 9)
留言 (0)