
記住我
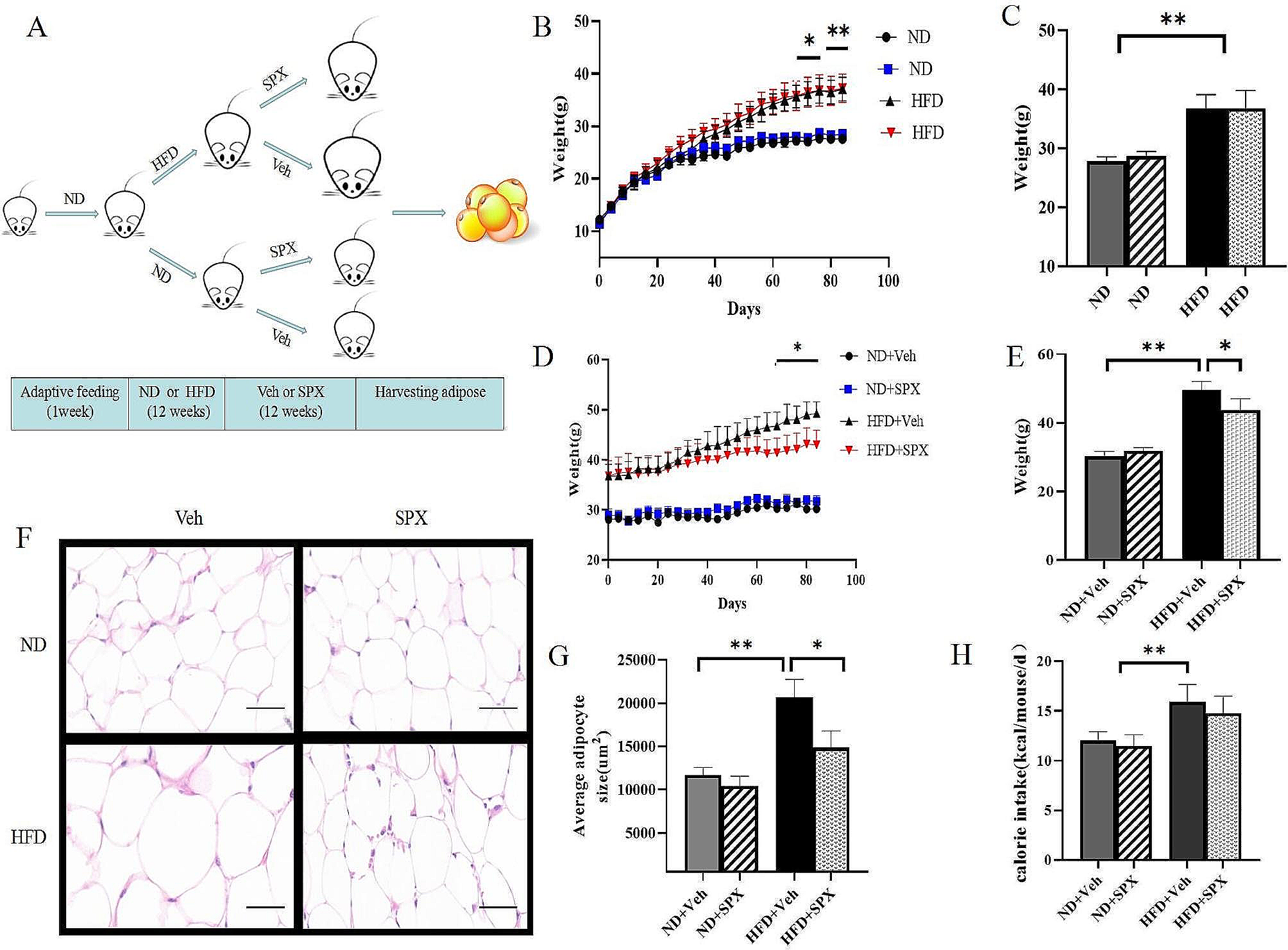
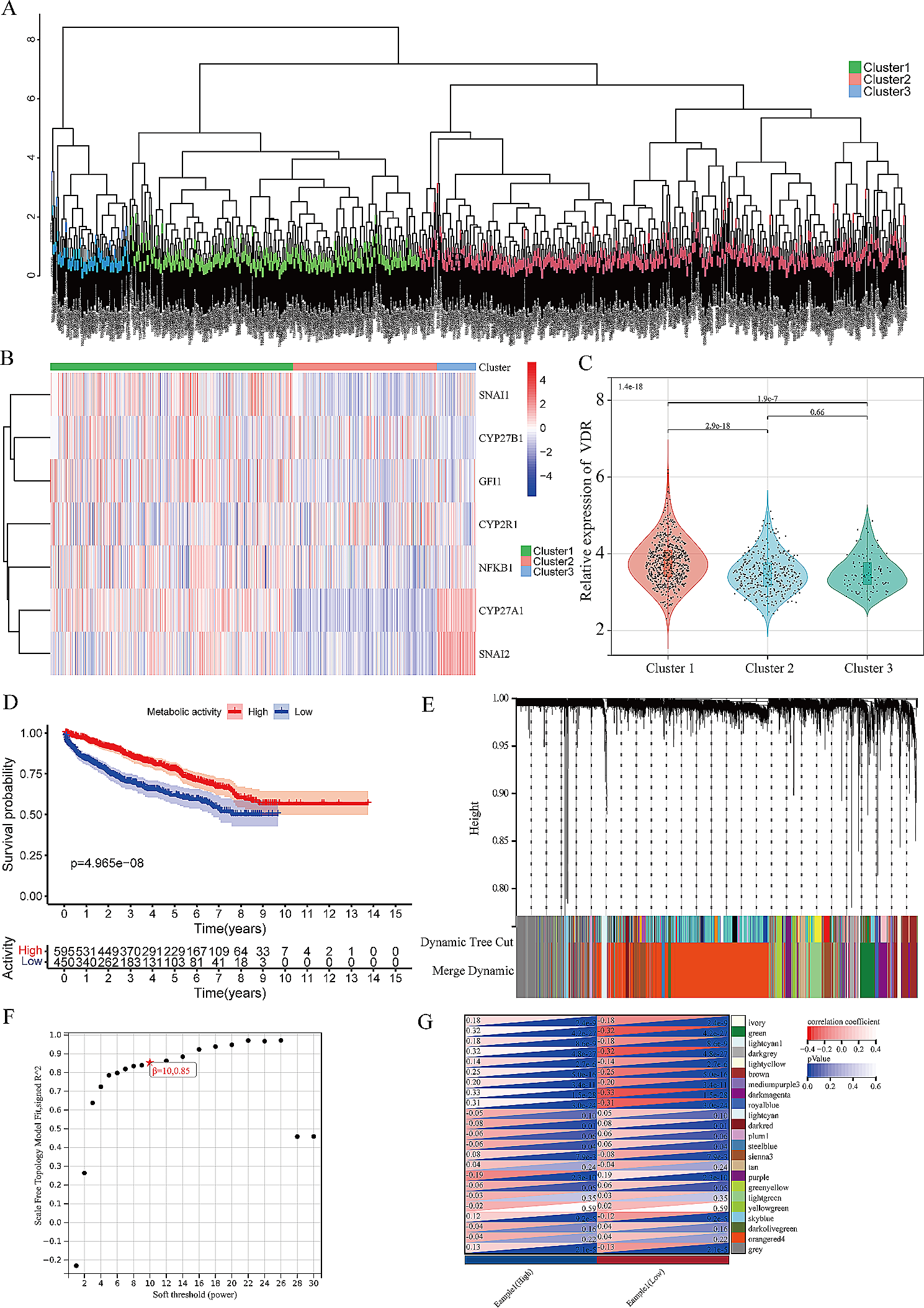


Age-related loss of muscle mass contributes significantly to frailty and mortality risk. We therefore firstly tested whether DFC could attenuate weight loss in aged mice with 8 weeks of dietary intervention. There was no significant change in body weight for either YG or AG groups in the first week following DFC intervention (p > 0.05) (Table 2). However, by week 8, the AG-NDF group showed decreased body weight compared to baseline (− 1.60 ± 1.56 g, p < 0.05), indicating age-related weight loss. In contrast, DFC supplementation attenuated this weight loss in a dose-dependent manner, with both AG-LDF (+ 3.10 ± 2.45 g, p < 0.05) and AG-HDF (+ 5.30 ± 2.04 g, p < 0.05) groups showing significantly increased body weight compared to AG-NDF controls. The body weight gains in AG-HDF mice were also significantly higher compared to AG-LDF (p < 0.05).
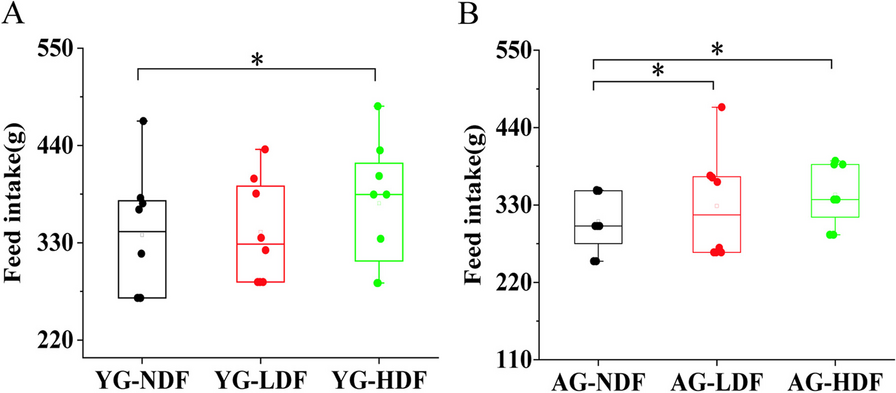
Table 2 Changes in body weight of mice in each group after 8 weeks of interventionIn young control mice (YG groups), DFC decreased body weight gain relative to YG-NDF controls, with enhancement more pronounced in YG-HDF (− 2.20 ± 1.84 g) versus YG-LDF (− 3.60 ± 2.38 g) groups (p < 0.05). Food intake followed a similar improvement with DFC in both aged and young groups (Fig. 1A, B), indicating food utilization efficiency was increased with DFC supplementation, particularly at higher doses. Overall, these data demonstrate that DFC attenuates age-associated weight loss and improves food utilization in mice.
Fig. 1
The changes in food intake of mice in each group after 8 weeks of intervention. A Food intake in each YG subgroup of mice. B Food intake in each AG subgroup of mice. Note *P < 0.05 **P < 0.01 vs NDF group
The impacts of DFC on organ coefficients in aged versus young miceTo assess changes in relative organ mass, organ coefficients (the ratio of organ weight to overall body weight) were determined.. The values of organ coefficients of mice after 8 weeks of DFC intervention were calculated and are shown in Table 3. Compared to YG-NDF controls, the organ coefficients of all examined organs were significantly decreased (p < 0.05) in AG-NDF mice. In the YG group, compared with the YG-NDF controls, the liver coefficient decreased significantly in the low-dose and high-dose DFC groups (p < 0.05), but there were no significant changes in other organ coefficients (p > 0.05). In the AG groups, heart, liver, spleen, and lung coefficients exhibited significant increases (p < 0.05) with both low and high-dose DFC versus AG-NDF controls. Kidney coefficients did not significantly differ between any AG subgroups (p > 0.05). These results demonstrate that organ decline occurred in aged mice to a certain extent, and DFC supplementation could increase the organ coefficient of aged mice, and the organ-protective effect of DFC appear somewhat more pronounced in higher doses of AG mice.
Table 3 Effect of DFC intervention on organ coefficients of mice in each groupEffects of DFC on antioxidant capacity in aged versus young mouseAs oxidative stress plays a key role in the aging process, we hypothesized that DFC intervention might help suppress oxidative stress through increasing antioxidant capacity. Therefore, we examined the effects of DFC on antioxidant capacity in the livers and serum of young versus aged mice by measuring total antioxidant capacity (T-AOC), superoxide dismutase (T-SOD), and glutathione peroxidase (GSH-Px) that reflect antioxidant capacity, as well as malondialdehyde (MDA), a biomarker of oxidative stress (Fig. 2) [22, 23].
Fig. 2
Effects of DFC intervention on antioxidant capacity in liver and serum of mice. Antioxidant capacity was assessed by measuring (A) T-AOC activity. (B) T-SOD activity. (C) MDA levels. (D) GSH-Px levels. Note *P < 0.05 **P < 0.01 versus NDF group
Compared to YG-NDF controls, T-AOC activities significantly increased in both the liver (76.51%) and serum (45.10%) of YG-HDF mice (p < 0.01). T-SOD activity also increased by 44.91% in YG-HDF serum (p < 0.01). MDA decreased by 77.90% in YG-HDF liver (p < 0.01). Liver GSH-Px increased by 63.26% in YG-HDF (p < 0.01). In AG mice, liver and serum T-AOC showed large significant increases of 95.64% and 115.05% in AG-HDF, respectively (p < 0.01). Liver T-SOD increased by 8.71% in AG-HDF (p < 0.05). Serum T-SOD increased by 77.55% in the AG-HDF (p < 0.01). MDA decreased by 70.76% in AG-HDF liver (p < 0.01) and 12.00% in serum (p < 0.05). Liver GSH-Px activity increased by 60.58% with AG-HDF (p < 0.01), but serum GSH-Px activity had no significant difference (p > 0.05). Collectively, DFC supplementation substantially improved antioxidant capacity in both young and aged mice, especially at high doses. The liver and serum of aged mice showed greater enhancements in T-AOC with DFC compared to young controls.
DFC supplementation shifted Characteristic bacterial genera compositionAs the gut microbiota plays an important role in health and can be impacted by diet and age, we evaluated the effects of 8 weeks of DFC supplementation on key gut bacterial groups in feces of YG and AG mice (Fig. 3). As we all know, when the amount of E. coli in the intestine is excessive, it will destroy the intestinal barrier and become an opportunistic pathogen [24]. Compared to NDF controls, the E. coli decreased with DFC treatment in both YG and AG groups. In AG mice, E. coli was significantly reduced by 20.84% in the LDF group (p < 0.05) and 45.62% in the HDF group (p < 0.01). In YG mice, it was significantly reduced by 38.66% (p < 0.05) in the LDF group and 63.64% (p < 0.01) in the HDF group. In the contrary, the beneficial genus Lactobacillus was significantly elevated by 128.46% (p < 0.01) in the YG-LDF group and 449.74% (p < 0.01) in the YG-HDF group. It was elevated in the AG groups as well by 79.31% (p < 0.05) in the AG-LDF group and 329.73% (p < 0.01) in the AG-HDF group. Another beneficial gut microbe Bifidobacteria was also elevated in both YG and AG groups upon DFC intervention, which was elevated by 109.05% (p < 0.05) in the YG-LDF group, 341.79% (p < 0.01) in the YG-HDF group, 66.34% (p < 0.05) in the AG-LDF group and 239.75% (p < 0.01) in the AG-HDF group. While being generally considered a beneficial genus of gut bacteria, however, the level of Bacteroides was significantly reduced by 39.22% (p < 0.01) in the YG-LDF group, 45.62% (p < 0.01) in the YG-HDF group, 52.00% (p < 0.05) in the AG-LDF group and 63.42% (p < 0.01) in the AG-HDF. It is known that some species of Bacteroides can become opportunistic pathogens if they spread beyond the gut and cause systemic infection [25]. Therefore, these results indicate that DFC supplementation, significantly increased beneficial bacteria and reduced potential pathogens in both young and aged mice, suggesting DFC may improve characteristic bacterial genera composition in aging.
Fig. 3
DFC supplementation alters characteristic bacterial genera composition in young and aged mice. Relative abundance of key gut bacterial genera was determined in the feces of young (YG) and aged (AG) mice after 8 weeks of DFC supplementation. (A) E. coli. (B) Lactobacillus. (C) Bifidobacterium (D) Bacteroides. Data shown as median and SD relative to respective NDF groups, analyzed by Student T-test. Note *P < 0.05 **P < 0.01 vs NDF group
Identification of metabolite changes in mouse feces upon DFC interventionTo investigate the metabolic alterations induced by DFC supplementation, we performed untargeted 1H NMR-based metabolomics analysis on fecal samples from mice after 8 weeks of DFC treatment. A representative metabolite 1H NMR spectrum in feces was shown in Fig. 4. A total of 46 metabolites were identified in mouse fecal samples, with chemical shifts mainly distributed in the range of 0.5–8.5 ppm, spanning diverse chemical classes including sugars, amino acids, fatty acids, organic acids, and others. Assignment of metabolites was confirmed using the human metabolome database (HMDB) (http://www.hmdb.ca/) and biological magnetic resonance bank (BMRB) (http://www.bmrb.wisc.edu/), which were calibrated and assigned in conjunction with references [26,27,28], and are summarized in Additional file 3: Table S1. The 1H signal in the 0.8–3.0 ppm region represents amino acids (leucine, valine, alanine, aspartic acid, proline, and lysine, etc.), short chain fatty acids (acetic acid, propionic acid, and butyric acid, etc.), organic carboxylic acids (bile acid, lactic acid, succinic acid, and citric acid, etc.), and other metabolites (dimethylamine, trimethylamine, and N-acetylglycoprotein, etc.). The 1H peak in the 3.0–5.5 ppm range mainly includes sugars (α-Glucose, β-Glucose, α-Galactose, and β-Galactose), amino acids (such as histidine, lysine, threonine, glycine, and serine), and other metabolites (such as choline and methanol). Finally, amino acids (tyrosine), organic acids (formic acid, fumarate), and other metabolites (inosine, uric acid, uracil, xanthine, and hypoxanthine, etc.) were found in the region of around 5.5–8.5 ppm.
Fig. 4
Representative 1H NMR metabolic spectrum of metabolites in mouse fecal samples
To characterize the overall metabolic profile in the feces of each mouse group, PCA was performed on the 1H NMR spectral data. As shown in Fig. 5, The PCA scores plot showed clustering of the samples according to the treatment group, with separation along the first principal component (PC1, 47%) distinguishing the young control, aging model, and DFC-treated aging groups. The second principal component (PC2, 11.6%) revealed the metabolic separation characteristics between the aging model group fed basal diet versus the low and high-dose DFC groups.
Fig. 5
Scatter plot of PCA scores of fecal metabolites in mice after DFC intervention. PCA reveals dose-dependent and age-dependent changes in the fecal metabolomic profiles of mice in response to 8 weeks of dietary DFC intervention. Note AN = aging mice group on a no dietary fiber feed (aging control); AL = aging mice group on a low-dose DFC feed; AH = aging mice group on a high-dose DFC feed; YN = young mice group on a no dietary fiber feed group (negative control)
Altered fecal metabolites in mice following DFC interventionWhile PCA demonstrated clear metabolic changes between different groups, there was some overlap in the PCA scores clusters. To better discriminate the metabolic variations between groups, OPLS-DA was applied. As shown in Additional file 3: Fig. S1, the R2Y and Q2 values were greater than 0.9 for all pair-wise comparisons between groups: R2Y = 0.994 and Q2 = 0.991 between the AG and YG group; R2Y = 0.994 and Q2 = 0.987 between the LDF and NDF group; R2Y = 0.994 and Q2 = 0.987 between the HDF and NDF group. When these values are greater than 0.5, it is considered a high degree of fitting. The closer the values to 1, the more reliable the OPLS-DA model. Therefore, the above results indicate the OPLS-DA models have excellent predictive ability and can effectively explain the significant differences in fecal metabolites among different mouse groups.
A permutation test was conducted under 200 conditions for further interactive validation, which confirmed the robustness of the OPLS-DA models (as shown in Additional file 3: Fig. S1). Based on the log2Fold Change (log2FC) value combined with variable importance in projection (VIP) > 1 and p < 0.05, differential metabolites and up-regulation ranges were identified (Table 4). Between AG and YG, 9 metabolites were altered including increased uric acid, threonine, inosine, fumarate, leucine, pyruvate, and aspartic acid, and decreased propionic acid and butyric acid in AG. Between LDF and NDF, 12 metabolites changed significantly with propionic acid, butyrate, proline, glutamate, serine, fumarate, and citric acid were elevated in LDF while uridylic acid, trimethylamine, threonine, aspartate, and inosine decreased. Finally, 11 metabolites differed between the HDF and NDF groups, with lactate, butyric acid, succinic acid, and propionic acid upregulated and uric acid, pyruvate, threonine, valine, β-glucose, alanine, and aspartic acid downregulated in HDF.
Table 4 Comparison of Fecal Metabolite changes in AG and YG MiceIn summary, the OPLS-DA models identified distinct metabolite signatures differentiating the aging, DFC-treated, and young control groups, providing insight into the metabolic pathways altered by aging and DFC supplementation.
Metabolic pathway analysis of age-related changes and the impact of DFC intake on fecal metabolitesTo gain further insights into the metabolic perturbations underlying the observed metabolite changes with DFC treatment and aging, metabolic pathway analysis was performed on potential differential metabolites (Fig. 6). Between AG and YG, the altered metabolites were enriched in pyruvate metabolism and alanine, aspartate, and glutamate metabolism pathways. Between LDF and NDF, 5 pathways were significantly affected including alanine, aspartic acid and glutamate metabolism, arginine biosynthesis, tricarboxylic acid cycle (TCA), arginine, and proline metabolism, as well as D-glutamine and D-glutamate metabolic pathways. Between HDF and NDF, enriched pathways included pyruvate metabolism, glycolysis/gluconeogenesis, and alanine, aspartate, and glutamate metabolism. The common enriched pathways across the comparisons highlight dysregulation of energy metabolism (TCA cycle, glycolysis/gluconeogenesis) as well as amino acid metabolism mediated by the microbiome (alanine, aspartate, glutamate pathways) with DFC treatment and aging. Specifically, the TCA cycle and glycolysis pathways suggest modulation of carbohydrate metabolism and cellular respiration. Changes in amino acid pathways indicate altered microbial fermentation and inter-conversion of amino acids. Taken together, the metabolic pathway analysis points to significant effects of DFC and aging on gut microbial activity related to carbohydrate and amino acid metabolism.
Fig. 6
Effect of DFC intake on key metabolic pathways altered in fecal metabolites of mice groups. A Metabolic pathways of differential metabolites between AG and YG groups. B Metabolic pathways of differential metabolites between LDF and NDF groups (C) Metabolic pathways of differential metabolites between HDF and NDF groups. Each bubble represents a distinct metabolic pathway. The Bubble color indicates the P-value, and the horizontal axis represents the enrichment factor. Note The larger the enrichment factor, the smaller the P-value, and the more significant the enrichment degree. Altered metabolic pathways were identified by screening for those with P < 0.05 and enrichment factors > 0.1
Correlation between characteristic bacterial genera and major differential fecal metabolitesTo further elucidate the interactions between the characteristic bacterial genera and main differential metabolites, Spearman correlation heatmap and correlation network analysis were performed between the characteristic bacterial genera and the main differential metabolites in each intervention group (Fig. 7).
Fig. 7
Spearman correlation analysis between characteristic fecal microbiota and main differential metabolites. A Heatmap and network diagram showing correlations for the LDF group. B Heatmap and correlation network diagram showing correlations for the HDF group. The heatmaps display the Spearman correlation coefficient (R) values, with red indicating a positive correlation and blue indicating a negative correlation. Note The network diagrams visualize the correlations, with green lines representing positive correlation, gray lines representing negative correlations, and line thickness proportional to the correlation coefficient. Red circles denote bacteria genera and blue rectangles denote metabolites
In the LDF group, E. coli was positively correlated with inosine and negatively correlated with glutamate. Lactobacillus displayed positive correlations with propionic acid and butyric acid, and negative correlations with aspartic acid, inosine, trimethylamine, and uric acid. Bifidobacterium was positively associated with propionic acid and butyric acid, and negatively related to uric acid and trimethylamine.
In the HDF group, E. coli showed negative correlations with butyric acid, lactic acid, and propionic acid, and was also negatively correlated with alanine. Lactobacillus displayed positive correlations with butyric acid, lactic acid, and propionic acid, while showing negative correlations with threonine, alanine, and aspartic acid. Bifidobacterium was positively correlated with butyric acid, lactic acid, and propionic acid, and negatively correlated with alanine. Bacteroides exhibited positive correlations with β-glucose and pyruvate.
These association patterns suggest the increased Lactobacillus and Bifidobacterium with HDF may drive elevations in short-chain fatty acids through carbohydrate fermentation. The decreased E. coli may relieve some of the reductions seen in metabolites like alanine. The higher Bacteroides could contribute to increases in lactate, pyruvate, and glucose.
留言 (0)