
記住我





We, and others, have previously shown the utility of GSC—human CO co-culture models for investigating GBM biology. Here, we describe a novel model we call long-term glioma cerebral organoids (ltGLICOs)—in which new COs were generated from mixed cultures of H1-ESC containing low levels (< 1:1000) of 320-GSCs (Additional file 1: Fig. S1A). 320-GSCs were derived in-house from an IDH-wt mesenchymal subtype primary GBM patient surgical sample, and thoroughly characterized in our previous studies [15,16,17]. To monitor the tumorigenic process, ltGLICOs spanning 1–24 months in culture (n = 17) (Additional file 1: Fig. S1B), were profiled by scRNAseq and in silico detection of 320-GSCs was performed (see methods). This strategy was able to unambiguously genotype > 99% of all cells (Additional file 1: Fig. S1), with the remaining cells identified according to nearest-neighbor clustering (Additional file 1: Fig. S1D–E). UMAP projection showed clear separation of cells by their genotype (Fig. 1a), which was further corroborated by inferred copy number variation analysis (Fig. 1b). Intriguingly, although GSCs were detected in all ltGLICOs profiled (Fig. 1d and Additional file 1: S1F), growth did not appear to be linear with age, rather ltGLICOs had either very low levels of detectable GSCs or they had undergone near-total replacement of CO cells with 320-GSCs. We observed that 2/6 at 8–9 months, 4/6 samples at 12 months, and all samples beyond 18 months had undergone this change. To investigate the extent to which 320-GSCs disrupted normal CO development we aggregated data across all cells to test how ltGLICO pseudo-bulk transcriptomes compared to different fetal brain developmental stages recorded in the BrainSpan atlas (Fig. 1d). We saw clear evidence of progressive maturation, with ltGLICOs correlating with later developmental stages as they aged, reaching a peak of maturation between 8 and 12 months by which point they showed similarity with fetal brain from late gestational stages and early neonates. However, in those ltGLICOs that had undergone the oncogenic expansion of the 320-GSC population, this close correlation to developing brains was lost.
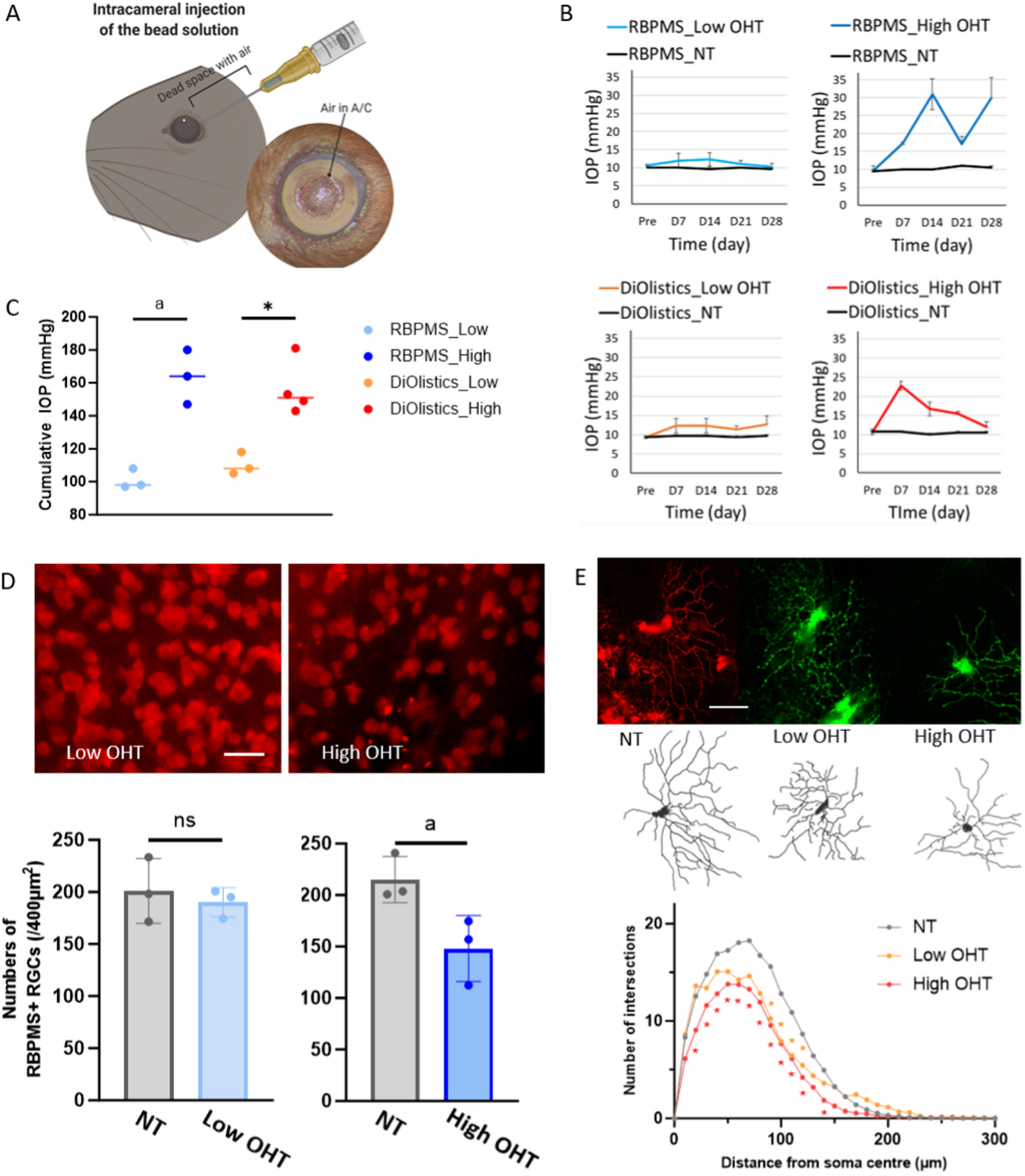
Fig. 1
In silcio genotyping reveals extended latency period before GSC expansion in longterm-GLICOs. a UMAPs of the ageing ltGLICO cohort (spanning 1–24 months) colored by age (left), and genotype (right). b Inferred copy number plot separates ltGLICO cells according to genotype. c Proportion of 320-GSCs cells per ltGLICO sample. d Mean Pearson correlations of ltGLICO pseudo-bulk transcriptomes and primary human fetal brain tissue from the BrainSpan dataset grouped by age (weeks post conception). e Proportion of H1-ESC and 320-GSC specific SNP fingerprints detected in controls and ltGLICO samples
In addition to transcriptomic analyses, we profiled 5 ltGLICOs spanning 2–24 months as well as H1-ESCs (from which the COs were derived) and 320-GSC controls by whole exome sequencing (WES) to determine the stage at which 320-GSC mutations become detectable in ltGLICO bulk DNA extracts. Mutually exclusive germline single nucleotide polymorphism (SNP) DNA ‘fingerprints’ were generated from comparison of H1-ESC and 320-GSC genomic profiles, and ltGLICO samples were assayed for the proportion of each genotype-specific DNA fingerprint that was detected (Fig. 1e). In concordance with the single-cell findings, some 320-GSC specific SNPs were detectable in all ltGLICOs profiled, and by 24 months H1-ESC DNA had been largely replaced by 320-GSCs DNA.
To accompany our sequencing analyses, we also performed a neuropathological assessment of ltGLICO sections across the aging cohort (Additional file 1: Fig. S2A). H&E-staining showed that young ltGLICOs were macroscopically normal, with clear embryonic neural rosettes that dissipated over time, glial cells that increase with time, choroid plexus epithelium and pigmented retinal epithelium as previously shown for COs in the literature. By 9 months, however, we observed the emergence of aberrant features in the central regions of ltGLICOs. By 18–24 months, ltGLICOs were entirely replaced by cells with frankly malignant histologic features, including cells with marked nuclear pleomorphism, cells with high nuclear-to-cytoplasmic ratios, and nuclear molding. The presence of spontaneous necrosis and frequent apoptotic bodies was consistent with high-grade histology, which is typical of CNS W.H.O. grade IV GBM (Additional file 1: Fig. S2B). Immunohistochemical staining for CD44, a marker of the mesenchymal GBM-subtype strongly expressed by 320-GSCs, showed increased expression as ltGLICOs aged. We also observed changes in the patterns of Ki67 staining, with cell proliferation largely restricted to neural rosettes of younger ltGLICOs, reduced in middle aged ltGLICOs, but then greatly increased in the oldest samples (Additional file 1: Fig. S2A). Taken together, our transcriptomic, genetic, and histological analyses of aging ltGLICOs reveal that minimally seeded 320-GSCs show a prolonged latency period prior to rapid expansion and replacement of organoid cells which occurs after approximately 8–12 months in culture. Based on these analyses we focused our attention on microenvironmental changes in the CO itself that occur over time and may help provide a permissive environment for GSC cell expansion.
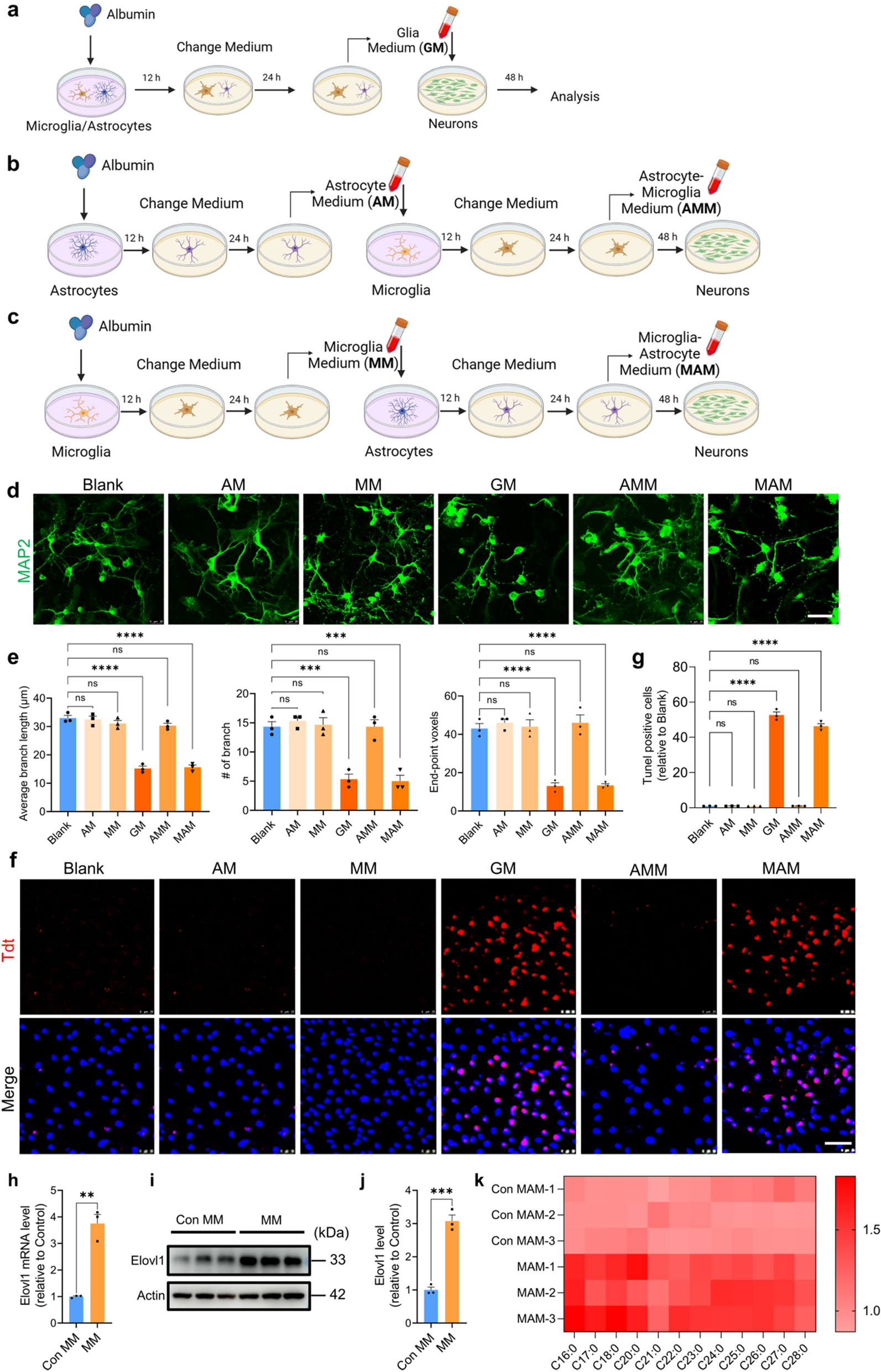
To better understand how the organoid component of the ltGLICOs developed over time, H1-derived cells were subset, new UMAPs were projected, and cell clusters were identified based on known marker genes (Fig. 2a–c). 1-month-old ltGLICOs harbored numerous neural rosettes (Fig. 2d) and were highly enriched for clusters expressing markers of proliferative radial glia (C.11, 4 & 2: MKI67, NES, PAX6, HES5). 2-month-old ltGLICOs were defined by a wave of neurogenesis with expansion in the pool of newborn excitatory neurons (C.1, 18 & 0: NEUROD2, DCX, CD24), and clusters expressing transcription factors and synaptic ionotropic receptor genes associated with more mature neurons (C.9 & 6: MEF2C, GRIN2A). By 4 months, ltGLICOs harbor a population of midbrain inhibitory interneuron progenitor cells (InN.NPCs), that specifically express distal-less homeobox family transcription factors (C5: DLX2, DLX5, DLX6-AS1). The initial neurogenesis was subsequently followed by a wave of gliogenesis with the first appearance of oligodendrocytes, and their precursors (C.10: OLIG1, DLL3, SIRT2) in 4-month-old ltGLICOs, concurrent with expansion of outer radial glia and astrocytes (C.7, 8: HOPX, GFAP, AQP4). This expansion of glial cell types was even more apparent in 8-month and ≥ 12-month-old ltGLICOs (Additional file 1: Fig. S3A), by which point a new population of astrocytes had emerged. This cluster maintained expression of astrocyte markers but also showed higher expression of genes associated with reactive gliosis and environmental stress (C.10; LGALS3, S100A10, SPP1, CD44). Given the concurrence of the emergence of this astrocyte population and the expansion of the 320-GSCs we scored cells for a gene signature derived from glioma-associated astrocytes [23], finding the greatest signature enrichment in this cluster (Fig. 2f). Other cell types detected included choroid plexus and meninges cells (C.14, 3, 7: TTR, IGFBP7, FOLR1 and C.20: COL1A1, MGP respectively) as well as three different retina cell types; retinal neural progenitors (C.21: SIX3, SIX6, HMX1), muller glial cells (C.16: HMX1, CRABP1) and rod cells (C.19: RCVRN, PDC). In line with previous studies [3], we also observed two clusters expressing genes associated with glycolysis (BNIP3, PGK1) and hypoxia (VEGFA, HILPDA), which corresponded to stressed neurons (C.13) and astrocytes (C.15) respectively, and increased over time as organoids grew in size. Neurodevelopment of primary fetal brain and cerebral organoids has now been thoroughly characterized and widely reported in the literature [3, 24,25,26,27,28,29]. We therefore turned to a selection of these published references to determine whether the presence of low numbers of GSCs had substantially perturbed CO development in the ltGLICO setting. Intriguingly, comparison of signatures from our assigned clusters against four primary brain and four organoid single cell datasets showed high levels of correlation between equivalent cell types (Additional file 1: Fig. S4). This suggests that the minimal levels of 320-GSC present in younger ltGLICOs has little effect on overall organoid development, which progressed in keeping with the published data.
Fig. 2
The organoid component of ltGLICOs recapitulates human neurodevelopment for up to 12 months. a UMAP of the CO cells from the ageing ltGLICO cohort colored by cell subtype clusters. b UMAP of the CO cells from the ageing ltGLICO cohort colored by cell type. c Dot plot of cluster marker gene expression used for cell type classification. Top; Cluster cell proportion by ltGLICO sample age. d Multiplex immunofluorescence staining of a representative neural rosette from a 1-month-old CO. DAPI; blue, NESTIN; cyan, EOMES; orange, NEUROD2; green. Scale bar = 100 µm. e Cell type and cell cycle stage proportions identified in each ltGLICO sample. f UMAP and Violin plot showing glioma tumor associated astrocytes (TAA) gene signature scores in single cells
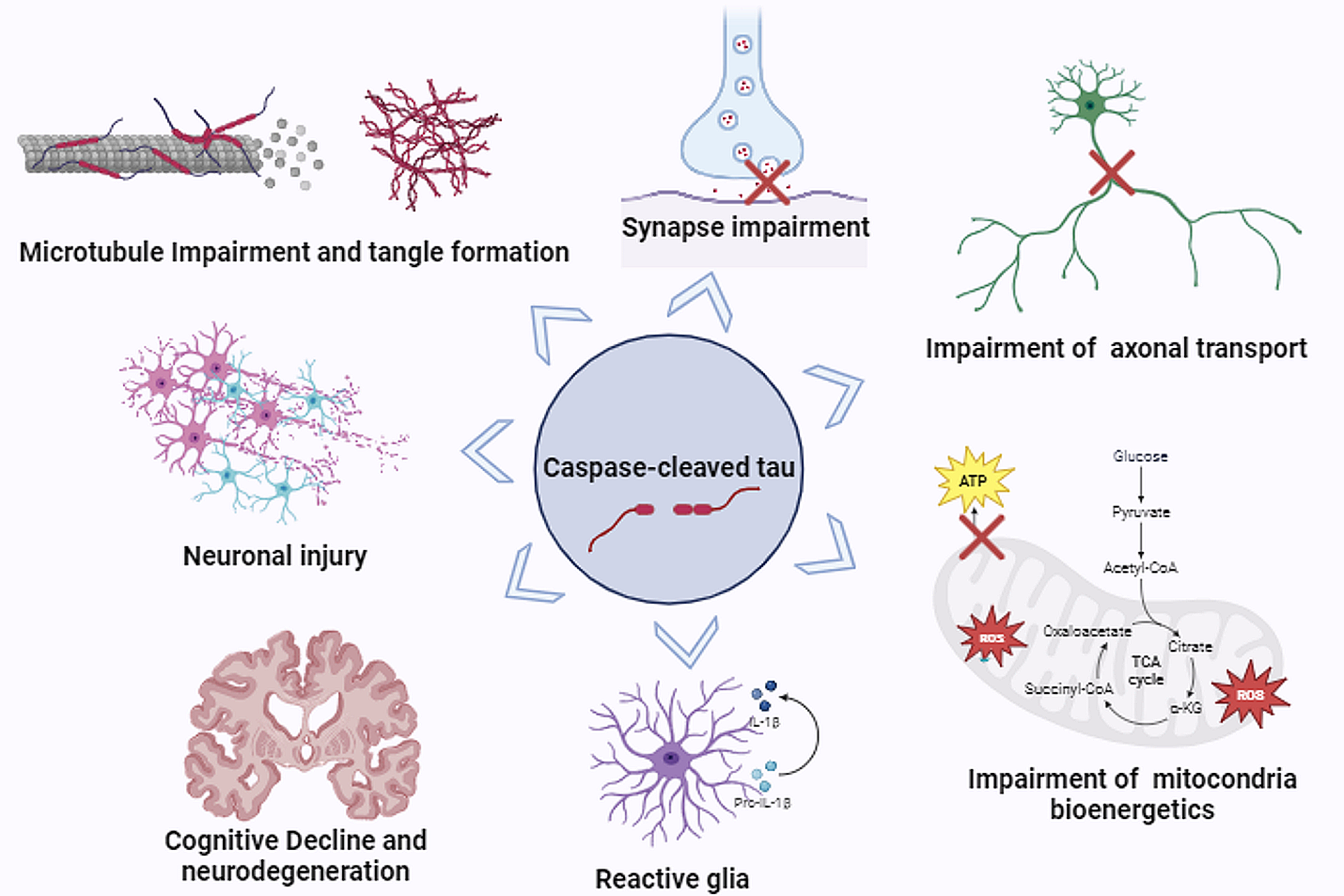
Cerebral organoids accurately recapitulate many aspects of human brain development [2], yet imperfect culture conditions, and the absence of functioning vasculature can lead to ectopic activation of cellular stress pathways [3]. We therefore scored ltGLICO cells for gene signatures associated with cellular stress and found that signature scores for hypoxia and glycolysis increased with ltGLICO age (Fig. 3a). As COs age they increase in size and become more densely cellular, which likely drives the steady increase in hypoxia that we confirmed by immunohistochemical staining for Carbonic Anhydrase 9 (Additional file 1: Fig. S3B). Chronic hypoxia has previously been linked to mitochondrial dysfunction and the generation of reactive oxygen species (ROS). Consistent with this phenomenon we saw a steady increase with age in signature scores associated with oxidative stress and the clearance of ROS, that was particular evident in astrocytes (Fig. 3a). This increase in ROS levels in aged ltGLICOs was validated by biochemical measurement of total ROS (Fig. 3b) and staining for mitochondrial ROS (Fig. 3c). When DNA is exposed to ROS, guanine can be modified to 8-oxo-dG, ultimately giving rise to DNA double strand breaks [30]. To detect whether elevated ROS levels were causing damage to ltGLICO cells we performed immunofluorescence against 8-oxo-dG and phosphorylated-yH2X (Fig. 3d–e). This showed marked increases in 8-oxo-dG and phosphorylated-yH2X over time indicating that oxidative stress, and its associated DNA damage, increases as ltGLICOs age. Whilst chronic hypoxia represents the most likely driver of oxidative stress, it is also possible that the presence of 320-GSCs in the microenvironment of CO cells may in some way contribute to this process. To confirm that CO hypoxia and oxidative stress increases with age in the absence of 320-GSCs we turned to published RNAseq data from 62 organoids generated from 6 different cell lines (Additional file 1: Fig. S3C). Transcriptomic signature scoring indicated that progressive oxidative stress was common to other brain organoid culture protocols in the absence of 320-GSCs.
Fig. 3
Aging ltGLICOs are characterized by increased cellular stress and oxidative DNA damage. a Scaled cell stress signature scores in H1-derived ltGLICO cells grouped by cell type and sample age. b Quantification of total ROS levels measured by 2′,7′-dichlorodihydrofluorescein diacetate (H2DCFDA)—assay. Graph shows mean + s.e.m. from four biological replicates. p-value = 0.0286, Mann–Whitney test. c Mitosox-Red staining for mitochondrial reactive oxygen species (ROS) levels in representative sections from young (4 month) and old (12 month) ltGLICOs; MitoSOX (red), and DAPI (Blue). d Immunofluorescence staining of ltGLICOs for 8-oxoDG (8-oxoDG in red, Phalloidin in green), representative images from a 2, 4, 9 and 18 month ltGLICOs. e Immunofluorescence staining of ltGLICOs for phospho-gamma-H2AX (yH2AX in red, DAPI in blue), representative images from a young (1.5 months) and old (18 month) ltGLICOs. Right, quantification of yH2AX positive cells per field of view at each CO age (plot shows mean + s.e.m.; 3 technical replicates). Scalebar = 200 µm
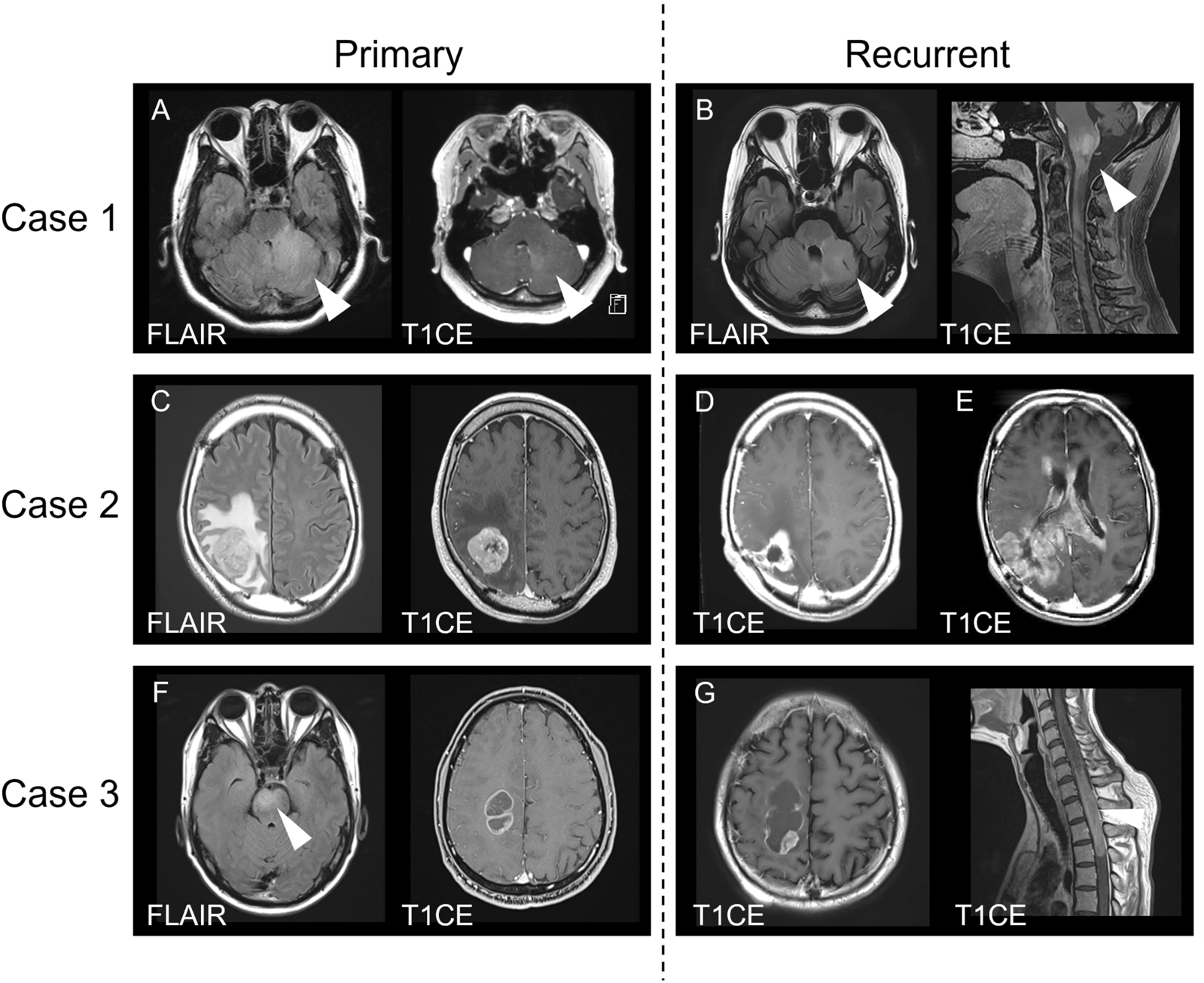
We next performed receptor-ligand modelling to query our scRNAseq data for candidate microenvironmental factors that may promote GSC growth (Fig. 4). Notably, 320-GSCs demonstrated their highest number of putative receptor-ligand interactions with organoid cells of the astroglial lineage including outer radial glia and astrocytes cells (Fig. 4a). Among the organoid-secreted ligands were numerous microenvironmental factors previously reported to play a pro-tumorigenic role in glioma (Fig. 4b). These included SPP1, FGF1/2, PDGFA and PTN [31,32,33,34,35] (Fig. 4c), several of these ligands are known to be upregulated under conditions of hypoxia and oxidative stress [36,37,38,39]. Furthermore, their expression levels peak during months 8–12 of ltGLICO culture coincident with the growth of the GSCs. Based on these findings we hypothesized that progressive hypoxia and oxidative stress may remodel the ltGLICO microenvironment to promote GSC growth by increasing the secretion of pro-tumorigenic ligands by glial cells.
Fig. 4
Receptor ligand modelling uncovers astrocyte expression of candidate pro-tumorigenic ligands. a Heatmap showing the total number of interactions between cell types in ltGLICO dataset obtained with CellPhoneDB. b Circos plot showing interaction between ligands secreted by organoid cell types and receptors expressed by 320-GSCs. c Violin plot showing expression of oxidative stress associated ligands (SPP1, PDGFA, FGF1, FGF2) in glial cells
To better understand the regulatory mechanisms underpinning a putative pro-tumorigenic role for hypoxic astrocytes, we next performed single-cell multiome-seq (scATAC and scRNA in the same cells) on a cohort of 8 ltGLICOs spanning 5–12 months. Computationally identified 320-GSCs were well separated from organoid cells in UMAP dimensional reductions calculated on RNA and ATAC independently (Additional file 1: Fig. S5A), or together and samples showed similar long latency periods before 320-GSC expansion (Fig. 5a). We next clustered cells and transferred cell type labels from the larger scRNAseq cohort. Cluster 10 was of particular interest since it was composed of mostly astrocytes, with some hypoxic and outer radial glia cells, and was enriched for cells from 8 to 12 month GLICOs—coinciding with the expansion of 320-GSCs. Furthermore, expression analysis of candidate pro-tumorigenic ligands (FGF1, FGF2, SPP1 and PDGFA) revealed enrichment in the cluster (Fig. 5b). We next used the chromosome accessibility data to identify cluster specific enrichment for transcription factor motifs (Additional file 1: Fig. S5B)—which identified enrichment for cell-type defining transcriptional regulators in the expected clusters: 320-GSCs were enriched for AP-1 transcription factors FOSL1/2 and JUN; excitatory neurons for NEUROD1/2; Inhibitory neuron progenitors for DLX2/5 and OPCs for ASCL1. For cluster 10, we considered transcription factors that had both motif enrichment and upregulated expression for the corresponding transcription factor, reasoning that these would be the most important factors in defining this pro-tumorigenic astrocytic state (Fig. 5c, d). Among the most enriched regulators were SOX9 and NFIA, both key regulators involved in the development and maintenance of astrocytes [40, 41], and RORA which is predominately expressed in adult astrocytes [42]. Other identified regulators are involved in reactive gliosis, such as SOX2 which plays a role in astrocyte activation after traumatic brain injury [43], and STAT3 which has been widely implicated in astrocyte activation and inflammation during neurological disease [44], and under hypoxic/ischemic conditions [45,46,47]. Another interesting regulator identified was NFATC2, an effector of the Calcineurin/NFAT pathway which is associated with chronic hypoxia in astrocytes during cerebellar vascular disease and is upregulated in astrocytes following CNS injury and disease [48]. Together these analyses suggest that astrocytes present in ltGLICOs are maintained by a regulatory network associated with reactive gliosis that characteristic of CNS disease and under conditions of ischemia.
Fig. 5
ltGLICO astrocytes are defined by regulators characteristic of reactive gliosis and ischemia. a UMAPs calculated on combined (weighted nearest neighbors) RNA and ATAC single cell data from 5 to 12 month ltGLICOs colored by genotype, age, clusters and cell type. b Expression density plots of pro-tumorigenic ligands (FGF1, FGF2, PDEFA and SPP1) in ltGLICOs. c Differential expression of transcription factors and enrichment of their associated motifs in cluster 10 astrocytes. d Motif plots of enriched regulators
To test the theory that chronic hypoxia promoted astrocytic secretion of pro-tumorigenic ligands, and to discount the possibility that the GSCs themselves were prompting the ligand secretion, we cultured standard COs—absent any GSC component—under hypoxic conditions (5% O2, n = 2) for 6 weeks and compared them to normoxic (~ 20% O2, n = 2) controls by scRNAseq. UMAP projections of cells colored by their oxygen culture conditions showed clear differences (Fig. 6a) and identifying cell types based on their marker gene expression (Fig. 6b–d) revealed changes in their cell type proportions. OPC/Oligos were completely absent, and InNPCs were depleted under hypoxic culture conditions likely indicating greater sensitivity to hypoxia for these cell types. By contrast, astrocytes—the primary cell type contributing to organoid-GSC crosstalk—were greatly expanded (Fig. 6c). As expected, scoring for gene signatures associated with cellular stress processes showed enrichment in hypoxic organoids (Fig. 6e). Of note was upregulation of the hallmark_protein_secretion gene set, as well as several associated with oxidative stress. This indicates that chronic hypoxia is the likely root cause of the oxidative stress phenotype seen in aging ltGLICOs, and COs more generally. Given the increase in the signature for protein secretion we next queried the major cell types present in both culture conditions (radial glial, astrocytes, excitatory neurons and Interneuron NPCs) for expression of candidate pro-tumorigenic secreted ligands (Fig. 6f). Interestingly the majority of these ligands showed upregulation in hypoxic astrocytes. Furthermore, scoring astrocytes for the glioma TAA gene signature [23] revealed that astrocytes kept under hypoxic conditions were significantly more similar to those observed in the glioma tumor microenvironment than their normoxic counterparts (Fig. 6g). Given that a pro-tumorigenic role for acidic-FGF1 has not previously been established for glioma, we evaluated the effect of recombinant FGF1 on 320-GSCs in growth factor depleted media, revealing a dose-responsive increase in 320-GSC growth (Fig. 6h). We observed similar results with a second patient derived cell line 1206-GSCs (Additional file 1: Fig. S5C).
Fig. 6
Hypoxia remodels the cerebral organoid microenvironment and increases the secretion of pro-tumorigenic ligands. a UMAP of hypoxic culture CO cohort colored by oxygen condition. b UMAP of hypoxic culture CO cohort colored by cell type. c Cell type proportions identified in each CO sample. d Dot plot of cluster marker gene expression used for cell type classification. e Scaled cell stress signature scores, single cells grouped sample. f Violin plots of candidate pro-tumorigenic ligand expression in hypoxic culture CO cohort. g Violin plot of glioma TAA signature scores in hypoxic vs control astrocytes. h Effect of recombinant acidic FGF1 on 320_GSC growth. i Proportion of H1-ESC and 320-GSC specific SNP fingerprints detected in control 5-month-old ltGLICO and 5-month-old ltGLICO treated for 6 weeks with 50uM Buthionine sulfoximine (BSO)
Together these results show that exposure of COs to chronic hypoxia increases oxidative stress and the population of astrocytes that secrete pro-tumorigenic ligands. Based on these findings we next tested whether pharmacological modulation of microenvironmental oxidative stress in ltGLICOs could accelerate GSC growth. ltGLICOs were cultured in 50uM buthionine sulphoximine (BSO), an inhibitor of glutathione-synthetase an enzyme critical for the clearance of ROS which we found to be upregulated in older ltGLICO astrocytes (Additional file 1: Fig. S5D). After 6 weeks treatment, we performed WES on a BSO-treated 5 month ltGLICO and a control sample. Comparison of the detected proportion of H1-320 and GSC-320 specific DNA fingerprints revealed a major increase in the proportion of 320-GSC DNA detected in the BSO treated ltGLICO, with Glioma driver mutations clearly detectable, whereas they remained below the threshold of detection in the control sample (Fig. 6i).
留言 (0)