
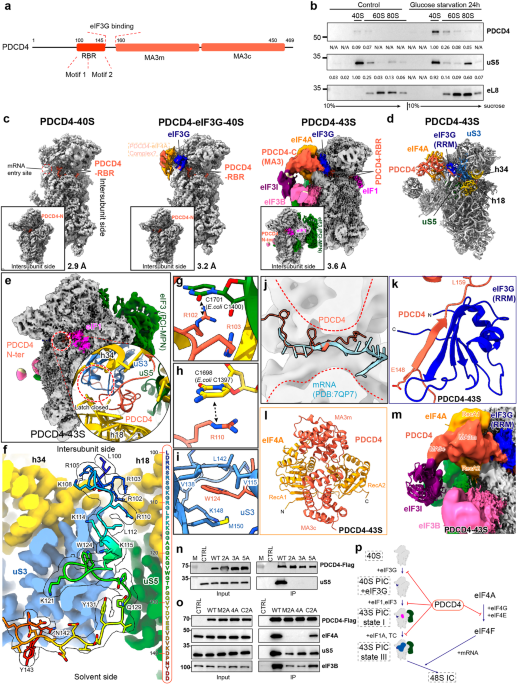
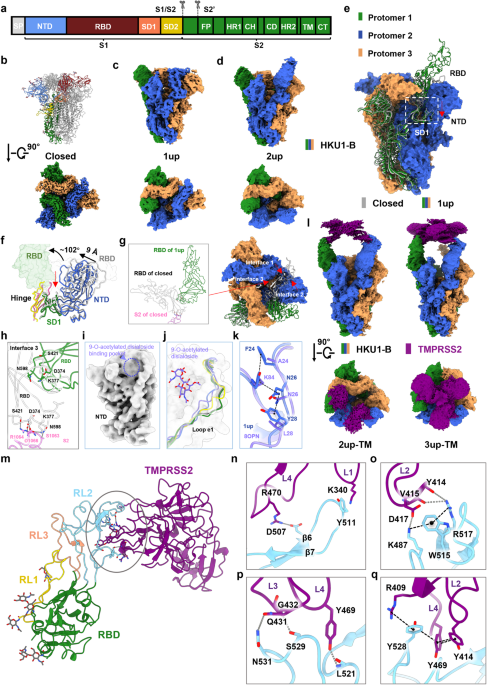
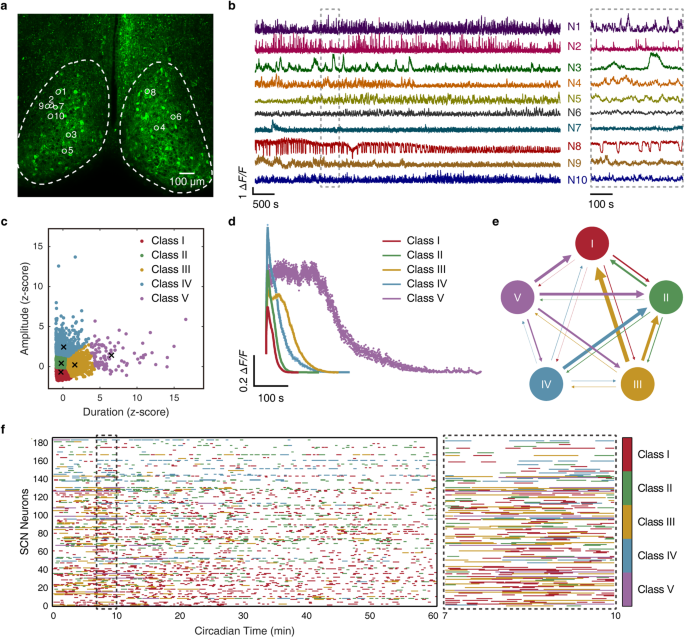
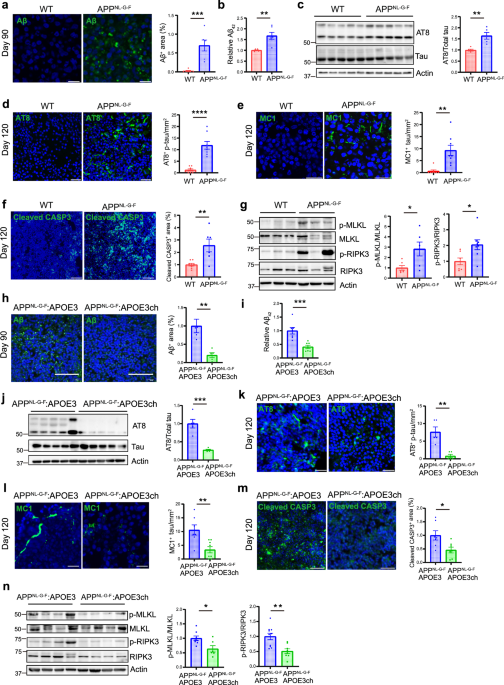
記住我


We sought to explore the mechanism of peripheral cancers communicating with the central nervous system. As the entry point of the study, several mouse allograft models were utilized, i.e., LLC lung cancer, RM1 prostate cancer, MC38 colon cancer, and 4T1 breast cancer. Notably, these allograft models have been commonly exploited in cancer immunology to investigate antitumor immunity and MDSCs in immune-competent wild-type mice.19,20 Mice were subcutaneously implanted with each cancer cell line, and brain tissues of tumor-bearing mice were then processed for the immunostaining of phospho-ribosomal protein S6 (p-S6), a specific marker for neuronal activation.21 We comprehensively assessed brain regions ranging rostrocaudally from olfactory bulbs to the brainstem. Surprisingly, we identified a shared pattern of brain responses under all the tumor-bearing conditions examined, including that in the paraventricular nucleus of the hypothalamus (PVN), a central brain region initiating efferent sympathetic signals (Fig. 1a, b). Further, responses of the suprachiasmatic nucleus (SCN), the left and right vestibular nucleus (Ve-L/-R), the left and right mesencephalic nucleus of the trigeminal nerve (Me5-L/-R), the red nucleus (RN), the hypoglossal nucleus (12N), and the left and right motor nucleus of the trigeminal nerve (5N-L/-R) were triggered in these allograft models (Fig. 1a, b; Supplementary information, Fig. S1a, b). On the other hand, there were no detectable neural activities in the brain regions related to visceral or metabolic signaling, e.g., the nucleus of the solitary tract (NTS), the left and right parabrachial nucleus (PBN-L/-R), and the arcuate nucleus (ARC) (Fig. 1c, d), exemplifying the specificity of cancer-induced brain activation. In parallel, we looked into the orthotopic allograft lung cancer model of LLC cells22 or the orthotopic prostate cancer model of RM1 cells.23 These two models of orthotopic tumors exhibited brain responses comparable to those in the heterotopic subcutaneous tumors (Supplementary information, Fig. S2a, b).
Fig. 1: An overlapping pattern of brain responses in multiple mouse models of peripheral cancers.
a–d Mouse allograft models of LLC, RM1, MC38, or 4T1 cells and the MMTV-PyMT mouse model were utilized. Brain responses were assessed by the p-S6 immunostaining. Representative images of the PVN, SCN, Ve-L/-R, and Me5-L/-R (a) or the NTS, PBN-L/-R, and ARC (c) were shown. Scale bars, 100 μm. Neural activities in the indicated brain regions were quantified (b, d). Data are shown as mean ± SD. One-way ANOVA test; ns, not significant; *P < 0.05. e, f A multi-omic screening for the cancer cell-derived factor(s) triggering brain responses. The scheme of integrating RNA-seq data of different cancer types and secretomic analyses of cultured cancer cells was illustrated (e). The published scRNA-seq dataset of mouse trigeminal ganglionic neurons (GSE213105) was exploited to identify candidate proteins for functional examination (f).
In addition to the allograft cancer models, we assessed the brain activation of MMTV-PyMT mouse, a standard genetic model of orthotopic breast cancer.24 Importantly, this genetic model exhibited the same set of responsive and non-responsive brain regions as the above allograft models (Fig. 1a–d; Supplementary information, Fig. S1a, b). These results suggested that despite their differential origins and mutational spectra, multiple types of peripheral cancers could induce an overlapping pattern of brain responses.
To determine whether the physical burden of a tumor might underlie the commonality of brain responses to different cancers, we implanted the wild-type mice with a pseudo-tumor made of medical-grade silicon. Although these pseudo-tumors effectively mimicked the volume and weight of “real” tumors, they failed to cause any neural activities in the signature brain regions, including the PVN, SCN, Ve-L/-R, and Me5-L/-R (Supplementary information, Fig. S3a, b). This observation mostly ruled out the involvement of physical properties of peripheral tumors in inducing brain activation.
Leukemia inhibitory factor (LIF) and galectin-3 (Gal3) as the cancer cell-derived factors triggering brain responsesWe considered the possibility that multiple cancer types might release the same factor(s) to trigger the overlapping pattern of brain activation. To pursue this hypothesis, we profiled the transcriptomes of tumors from LLC, RM1, MC38, and 4T1 allograft models and MMTV-PyMT genetic model by RNA sequencing (RNA-seq) (Fig. 1e), and identified ~2000 candidate genes exhibiting a shared expression pattern among different cancer types. In parallel, proteins in the culture media of LLC, RM1, MC38, or 4T1 cells were examined by proteomic analyses (Fig. 1e). We found that 114 proteins were commonly present in the secretomes of these cancer cell lines, 89 of which were included in the list of candidate genes identified by RNA-seq profiling. Among them, 61 candidate proteins were then filtered for being a bona fide secreted factor, i.e., possessing a signal peptide, 11 of which have the specific receptor(s) documented (Supplementary information, Table S1). We further analyzed the expression of these receptors in the published single-cell RNA-seq (scRNA-seq) dataset of mouse trigeminal ganglionic neurons (GSE213105),25 whose axonal projections control Me5-L/-R and 5N-L/-R, the signature regions activated by multiple cancer types. Through this multi-omic screening, three candidate proteins (LIF, pigment epithelium-derived factor, and thrombospondin-1) were obtained for in-depth functional examination (Fig. 1f).
We treated the non-tumor-bearing wild-type mice with each recombinant protein of the three candidates, testing whether one could recapitulate the same pattern of brain responses to peripheral cancers (Fig. 2a). Remarkably, a single intraperitoneal injection of 1 μg mouse LIF could effectively achieve the activation of signature brain regions, i.e., PVN, SCN, Ve-L/-R, Me5-L/-R, RN, 12N, and 5N-L/-R, for up to 24 h (Fig. 2b, c; Supplementary information, Fig. S4a, c). Meanwhile, reminiscent of that observed in the tumor-bearing mice, this LIF treatment did not cause detectable neural activities in the NTS or PBN-L/-R (Fig. 2d, e). In addition, plasma levels of LIF protein were significantly upregulated in all the allograft models of LLC, RM1, MC38, or 4T1 cells, as well as in the tumor-bearing MMTV-PyMT mice (Fig. 2f; Supplementary information, Fig. S2c). Moreover, increased plasma levels of LIF were detected in patients with multiple cancer types, e.g., prostate cancer, bladder cancer, ureteral cancer, non-small cell lung cancer, colorectal cancer, and renal cancer (Fig. 2g). These results supported LIF as a cancer cell-derived factor that triggers brain responses.
Fig. 2: LIF and Gal3 are the cancer cell-derived factors that cooperatively activate specific brain regions.
a–e Non-tumor-bearing C57BL/6 wild-type mice were administered with the indicated amounts of recombinant mouse LIF or Gal3 protein. Brain responses were assessed by the p-S6 immunostaining. The scheme of the experimental procedure was illustrated (a). Representative images of the PVN, SCN, Ve-L/-R, and Me5-L/-R (b) or the NTS and PBN-L/-R (d) were shown. Scale bars, 100 μm. Neural activities in the indicated brain regions were quantified (c, e). Data are presented as mean ± SD. One-way ANOVA test; ns not significant; *P < 0.05. f Mouse allograft models of LLC, RM1, MC38, or 4T1 cells and the MMTV-PyMT mouse model were utilized. Plasma levels of LIF and Gal3 in different mouse models were examined by enzyme-linked immunosorbent assay (ELISA). Data are presented as mean ± SD. One-way ANOVA test; *P < 0.05. g Plasma levels of LIF and Gal3 in human patients with different peripheral cancers were measured by ELISA. Data are presented as mean ± SD. One-way ANOVA test; *P < 0.05.
However, it came to our attention that the intraperitoneal administration of 1 μg recombinant LIF protein resulted in plasma levels of ~50 ng/mL, which would be 100-fold higher than those in the tumor-bearing mice. On the other hand, although a single injection of 2 ng LIF led to its plasma levels comparable to those of the tumor-bearing conditions, this low dose was not sufficient to elicit brain activation (Fig. 2b, c; Supplementary information, Fig. S4b, d). This phenomenon raised the challenging issue that additional cancer cell-derived factor(s) might exist to function cooperatively with LIF in neural activation. To this end, we moved on to laboriously screen the commercially available recombinant proteins of the remaining 60 candidate secreted factors (Supplementary information, Table S1) in the hope of identifying one that could induce neural activity under the condition of the low-dose LIF treatment. We managed to find that while the intraperitoneal injection of 5 μg galectin-3 (Gal3) alone only induced neural activities in the Ve-L/-R of non-tumor-bearing mice, it acted in a cooperative manner with 2 ng LIF to trigger responses of all the signature brain regions (Fig. 2b, c; Supplementary information, Fig. S4b, d). As an aside, both recombinant LIF and Gal3 proteins showed a relatively fast clearance in the blood (t1/2 of LIF = 2.9 min and t1/2 of Gal3 = 3.2 min). Notably, plasma levels of Gal3 were significantly upregulated in the allograft models of LLC, RM1, MC38, or 4T1 cells and the MMTV-PyMT genetic model (Fig. 2f; Supplementary information, Fig. S2c). Further, Gal3 plasma levels markedly increased in patients with different cancers (Fig. 2g). These results together identified LIF and Gal3 as the key factors released by multiple cancer types to communicate with the brain.
Blockage of LIF and Gal3 signaling to the brain inhibits tumor progressionWe next explored the disease relevance of cancer cell-derived LIF and Gal3 signaling to the brain. LLC, RM1, MC38, or 4T1 cells with the genetic deletion of LIF or Gal3 were generated by CRISPR/Cas9 and then tested in the mouse allograft models (Fig. 3a; Supplementary information, Fig. S5). Significantly, LIF knockout (KO) in these cells blocked the cancer-induced activities in the signature brain regions, such as the PVN and Me5-L/-R (Fig. 3b, c). Intriguingly, LLC, RM1, MC38, or 4T1 cells with the LIF deletion did not entirely abolish neural activities in the Ve-L/-R (Fig. 3b, c), which appeared in accordance with the above observation that Gal3 alone could stimulate this brain nucleus (Fig. 2b, c). In parallel, Gal3 KO in cancer cells strongly abrogated neural activities in all the signature brain regions of tumor-bearing mice (Fig. 3b, c). These results validated the cooperative action of cancer cell-derived LIF and Gal3 in mediating brain responses to multiple types of peripheral tumors.
Fig. 3: Cancer cell-derived LIF and Gal3 are essential for brain responses.
a–c LLC, RM1, MC38, or 4T1 cancer cells with LIF KO or Gal3 KO were generated by CRISPR/Cas9 and then tested in mouse allograft models. Brain responses were assessed by the p-S6 immunostaining. The scheme of the experimental procedure was illustrated (a). Representative images of the PVN, SCN, Ve-L/-R, and Me5-L/-R were shown (b). Scale bars, 100 μm. Neural activities in the indicated brain regions were quantified (c). Data are presented as mean ± SD. One-way ANOVA test; ns, not significant; *P < 0.05.
Blockage of brain activation by LIF KO or Gal3 KO delayed tumor growth in the mouse allograft models of LLC, RM1, MC38, or 4T1 cells (Fig. 4a). In further support of the critical role of LIF and Gal3 signaling, we daily treated the mice bearing “wild-type” tumors with EC330, a small-molecule inhibitor of LIF.26 This pharmacologic approach dampened brain responses in the signature brain regions, e.g., PVN, SCN, and Me5-L/-R (Supplementary information, Fig. S6a). Meanwhile, consistent with the findings with LIF KO, neural activities in the Ve-L/-R persisted in those tumor-bearing mice treated with EC330 (Supplementary information, Fig. S6a). Of importance is that the EC330 treatment delayed tumor growth in the mouse allograft models of “wild-type” LLC, RM1, MC38, or 4T1 cells (Supplementary information, Fig. S6b). Similarly, the mice bearing “wild-type” tumors exhibited no brain response in all the signature regions when treated with the Gal3 small-molecule inhibitor GB1107 (Supplementary information, Fig. S7a).27 Further, this pharmacologic inhibition of the Gal3 signal led to the strong suppression of tumor growth (Supplementary information, Fig. S7b). These results elucidated the function of cancer cell-derived LIF and Gal3 signaling to the brain in modulating tumor progression.
Fig. 4: Blockage of LIF or Gal3 signaling inhibits tumor progression.
a–c LLC, RM1, MC38, or 4T1 cancer cells with LIF KO or Gal3 KO were examined in mouse allograft models. Tumor growth rates under the indicated conditions were monitored (a). n = 8. Data are shown as mean ± SD. Two-way ANOVA test; *P < 0.05. MDSCs in the blood or tumors under the indicated conditions were quantified by fluorescence-activated cell sorting (FACS) (b). CD8+ T cells in the tumors under the indicated conditions were examined by FACS (c). Data are shown as mean ± SD. One-way ANOVA test; *P < 0.05.
Sympathetic signaling promotes MDSC generation for tumor progressionResearch has documented MDSCs, which can be further categorized as polymorphonuclear MDSCs (PMN-MDSCs) or monocytic MDSCs (M-MDSCs), in suppressing CD8+ T cell-mediated antitumor immunity.28,29,30 We found that the presence of PMN-MDSCs and M-MDSCs diminished in the blood and tumors of mouse allograft models of LIF KO or Gal3 KO cells (Fig. 4b). Accordingly, the recruitment of CD8+ T cells increased within the allograft tumors of LIF KO or Gal3 KO cells compared to their parental “wild-type” cells (Fig. 4c). These results revealed that blockage of cancer cell-derived LIF and Gal3 signaling to the brain could enhance antitumor immunity.
We thus sought to determine the neuroimmune mechanism afforded by such cancer-induced brain responses. MDSCs are primarily generated through myelopoiesis in lymphoid organs such as the bone marrow and the spleen. Because cancer cell-derived LIF and Gal3 cooperatively activated the PVN, a central brain region that controls efferent sympathetic action, we explored whether local sympathetic inputs in specific lymphoid organs might modulate MDSC generation. We took advantage of the Th-Cre;TrkAfl/fl mouse model, in which the NGF high-affinity receptor TrkA is deleted in sympathetic neurons. Our previous works with advanced imaging techniques showed the complete ablation of sympathetic inputs in the bone marrow and the spleen of Th-Cre;TrkAfl/fl mice.31,32Th-Cre;TrkAfl/fl (i.e., sympathetic ablation) and control Th-Cre;TrkA+/+ littermates were examined in mouse allograft models. We compared brain activation in these tumor-bearing mice, revealing similar levels of neural activities in the signature regions between sympathetic ablation and control conditions (Supplementary information, Fig. S8a, b). Further, there was no significant change in plasma LIF or Gal3 levels of the tumor-bearing mice with sympathetic ablation (Supplementary information, Fig. S8c). These findings substantiated that cancer-induced brain responses would act upstream of sympathetic signaling. However, tumor growth in the Th-Cre;TrkAfl/fl mice was significantly inhibited (Fig. 5a), which correlated with decreased MDSCs but increased CD8+ T cells in the blood and tumors of these mice (Fig. 5b–d). We further tested LIF KO or Gal3 KO cancer cells in Th-Cre;TrkAfl/fl and control littermates. Importantly, growth rates of LIF KO or Gal3 KO tumors were comparable between sympathetic ablation and control conditions (Fig. 5e, f), showing no additive effect of the genetic deletion of LIF or Gal3 in this context. These results supported the notion that LIF and Gal3 could act via sympathetic signaling to promote MDSC generation for tumor progression.
Fig. 5: Cancer cell-derived LIF and Gal3 act via sympathetic signaling to facilitate MDSC generation.
a–d Th-Cre;TrkAfl/fl (sympathetic ablation) and control Th-Cre;TrkA+/+ littermates were utilized for LLC or RM1 allograft models. Tumor growth rates under the indicated conditions were monitored (a). n = 10. Data are presented as mean ± SD. Two-way ANOVA test; *P < 0.05. MDSCs in the blood or tumors under the indicated conditions were quantified by FACS (b). CD8+ T cells in the tumors of LLC (c) or RM1 (d) allograft models were examined by FACS. Data are presented as mean ± SD. Student’s t-test. e, f LLC (e) or RM1 (f) cancer cells with LIF KO or Gal3 KO were assessed in the allograft models with sympathetic ablation. Tumor growth rates under the indicated conditions were monitored. n = 10. Data are presented as mean ± SD. Two-way ANOVA test; *P < 0.05.
We next exploited the surgical approach of sympathectomy to specifically remove sympathetic inputs in the spleen of mice (Fig. 6a), which were then utilized for allograft cancer models. Reminiscent of that observed with sympathetic ablation, plasma levels of LIF or Gal3 were not significantly affected by spleen sympathectomy (Supplementary information, Fig. S8c). On the other hand, this local removal of sympathetic inputs was sufficient to reduce PMN-MDSCs and M-MDSCs in the spleens (Fig. 6b) and also in the blood circulation (Fig. 6c) of tumor-bearing mice, suggesting that sympathetic signaling in the spleen facilitated the generation of MDSCs.
Fig. 6: Sympathetic signaling directly promotes the immunosuppressive function of MDSCs.
a–c C57BL/6 wild-type mice were subjected to spleen sympathectomy and then utilized for LLC or RM1 allograft models. Sympathetic axons in the spleen after sham surgery or sympathectomy were visualized by the immunostaining of tyrosine hydroxylase (a). MDSCs in the spleens (b) and the blood (c) under the indicated conditions were quantified by FACS. Data are presented as mean ± SD. Student’s t-test. d PMN-MDSCs or M-MDSCs were FACS-sorted from the spleens of tumor-bearing mice and in vitro treated with NE in combination with the β2-adrenergic receptor antagonist propranolol. mRNA levels of immunosuppressive genes were examined by qPCR analyses. Data are presented as mean ± SD. One-way ANOVA test. e Diagram of cancer cell-derived LIF and Gal3 hijacking the nervous system to promote tumor progression.
Finally, we FACS-sorted PMN-MDSCs and M-MDSCs from the spleens of mouse allograft models for in vitro cultures. Interestingly, the sympathetic neurotransmitter norepinephrine (NE) boosted the expression of immunosuppressive genes in MDSCs, e.g., iNOS2, Arg1, and Il-10. This neuroimmune effect could be entirely abrogated by the β2-adrenergic receptor antagonist propranolol (Fig. 6d). Meanwhile, we found that the plasma LIF or Gal3 levels of tumor-bearing mice were not affected by the exogenous NE treatment (Supplementary information, Fig. S8c), confirming that the release of LIF or Gal3 by cancer cells was independent of sympathetic signaling. These results elucidated that sympathetic signaling could directly promote the immunosuppressive function of MDSCs.
留言 (0)