
記住我
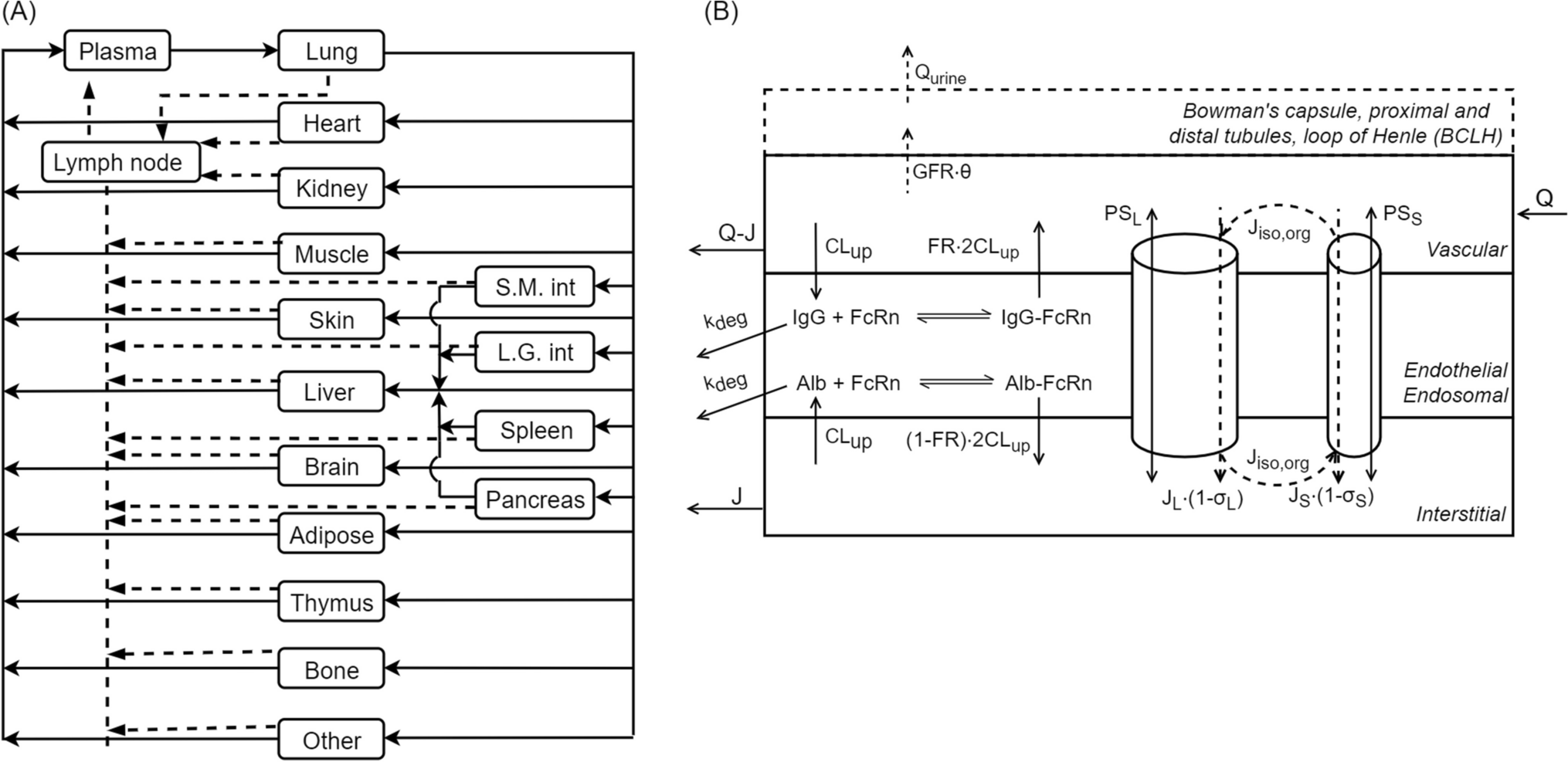


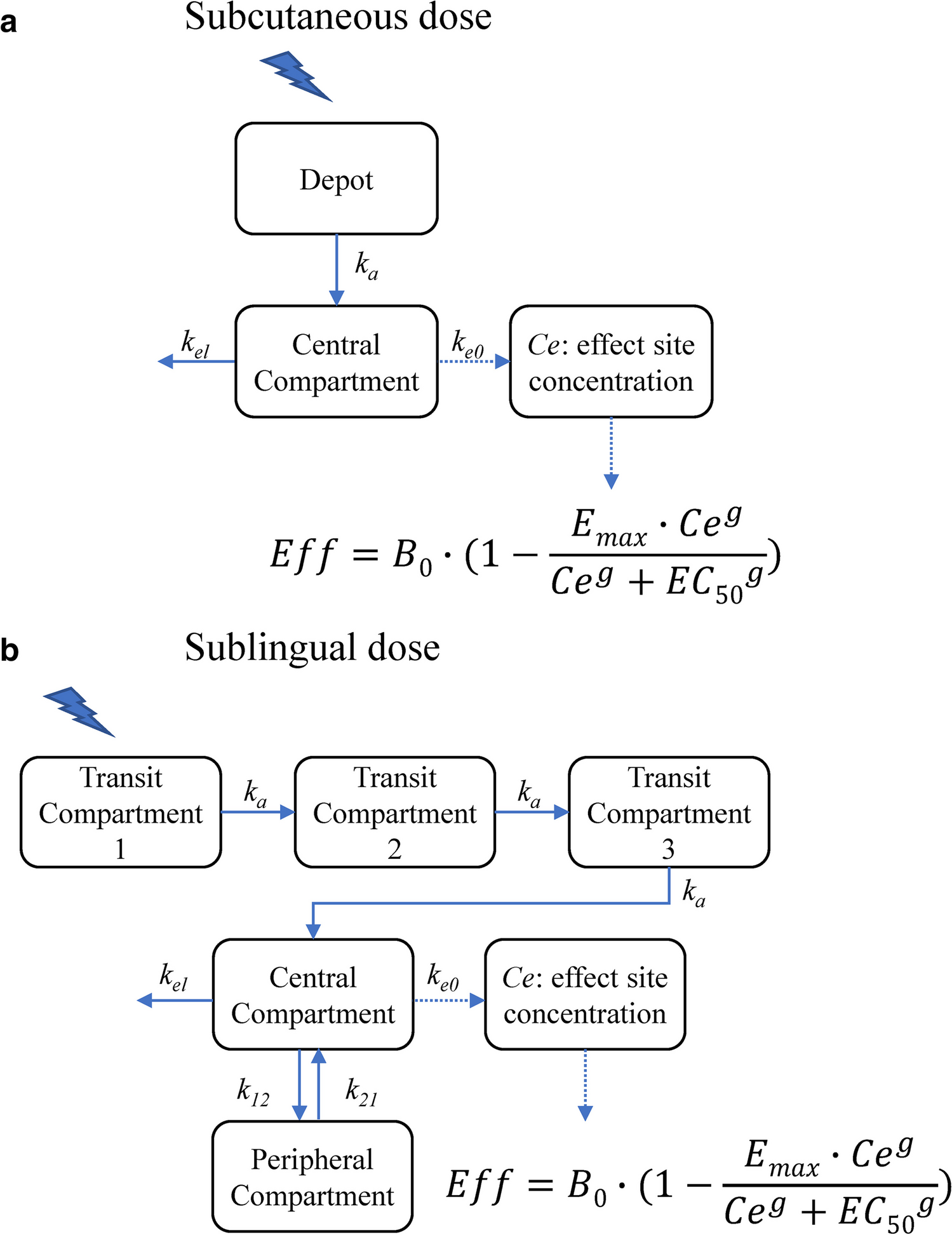
The structure of the mPBPK model, a schematic diagram of which is shown in Fig. 1, is mainly based on previously developed minimal and full PBPK models [11, 24,25,26,27]. It comprises plasma, lymph, and two lumped tissue compartments, tight and leaky. Tight tissue represents muscle, fat, brain, and skin, whereas leaky tissue the rest of the body tissues. The tissues with similar kinetics are lumped into respective tight or leaky compartments as previously proposed by Cao et al. [26]. As reported by Sarin et al., the blood capillary types in different tissues and organs can be distinguished based on the upper limit of pore size to the transvascular flow of large molecules [28]. The tissues with upper limit of capillary pore size approximately below 5 nM (brain, muscle, skin, and adipose) were lumped together as a compartment, and all other tissues were lumped as another compartment [28]. Arterial blood flow to tight/leaky tissue compartments is dependent on the vascular reflection coefficient for each tissue (\(_, _\)). The lymph flow out of each tissue compartment is collected in the lymph compartment, which is dependent on the lymphatic reflection coefficient (\(_\)). The lymph flow rate to each tissue compartment (\(_, _\) ) is calculated as a sum of individual tissue lymph flow rates reported previously [27]. Lymph flow from tight/leaky tissues is delivered back to plasma via the lymph compartment. Each tissue is divided into vascular, endosomal, and interstitial spaces. The unbound drug and drug-antigen complex drains from the interstitial space into the lymph compartment and is delivered back to the plasma and the systemic circulation. In our model, plasma is nested with an endosomal sub-compartment (Fig. 1), which represents endothelial cells in the systemic vascular space as discussed in previous models [24, 29]. Endothelial cells found in blood vessels can be different from the ones present in the organs, which add another dimension of complexity to our model [29].
Fig. 1
A schematic diagram of the minimal PBPK model developed. Minimal PBPK model (1) consists of plasma, lymph, tight tissue, and leaky tissue compartments. Each tissue compartment is divided into vascular, endosomal, and interstitial space as shown in the right (3). The plasma compartment is nested with endosomal space present in the systemic vascular endothelial cells. Monoclonal antibody (A) can intravenously (IV) administered in the plasma compartment. mAb (A) interacts with antigen (T) to form an antibody-antigen complex (A-T). mAb (A), soluble antigen (T), and complex (A-T) enter the nested endosomes in plasma through periodic pinocytosis/exocytosis processes (2). Free mAb (A) and mAb-antigen complex (A-T) binds to soluble and membrane antigens (red diamonds), undergoes pinocytosis into the endothelial cells, binds to FcRn (black rectangle) interacts with negatively charged membrane proteins (dashed line), and transports through the paracellular pores based on their size. In the endosomal space, mAb binds to FcRn and soluble target receptors at slightly acidic pH (pH = 6). Free mAb in the endosomes degrades through lysosomal degradation. Unbound antigen and mAb-antigen complex catabolize within endosomes. FcRn-bound mAb and FcRn-bound mAb-antigen complex are recycled back to vascular and interstitial space of the tissues. mAb undergoes size-based renal clearance in leaky tissue compartment. are vascular reflection coefficients. L1 and L2 are lymph flow rates in the tight and leaky tissue, respectively. L is the lymph flow rate. A representation of nested endosomes in the plasma compartment is provided (2) Antibody (Y-shape) interacts with soluble targets (red diamond) in plasma compartment to form antibody-receptor complex. In the nested endosomes, antibody freely interacts with soluble targets and FcRn receptor (black rectangle) at pH = 6. The unbound antibody degrades at a rate . The soluble receptors and antibody-receptor complex are cleared through catabolism at a clearance rate, . Subfigure (3) created with BioRender.com (Color figure online)
Model dynamicsFcRn/antigen bindingThe free mAb binds to the soluble antigen in plasma to form a soluble drug-antigen complex. The free mAb, antigen, and the mAb-antigen complex are taken up via pinocytosis into the nested endosomal sub-space in plasma, where free mAb and mAb-antigen complex can bind to FcRn receptor at slightly acidic pH (pH = 6) in the endosomes. The model assumes that available FcRn concentration in tissue and plasma endosomes remains same but varies with species [27] whereas the administered mAb are assumed to bind to FcRn at only slightly acidic, pH = 6, in the endosomes [24]. The FcRn-bound mAb and FcRn-bound mAb-antigen complex are recycled back to plasma and salvaged from lysosomal degradation. The unbound mAb within the endosomes is degraded via a first-order rate constant by lysosomal degradation, whereas unbound antigen and mAb-antigen complex is catabolized in the endosomes. The unbound mAb and mAb-antigen complex circulate to the tissue compartments and enter the vascular sub-space in the tissues. The unbound drug can bind competitively with soluble and membrane-bound antigens. The soluble targets are assumed to be synthesized both in plasma and in vascular space in the tissues, whereas membrane-bound targets are assumed to be synthesized in the tissue vascular space [24]. The membrane-bound mAb-antigen complex is assumed to internalize at the same rate as the rate of antigen degradation as drug often times does not affect complex internalization [1]. The drug target complex and unbound target are assumed to be catabolized at the same rate (\(C_\)) [24]. The mAb-antigen complex internalized in the tissues, and the internalized molecules are assumed to be metabolized further but were not explicitly modeled. The model does not account for a cellular space to describe downstream processes, and remains a current limitation of the model. The soluble mAb-antigen complex is taken up by tissue endosomes via pinocytosis. In the tissue endosomes, the unbound mAb and mAb-antigen complex interact with FcRn. The FcRn-bound mAb and FcRn-bound mAb-antigen complex are recycled back to both vascular space and interstitial space of the tissues.
Size-based transportIn our model, the unbound mAb present in the tissue vascular space transports to the interstitial sub-space in the tissues based on the two-pore model. Rippe and Haraldsson proposed the two-pore theory to study the transcapillary movement of large molecules through the paracellular pores in the tissues [30]. The tissue vasculature is assumed porous, and the radius of pores is loosely classified into small pores (40 nm) and large pores (220 nm) [10]. Large molecule drugs are transported through these two sets of pores in a size-dependent manner. The two-pore theory has been successful in explaining the extravasation of antibodies into the interstitial space of the tissues previously [9, 10, 30, 31]. Li and Shah presented the two-pore PBPK model, using de novo derived parameters, to predict the plasma PK of different sized proteins (13–150 kDa) without any parameter estimation [10]. Our mPBPK model includes size-based transport of large molecule drugs through small and large pores. The parameters such as permeability-surface area product, Peclet number, vascular reflection coefficients, fractional tissue lymph flows through pores are dependent on molecular weight and molecular size of the antibody and are calculated using derived equations (eqs. A40–A48) in [10].
Charge-based interactionThe charge-dependent effect on PK is reflected in the mPBPK model in the following processes. First, the charge variations of mAb affect the rate of pinocytosis or uptake into the endosomes, which is accommodated by multiplying a scaling factor (\(_\)) with the pinocytosis rate [22, 32]. Second, the unbound mAb undergo non-specific interactions with negatively charged cell membrane proteins (\(}}_},}}\)) in the tissue vasculature [22, 33]. The total concentration of membrane protein receptor (\(_\)) is assumed fixed across tissues [22]. The unbound mAb with a net positive charge is assumed to have high affinity for negatively charged cellular components, whereas mAb with a net negative charge is assumed to have lower affinity for cellular components due to repulsive forces between them. The non-specific binding between mAb and membrane proteins is characterized by an association rate constant (\(_\)), which is assumed equal to the mAb-FcRn association rate constant (\(_\)) [33], and a dissociation rate constant (\(_\)). Third, slight positive or negative charge on the mAb affect the volume of distribution in the interstitial space in the tissues [25, 34]. The change in volume of distribution for charge-variants is included in the model by multiplying a scaling factor (\(_\)) to the volume of interstitial space [23, 25].
Model ParametersOur mPBPK model structure remains the same across species. The model equations are provided in the Supplementary File, eq. A1–A39. The model parameters for the mPBPK model are loosely divided into two groups, physiology-based parameters and physicochemical or compound-based parameters. Additionally, some compound-specific parameters like molecular weight, size, and charge are related to model parameters through derived and empirical equations. The compiled list of physiology-based parameters is given in Table 1 in the Supplementary File. The physiological parameters such as body weight, volumes, flow rates etc. is dependent on the species of interest and adopted from respective literature sources [24,25,26,27]. The kinetic rate constant parameters such as pinocytosis rate, FcRn recycling rate, catabolic rate are assumed same across tissue compartments [27], unless stated otherwise. The kinetic rate constants such as endosomal uptake rate (\(_, _\)) were estimated among a range provided previously [27]. The endosomal recycling rate (\(C_\)) for nested endosomes in plasma and tissue endosomes were calculated using endosomal transit time (8 min) and volume of endosomes in mice as discussed by Yuan et al. [24]. The target-specific parameters such as baseline expression (\(IC_\)), half-life (\(C_\)), and internalization rate (\(_\)) are considered in the model. The rate of synthesis of soluble and membrane-bound antigens is calculated as a product of baseline concentration and half-life of the antigen. The baseline concentration of soluble and membrane-bound antigens was obtained from previous studies [35, 36]. The half-life of the soluble antigens is assumed to be 2 h based on [36], and half-life of membrane-bound antigens is estimated between 10 and 40 h [36, 37].
The compound parameters vary with drug’s physicochemical properties. In our mPBPK model, molecular weight (\(}\) ) of drug, Stoke’s radius (\(_\)) of drug, net surface charge on drug, fraction of interstitial volume available (\(_\)) for drug, association rate and dissociation rate constants for antibody-FcRn binding at slightly acidic pH (pH = 6) (\(_, _\)), association rate and dissociation rate constants for antibody-antigen binding at physiological pH (pH = 7.4) (\(_, _\)) and slightly acidic pH (pH = 6) (\(_, _\)), and dissociation rate constant for non-specific drug interactions are considered (\(_\)). \(_\mathrm_\) values for antibody-FcRn binding in acidic pH and \(_ , _ ,_,\mathrm_\) values for antibody-antigen binding are obtained from published experiments [22, 35]. These compound-specific properties of the drug are provided in Table 1 in the Supplementary File.
The derived and empirical equations are used to incorporate the effect of MW, size, and charge on antibody pharmacokinetics. Li and Shah have previously incorporated size-dependence based on a two-pore hypothesis [10]. The two-pore theory provides transport equations to derive parameters such as re-circulation rate, fluid flow rate through different sized pores, vascular reflection coefficients for different sized pores, permeability-surface area product, Peclet number etc., which are used in the model to describe the size-based transcapillary movement of antibodies through paracellular pores without any parameter estimation. Additionally, a renal clearance parameter (\(C_\)) is empirically-derived using a quantitative relation between glomerular sieving coefficient and Stoke’s radius of an antibody (Fig. A2). The fixed values of size-dependent model parameters are given in Table 2. In our model, effect of charge on mAb PK is included through a set of empirically-derived parameters (\(_, _, _\)) and relate them to the antibody charge (Fig. A3-A4). We provide additional detail on equations and methodology used to derive these parameters in the Model Development and Supplementary File.
Model developmentThe proposed mPBPK model was fitted and validated using published experimental data from multiple sources. We performed model fitting only where necessary as most model parameter values are known a priori. The mPBPK model parameters were fitted against published data for the following cases, (1) non-specific IgG antibody binding to FcRn in mice (\(_, _, _,\mathrm _\)) [16], (2) non-specific IgG antibody binding to FcRn knockout mice (\(_\)) [16], (3) charge-variants of non-specific IgG with an intact Fc region (\(_,K}_, _, _\)) [22], and (4) anti- carcinoembryonic antigen (CEA) IgG in mice (\(_, _\)) [37]. The mPBPK model was evaluated using the fitted and derived parameters for the following cases, (1) non-specific IgG antibody binding to FcRn in mice [38], (2) size-variants of non-specific IgG with or without intact Fc region, (3) charge-variants of non-specific IgG with an intact Fc region [18, 39]. To compare the predicted concentration in plasma and tissues, we digitized observed data for each case from published sources using WebPlotDigitizer [40]. Most studies report observed concentrations in individual tissues. The concentration in different sub-spaces in the tissue such as vascular, interstitial, or endosomal space were often not measured. Therefore, we calculated observed total tight tissue concentration by adding the observed drug amount in brain, muscle, skin, and fat tissues and dividing the sum by total tight tissue volume. The observed total leaky tissue concentration was calculated in a similar manner for other tissues. For each case, we calculated the total predicted tight tissue and leaky tissue concentrations using Eqs. 1 and 2, where \(_, _\) are endosomal concentrations, \(_, _\) are vascular concentrations, \(}}_}1}, }}_}2}\) are interstitial concentrations in tight and leaky tissue compartments, respectively. We acknowledge that the total tissue concentration of antibodies may not be the most appropriate to correlate with therapeutic effect of an antibody [41]. However, model development for each scenario was limited by the available published data. The total tissue concentrations were compared against observed total tissue concentrations in leaky and tight tissues. The prediction error between predicted and observed plasma and tissue concentration is calculated as a sum of squared error (SSE) normalized by the mean of the observed data. For parameter estimation, we used a multi-start non-linear least squares optimization method (fmincon) in MATLAB, where 10 starting values for each parameter were sampled using Latin hypercube sampling method in MATLAB. The best fitted parameters are obtained based on the least SSE between prediction and data.
$$}_},}}} \, = \,\frac}_}1}} }_}1}} + }_}1}} }_}1}} + }_}1}} }_}1}} )}}}_}1}} + }_}1}} + }_}1}} }}$$
(1)
$$}_},}}} \, = \,\frac}_}2}} }_}2}} + }_}2}} }_}2}} + }_}2}} }_}2}} )}}}_}2}} + }_}2}} + }_}2}} }}$$
(2)
Local sensitivity analysisFirst, we performed a local sensitivity analysis on the mPBPK model to obtain the most sensitive parameters. Each parameter value was increased by 20% from its original value, and a relative percent change (\(\mathrm\left(\mathrm\right)\)) in plasma and tissue exposure (AUC) was calculated as shown in Eq. 3, where \(}}_},+20\mathrm}\) is AUC calculated with perturbed parameter value, and \(}}_}}\) is AUC calculated with original parameter value. Fig. A1 in the Supplementary File shows the relatively sensitive parameters and respective change in AUC. The perturbed parameter that causes a relatively higher change in exposure is considered more sensitive. The most sensitive parameters were \(_, _, _,\mathrm _\) in the model and were fitted to the non-specific FcRn binding IgG1 dataset [16].
$$\Delta }\left( } \right)\, = \,\frac}_}, + 20}}} - }_}} }}}_}} }} \times 100$$
(3)
Model fitting and validationFor Case (1), we fitted the model to a non-specific IgG1 dataset in mice, where IgG1 does not bind to a specific target, here target-mediated specific clearance was not included in the model. Case (1) was validated using a validation dataset for non-specific IgG1 in mice [38]. Case (2) demonstrates the effect of FcRn knockout on IgG PK in plasma and tissues. Here, we recalibrated the rate-limiting parameter \(_\) to capture the faster clearance of IgG in absence of FcRn receptor in mice [16].
The effect of size of an antibody was reflected in the model using a transcapillary two-pore clearance from vascular to interstitial space in tissues, and a size-based renal clearance term. The size-based clearance through large pores and small pores is calculated using derived equations based on the two-pore hypothesis (eqs. A47–A48) [10]. For smaller sized antibody fragments with antibody size less than 4 nm, renal clearance plays an important role in non-specific clearance of an antibody [11]. The size-based renal clearance (L/h) is calculated using Eq. 4, where GFR is the glomerular filtration rate of kidneys in mice (L/h), and ϴ is the sieving coefficient. We fitted an empirical relationship between ϴ and Stoke’s radius (\(_\)) (Fig. A2) using data provided by Haraldsson et al. [42]. For larger antibodies (> 4 nm), clearance through kidney is insignificant [10]. The effect of size of various IgG and IgG fragments on plasma and tissue PK was validated using published data for non-specific IgG ranging from 50 to 150 kDa. For this case, no model fitting was necessary.
$$}_}}} \, = \,} * $$
(4)
The effect of charge on antibody PK is reflected in the model through a modulation factor (\(_\)) for interstitial volume of distribution, a modulation factor (\(_\)) for pinocytosis rate, and a charge-dependent non-specific equilibrium dissociation constant (\(_\)). Previous PBPK models have related charge with pinocytosis [22, 32, 43] and non-specific interactions [4, 22, 33]. Recent work by Liu et al. provided a good description of surface charge modifications on antibody disposition [18]. In their work, the complementarity determining regions (CDRs) of an antibody were systematically engineered to create a series of variants with an isoelectric point (pI) range of 6.3–8.9 that had a variable Fv charge identified using protein sequences [18]. Liu and Shah used part of this dataset to model the effect of charge on pinocytosis and non-specific interactions [22]. In the mPBPK model, we use this dataset to incorporate the effect of charge on volume of distribution in addition to pinocytosis and non-specific interactions. We use this published data for three different charge-variants of an antibody (150 kDa) to fit \(_\), \(_,\) and \(_\) parameters in the mPBPK model. The neutral IgG variant had a zero net surface charge, a positive IgG variant had a + 5 net surface charge, and negative IgG variant had a -8 net surface charge. The non-specific interactions between an IgG and negatively-charged membrane proteins (\(_\)) are incorporated using a dynamic set of equations, where concentration of total membrane protein receptor (\(_\)) is assumed fixed. \(_\) is estimated in our model for the neutral IgG dataset [22], and kept constant for other charge-variants of IgG. \(_\) is set to 1 for neutral IgG and estimated for slightly positive and slight negative charge-variants of IgG. We fixed \(_\) to 1 in nested endosomes in plasma, as charge did not affect IgG uptake in nested pl
留言 (0)