
記住我


As we age, the functionality of our major organs declines. Aging-associated phenotypes in all cell types of the body emerge as a result of cellular and tissue microenvironment changes, driven by various genetic and environmental stressors. Numerous hallmarks of the aging process have been identified over the years, including genomic instability, telomere attrition, mitochondrial dysfunction, cellular senescence, and stem cell depletion [1]. Cardiovascular health is particularly impacted by these manifestations of aging, due to (at least in part) the compromised health of endothelial cells (ECs). ECs form the inner lining of our blood vessels and play a crucial role in safeguarding and maintaining the proper function of all organs and physiological systems in our body by acting as barriers, filters, homeostatic regulators, and mediators of immune cell trafficking and cell–cell communication. The aging process affects the endothelium in countless ways, for instance via remodeling of the vascular wall, oxidative and nitrosative stress, and impairment of angiogenesis (we refer to Ungvari et al. [2] and Brandes, Fleming and Busse [3] for in-depth reviews on this topic).
To gain deeper insights into the molecular changes associated with endothelial aging, it is imperative to understand the fundamentals of EC behavior, molecular circuitries, and interactions across lifespan as well as in aging-associated disease conditions. Numerous methodologies and models have been used over the years to, typically, profile one or a handful of aging-associated features of the endothelium at a time, usually in ECs derived from one particular tissue or vascular bed (i.e., the sum of the blood vessels supplying an organ or region). For the longest time, technological challenges have however hindered our ability for a highly precise and comprehensive investigation of EC biology at scale, and in an untargeted fashion. Fortunately, recent advancements in single-cell omics have paved the way for creating detailed tissue atlases and examining single cells in health and disease. In that light, numerous atlasing efforts have drastically increased our understanding of the heterogeneous vascular endothelium, now widely appreciated to exhibit distinct gene and protein expression profiles based on its tissue of origin and disease status [4,5,6,7,8,9,10,11,12]. Our knowledge of the precise molecular changes in response to age-related challenges in ECs from individual organs, however, remains poorly understood. Obtaining a high-resolution view of the aging endothelium, by exploiting single-cell technologies, will be imperative for (i) the identification of specific EC subtypes and gene expression programs influenced by the aging process, (ii) the unraveling of mechanisms underlying the initiation and progression of age-related EC decline, (iii) the definition of new markers indicative of various stages of vascular lifespan, and, ultimately, (iv) the discovery and development of innovative molecular, gene, and EC-centered therapies for rejuvenation and/or treatment of age-related (vascular) diseases.
While we are still far from achieving those goals, the number of studies exploring aging of tissues using single-cell omics is steadily expanding, and increasingly incorporating the vascular endothelium. Here, we provide an overview of the current state of the literature regarding the heterogeneous adaptations of the vascular endothelium over the course of aging. We highlight findings from single-cell transcriptomics studies, complemented with high-resolution insights obtained using alternative methods, to shed light on pressing, emerging questions in the field of vascular aging. For instance, does the process of aging affect ECs in different tissues or vascular beds in a similar way? Is there a pan-EC aging phenotype or signature across tissues? What is the true age of ECs in our body, and do all ECs exhibit a similar degree of turnover? Are ECs more or less prone to acquire aging-associated phenotypes (e.g., senescence, apoptosis) in comparison to other cell types? Lastly, we discuss the future perspectives and challenges ahead in uncovering vascular aging-associated endothelial heterogeneity.
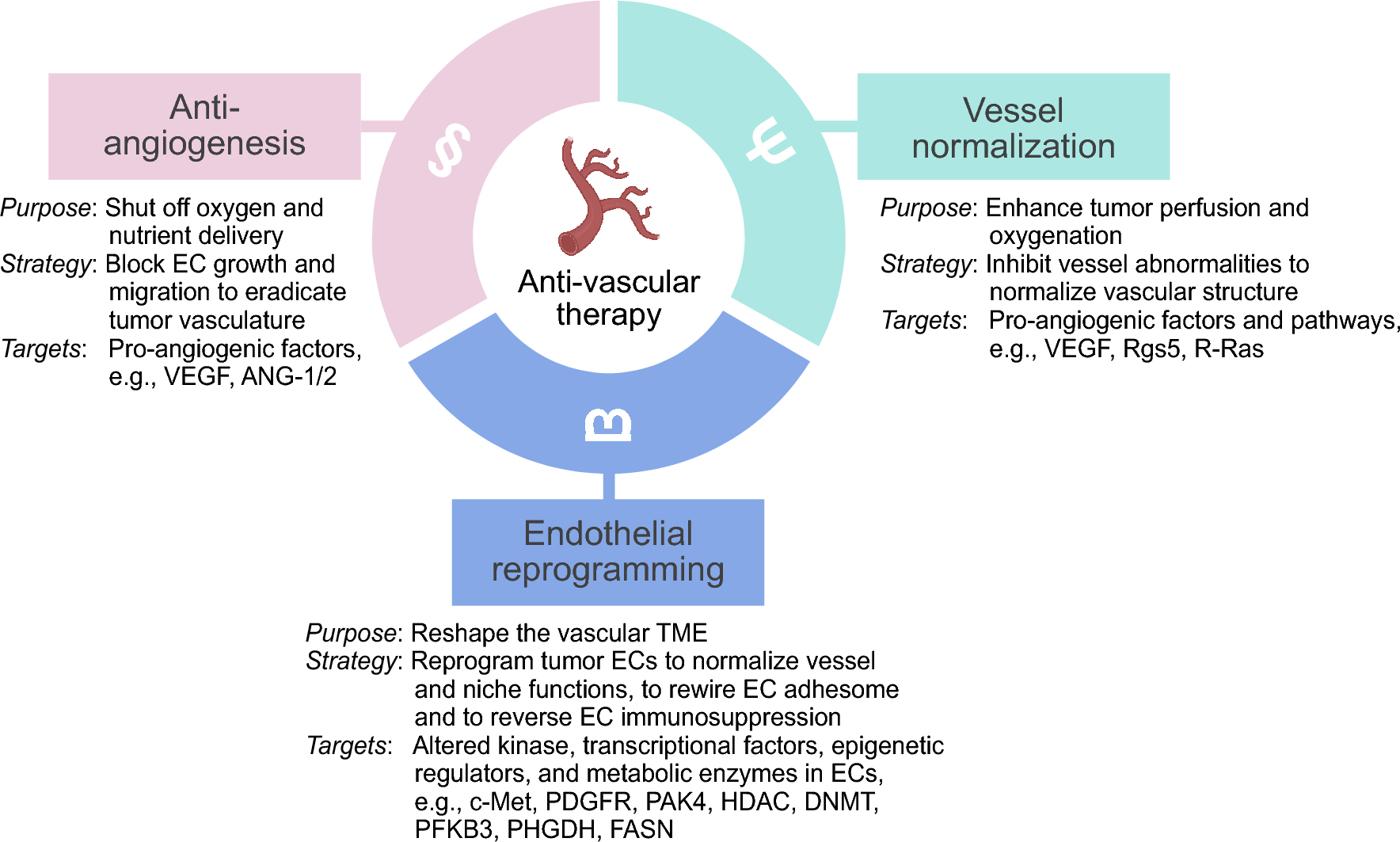
EC heterogeneity and aging in the single-cell eraComprehensive atlasing efforts dedicated to exploring the endothelial transcriptome across tissues and ages are yet to be conducted. Nevertheless, recent endeavors have started to reveal the first insights into diverse responses of ECs and their subtypes to the aging process in individual organs and tissues. In the subsequent sections, we will highlight the reported findings thus far for several tissue vasculatures. Across studies, “core” aging-associated changes of the endothelium include the loss of microvascular marker gene expression and a reduced abundance of (capillary/microvascular) ECs, as well as increased expression of inflammation and immunoregulatory gene signatures. These common changes are accompanied by a wide range of tissue-specific features of the aging endothelium, which we summarize in Fig. 1 and Table 1.
Fig. 1
Global and tissue-specific changes of the healthy aging endothelium. Middle: Core aging-associated changes in the endothelial landscape, identified in single-cell transcriptomics studies across different tissues (human and/or primate and/or mouse), include a decreased abundance of ECs (in comparison to other cell types) with increasing age, typically via a reduction in capillary/microvascular subtype abundance. Gene signatures associated with immunoregulatory processes and inflammation are generally increased in aging ECs across organs and tissues. Tissue-specific changes of the endothelium with increasing age are indicated in circled boxes next to each tissue. Phenotypes shared among two–three tissues are indicated at the bottom, including elevated expression of senescence-associated gene signatures (FRS, heart, kidney, lung), either a reduced (muscle, heart) or increased (brain, FRS) expression of angiogenic gene signatures, reduced expression of DNA repair signatures (skin, FRS), fewer interactions between ECs and other cell types (skin, FRS), increased transcriptional noise (brain, heart, lung, skin), and increased expression of VWF (lung, brain) or IL7 (heart, lung), as specified for each tissue by colored icons. EC = endothelial cell; ECM = extracellular matrix; DEG = differentially expressed gene; FRS = female reproductive system; Klfs = Krüppel-like factors; TF = transcription factor
Table 1 Common and tissue-specific changes in the healthy aging endothelium transcriptome landscapeBrainBrain endothelial cells (BECs) are key components of the cerebral microvascular system, which provides a continuous supply of nutrients and oxygen to fulfill the high energy requirements of the brain [13]. As the primary cell type of the blood–brain barrier (BBB) and neurovascular unit, BECs have an essential role in the regulation of molecular transport in and out of the brain, protection of the brain from pathogens, and in the regulation of cerebral blood flow [14,15,16]. Representing one of the very first tissues for which the endothelium was profiled at single-cell resolution [17], we now have an in-depth understanding of endothelial arteriovenous zonation and the associated EC subtypes and gene expression patterns in the mouse and human brain [18, 19]. Aging has been associated with impaired brain function and an increased susceptibility to cognitive decline and neurodegenerative disease [20]. The age-related dysfunction of BECs contributes to deterioration of the cerebral microvascular system, and is responsible for increased BBB permeability [15, 21, 22], decreased cerebral blood flow [23], and impaired neurovascular coupling [13, 24], all of which are hallmarks of both normal aging and neurodegenerative diseases [25, 26].
Healthy agingA wide range of single-cell transcriptomics studies of young versus aged disease-free mouse brains revealed endothelial aging-associated transcriptional profiles [16, 26,27,28,29,30,31]. While some of these studies report that the overall abundance of ECs, or their arteriovenous zonation patterns, were not changed between young and aged animals [16, 29, 31], others reported a decrease in capillary-venous ECs (i.e., EC subtype with mixed distribution of both capillary and venous marker genes) in brain tissue from old animals [30]. In terms of aging-associated gene expression changes, a common finding in these studies was a typical increase in expression for the majority of differentially expressed genes (DEGs) upon aging, rather than a decrease [16, 30]. Commonly upregulated genes in aged BECs include those related to inflammation and the immune response (e.g., Vcam1, B2m, Cxcl12, Oas1) and members of the interferon (IFN) signaling pathway (e.g., Stat1, Bst2, Ifnar1, Gbp6 [16, 29, 31, 32]), in line with chronic inflammation as a well-established hallmark of aging [1]. These changes may be associated with age-dependent BBB dysfunction, as increased endothelial expression of pro-inflammatory cytokines has been linked with reduced expression of EC tight junction proteins [33] and BBB permeability [34]. Findings of decreased expression of classical BBB junction genes (e.g., Ocln, Cldn5) [30] in aged BECs are in line with this. In some studies, aged BECs also commonly exhibited upregulation of genes related to the hypoxia response (e.g., Hif1a, Ldha, Aldoa), and genes involved in oxidative stress pathways (e.g., Sod1, Alpl, Apoe), indicating an increased cellular stress response [16, 28]. Increased oxidative stress in BECs has been associated with age-induced impairment of the cerebral microvascular system and neurovascular uncoupling [13, 24, 35], age-related alterations of vascular reactivity [36] and reduced pericyte coverage of cerebral microvessels [37, 38], which could ultimately contribute to neuronal cell death [15]. Another common finding was the age-related upregulation of Vwf, a key regulator of hemostatic control, various Krüppel-like factors (e.g., Klf2, Klf4, Klf6), important for vasoprotection and response to injury, and genes involved in cell adhesion and/or extracellular matrix (ECM) organization (e.g., Adamts1, Itga6, Cyr61) [16, 28, 31]. Angiogenic signaling pathways, such as insulin-like growth factor-1 receptor (IGF-1R) , vascular endothelial growth factor (VEGF) and transforming growth factor beta (TGF-β),were also upregulated in aged BECs, indicated by the upregulation of genes such as Igf1r, Kdr, Flt1, Edn1, Eng, Id1, Bmpr2, and Acvrl1 [16, 28, 30, 31]. These findings are in contrast with the well-known phenomenon of capillary rarefaction in aging tissues [39], including the brain [40], suggesting that these genes may be activated in a counteracting attempt to restore the lost vascular density. Besides enhancing angiogenesis, VEGF signaling has however also been associated with BBB disruption [41, 42]. Aged BECs furthermore showed an upregulation of glycolytic genes (e.g., Pkm, Aldoa, Ldha), known to be important for angiogenesis, and which could indicate a shift toward glycolytic metabolism [16]. Altered metabolism is a well-known characteristic of EC aging and senescence (Box 1). Caution is nevertheless warranted, as Zhao and colleagues reported a selectively decreased expression for several metabolic genes involved in glucose/energy metabolism and ATP synthesis in aged brain capillaries [30]. Furthermore, Jin and colleagues reported decreased levels of Aplnr in aged mouse brain ECs, potentially impairing angiogenesis in healthy aging and after stroke [43]. BECs additionally exhibited an age-associated increase in protein synthesis, as demonstrated by an enrichment of genes involved in the proteostasis pathway [28] and the upregulation of ribosomal subunit genes, such as Rpl37, Rps20, and Rps21 [16, 31]. A slight age-associated increase of senescent BECs was furthermore reported in a scRNA-seq study of the mouse brain [26]. The accumulation of senescent ECs has been directly connected to BBB dysfunction in vitro and in vivo [44], at least in part via induction of the senescence-associated secretory phenotype [15], and the dysregulation of nitric oxide signaling in BECs [45].
Of note, age-related changes of the BEC transcriptomic profile were reversible by infusion with plasma derived from young mice [16] or by heterochronic parabiosis (a surgical procedure where young and old mice are joined together to share a common circulatory system) [28]. Here, capillary BECs were found to be the most susceptible cell type in the mouse brain to the aging or rejuvenating effects of old and young blood or plasma, respectively [16, 28]. Moreover, exercise has been shown to exert beneficial effects on brain microvascular perfusion [46, 47], and single-cell profiling revealed a putative endothelial-specific role in this phenomenon [27]. Precisely, subventricular zone cell types in the mouse brain were shown to interact with ECs via TWEAK signaling and this interaction was lost in aged mice, likely driven by a loss of Tnfrsf12a expression in ECs. Exercise was associated with the restoration of Tnfrsf12a expression in ECs, suggesting this signaling axis to be at least one of the underlying drivers of the beneficial effect of exercise on health of the brain vascular endothelium [27]. Lastly, glucagon-like peptide-1 receptor agonist treatment (known to alleviate various cellular phenotypes of aging) was also shown to improve BBB integrity and reverse, at least in part, aging-associated gene expression changes in brain ECs of aged mice [30]. This again showcases the adaptable nature of aging-related gene expression changes in the aged brain endothelium.
Aging-associated diseaseBased on recent scRNA-seq studies of Alzheimer’s disease (AD) and vascular dementia (VaD) in humans and mice, some of the genes that were upregulated in healthy aged BECs were also upregulated in AD- or VaD-associated ECs (e.g., APOE, ALPL, B2M, HSPA4 and VWF) [18, 48, 49]. Moreover, expression of the amino acid transporter Slc38a5, a finding unique to aged BECs in the mouse (compared to other aging tissue ECs) [31], was confirmed to be decreased in human brain tumors and vascular malformations as well [50], indicating a possible link between the transcriptomic profile of age-related brain diseases and aging. This transporter is predominantly expressed in ECs of a capillary(-venous) nature in the human brain [51], possibly in line with the increased susceptibility of murine microvascular BECs to the aging process.
MuscleSkeletal muscle is typically highly vascularized, yet vascular density is known to be adaptable and dependent on the energy demands of muscle tissue [
留言 (0)