
記住我


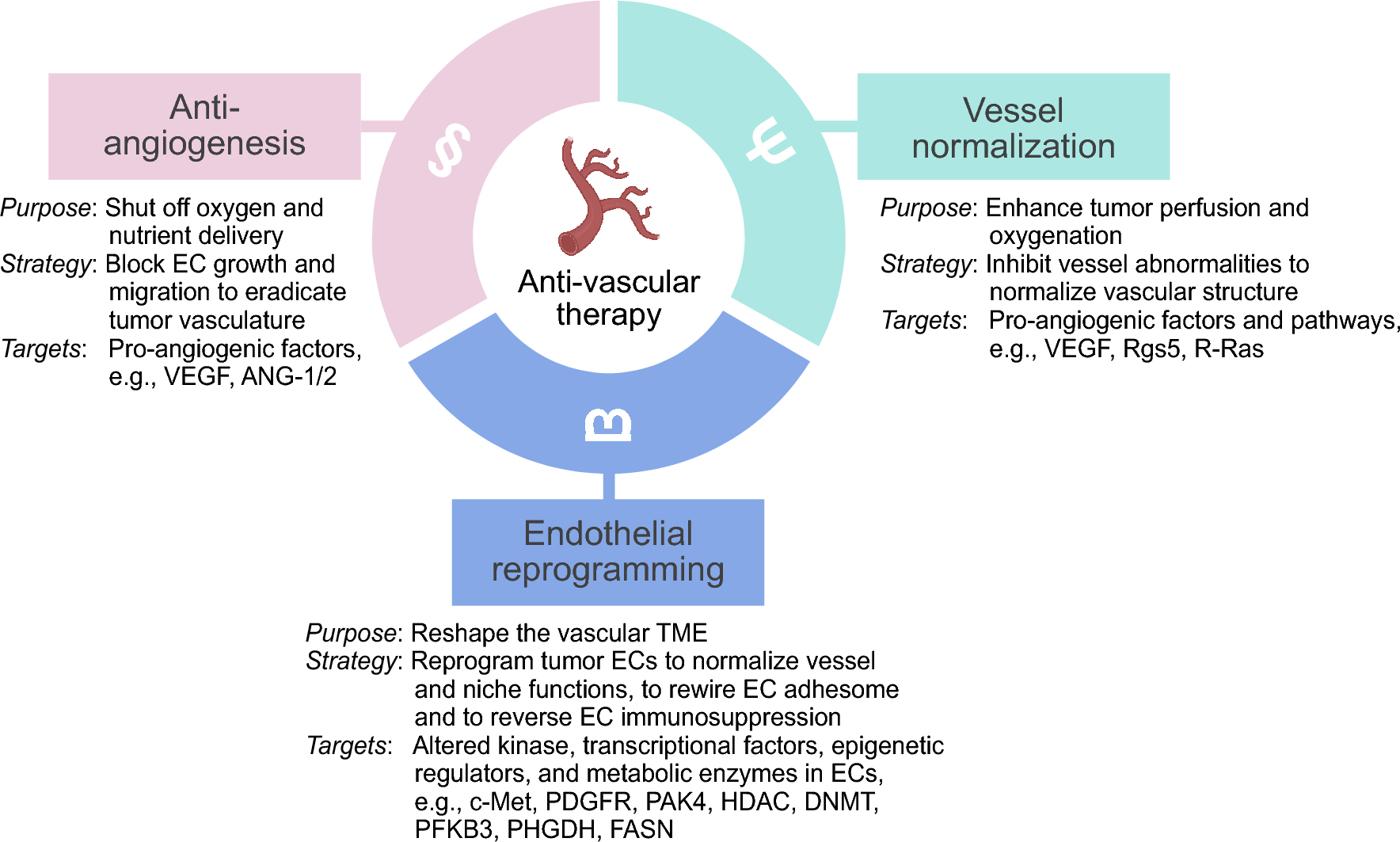
Control (CTL-H) and ALK1-mutated ECFCs (MUT-H) from newborns who have inherited a heterozygous ALK1 mutation from an HHT parent were clonally isolated from cord blood following the recommendations of the Vascular Biology Standardization Subcommittee [24]. Isolated ECFCs (3 CTL-H and 2 MUT-H, Table 1) displayed the classical endothelial cobblestone-like morphology (Fig. S1a) and were VE-cadherin (CD144) positive (Fig. S1b, c). These cells were also positive for the EC markers CD31 and CD146 and negative for the hematopoietic cell-specific surface antigen CD45 (Fig. S1d–f), confirming their endothelial identity. The functional activity of the two ALK1 mutants (MUT-H1; non sense mutation p.Trp141X and MUT-H2; missense mutation p.His280Asp, Table 1) was tested using the BMP response element (BRE) luciferase reporter assay in cells transfected with equal amounts of plasmids encoding either WT or mutated ALK1 (Fig. 1a), as previously described by our group [12]. Unlike cells transfected with wildtype ALK1, those exogenously expressing either of the two ALK1 mutants were unable to respond to a BMP9 stimulation (100 pg/mL, Fig. 1a), supporting that both studied mutations lead to loss of function.
Table 1 List of ALK1-mutated ECFCs and HUVECs isolated from HHT donorsFig. 1
BMP9 and BMP10 induce a similar transcriptomic response in control ECFCs. a Relative BMP Response Element (BRE) luciferase activity measured in NIH-3T3 cells overexpressing either WT or mutant ALK1 plasmids identified in ALK1-mutated ECFCs (p.Trp141X, MUT-H1) and (p.His280Asp, MUT-H2) used in the ECFC RNA-seq analysis. BRE firefly luciferase activities were normalized to renilla luciferase activity. Data shown are mean ± SEM from three independent experiments. b–f 3 CTL (CTL-H1, CTL-H2 and CTL-H3) and 2 ALK1-mutated ECFCs (MUT-H1 and MUT-H2) were stimulated or not with BMP9 or BMP10 (10 ng/mL) for 18 h. The experiment was repeated three times after which bulk RNA-seq analysis was performed. b Principal component analysis (PCA) showing clustering of RNA-seq samples by treatment (BMP9 or BMP10 stimulation vs NS) in CTL and MUT ECFCs. Each dot represents the mean of three experiments for one sample. c Scatter plot comparing log2 fold change (LFC) values of protein coding DEGs regulated in CTL ECFCs by BMP9 vs those regulated by BMP10 both compared to NS. Pearson correlation is reported. d, e Volcano plot representations showing global changes in gene expression in CTL ECFCs after BMP9 (d) or BMP10 (e) stimulation. DEGs with high LFC and high statistical significance are annotated. f Gene-set enrichment analysis (GSEA) performed using hallmark gene sets. The bar plot represents the top significant gene set categories enriched in CTL ECFCs upon BMP10 or BMP9-stimulation ordered using normalized enrichment scores (NES)
RNA-seq analysis in CTL ECFCs in response to BMP9 or BMP10 stimulationIn order to decipher the impact of ALK1-mutations on gene regulation in response to BMP9 and BMP10, RNA-sequencing was performed on ECFCs from 3 CTLs (CTL-H1–3) and 2 ALK1 mutated ECFCs (MUT-H1–2) (Table 1) that were either nonstimulated (NS) or stimulated with BMP9 or BMP10. To facilitate the detection of a high number of regulated targets and to better mimic physiological conditions, where circulating BMP9 and BMP10 are continuously replenished, cells were stimulated with BMP9 or BMP10 for a prolonged duration of 18 h. In line with that, as depicted in Fig. S2a, the mRNA expression of the BMP target ID1 increased with time (1–18 h) in response to BMP9 stimulation (10 ng/mL). Subsequently, having chosen to study gene regulation following a prolonged BMP9 or BMP10 stimulation, a relatively high BMP dose was required, as even the strong BMP target ID1 displayed significant upregulation only at 10 ng/mL in ECFCs (Fig. S2b). The experiment was repeated three times, after which RNA extraction for all samples was performed, followed by quality control, library preparation and RNA sequencing. Principal component analysis (PCA) could discriminate stimulated (BMP9 or BMP10, blue and red shapes, respectively) from NS conditions (grey shape) in all ECFCs but could not discriminate CTL ECFCs from ALK1-mutated ECFCs (Fig. 1b).
We first analyzed the BMP9 and BMP10 response versus (vs) NS condition in CTL ECFCs. Differential gene expression analysis using an absolute log2 fold change (LFC) threshold of 1 (lLFC(BMP response/NS condition)l ≥ 1) and an adjusted p-value (padj) ≤ 0.05 (Benjamini–Hochberg procedure; Supplementary information SI1) identified respectively 828 and 787 protein-coding differentially expressed genes (DEGs) upon BMP9 or BMP10 stimulation (Table 2). These DEGs were nearly equally distributed between up and down-regulated genes (Table 2). Interestingly, BMP9 and BMP10 induced a highly similar transcriptomic response evidenced by the high Pearson correlation coefficient of 0.993 when plotting the LFCs of the DEGs obtained in response to BMP9 vs those obtained in response to BMP10 (Fig. 1c) and the high number of shared DEGs regulated by both ligands (81.4%; Fig. S3a). This similarity was also reflected in the volcano plots of BMP9- and BMP10-stimulated vs NS ECFCs, which highlighted the same top dysregulated genes in terms of LFC or padj (Fig. 1d, e). Among these top DEGs, we detected many genes known to be regulated by BMP9 and BMP10 in other ECs (SMAD6, SMAD7, SMAD9, ID1, ID2, ENG, HEY2, BMPER, TMEM100, APLN) (Fig. 1d, e). We also identified new DEGs with very similar regulation patterns between BMP9 and BMP10, including STAT4, SLC40A1, SLC9A9, CLEC1B, CLEC3B, BMP4, KAT7, MPP4, HDDC2, CREB3L1, DRC1, GCHFR, and LEFTY2 (Fig. 1d, e). In accordance with the high similarity between BMP9 and BMP10 response, no DEGs could be identified when directly comparing BMP9 to BMP10-stimulated CTL ECFCs (Table 2). Additionally, gene set enrichment analysis (GSEA) using hallmark gene sets from MsigDB was performed independently on total genes (whether DEGs or not) regulated by BMP9 or BMP10 in CTL ECFCs (Supplementary information SI2). The top 20 enriched genesets identified were very similar between BMP9 and BMP10 (Fig. 1f). As expected, TGFß signaling was positively enriched by BMP9 and BMP10 stimulation, but we could also identify epithelial-mesenchymal transition, protein secretion and several signaling pathways (IL6-JAK-STAT3, KRAS and MTORC1) as positively enriched terms (Fig. 1f). On the other hand, many hallmarks related to cell cycle (MYC-targets, mitotic spindle, G2M checkpoint and E2F targets) were negatively enriched (Fig. 1f), supporting the reported role of BMP9 and BMP10 in maintaining vascular quiescence [8, 25]. Together, these results demonstrate that under these stimulatory conditions, BMP9 and BMP10 induce a very similar transcriptomic response in CTL ECFCs.
Table 2 Number of protein-coding DEGs identified by differential expression analysis in CTL and ALK1-mutated ECFCs and HMVECs ALK1 heterozygosity in ECFCs does not impair the global transcriptomic response to BMP9 or BMP10To uncover the effect of heterozygous ALK1 mutations on gene regulation, we analyzed the basal transcriptome of MUT versus CTL ECFCs, before any stimulation, and in response to BMP9 or BMP10. In accordance with the PCA, which could not differentiate CTL from MUT ECFCs (Fig. 1b), only 28 DEGs were identified between NS CTL and NS MUT ECFCs (Table 2). Similarly, following a BMP9 or BMP10 stimulation, only 19 and 30 DEGs, respectively, were significantly differentially expressed between the two ECFC groups (Table 2).
Upon analyzing the transcriptomic response of MUT ECFCs to BMP9 or BMP10, 604 and 564 DEGs were identified, respectively (Table 2). As in CTL ECFCs, BMP9 and BMP10 induced a similar response in ALK1-mutated cells, with 77.2% DEGs commonly regulated by both ligands (Fig. S3b) and a highly similar global regulation pattern (Pearson correlation coefficient = 0.989, Fig. S3c).
We next compared the LFCs of each list of target genes in stimulated CTL vs stimulated ALK1-mutated ECFCs and found that most regulated genes demonstrated the same regulation patterns in the two cell groups upon BMP9 or BMP10 stimulation (Pearson correlation coefficient = 0.958 and 0.962 respectively, Fig. 2a, b). Consistently, we could identify in ALK1-mutated ECFCs the same top protein-coding DEGs upon BMP9 or BMP10 stimulation as in CTL ECFCs (SMAD6, SMAD7, SMAD9, ID1, ID2, ENG, HEY2, BMPER, TMEM100, STAT4, APLN, SLC40A1, SLC9A9, CLEC1B, CLEC3B, BMP4, KAT7, MPP4, HDDC2, CREB3L1, DRC1, GCHFR, LEFTY2; Fig. 2c, d). Altogether, these data unexpectedly show that CTL and ALK1-mutated ECFCs display highly similar transcriptomic profiles following BMP9 or BMP10 stimulation.
Fig. 2
ALK1 heterozygosity in ECFCs does not impair the global transcriptomic response to BMP9 or BMP10. a–d 3 CTL (CTL-H1, CTL-H2 and CTL-H3) and 2 ALK1-mutated ECFCs (MUT-H1 and MUT-H2) were stimulated or not with BMP9 or BMP10 (10 ng/mL) for 18 h. The experiment was repeated three times after which bulk RNA-seq analysis was performed. a, b Scatter plots comparing log2 fold change (LFCs) values of protein coding DEGs regulated by BMP9 (a) or BMP10 (b) in CTL ECFCs vs MUT ECFCs. Pearson correlation is reported. c, d Volcano plots representations showing global changes in gene expression in ALK1-mutated ECFCs after BMP9 (c) or BMP10 (d) stimulation vs NS. DEGs with high LFC and high statistical significance are annotated
ALK1 heterozygosity in ECFCs does not impair p-Smad1/5 response to BMP9To understand this surprising result, we next investigated the status of Smad1/5 phosphorylation induced by BMP9 in ALK1-mutated (n = 6) vs CTL ECFCs (n = 4). This analysis included the previously described ECFC clones that were studied by RNA-seq, as well as subsequently isolated clones from four additional newborns with different ALK1 mutations and one additional CTL (Table 1; Fig. S4). We focused on BMP9 regulation only, as BMP9 and BMP10 showed similar transcriptomic responses (Fig. 1c). ALK1 MUT ECFCs stimulated with BMP9 (10 ng/mL) for 1 h displayed similar levels of nuclear p-Smad1/5 immunofluorescence intensities to stimulated CTLs (Fig. 3a, b). To validate this result, we performed a BRE luciferase reporter assay in CTL (n = 4) and MUT (n = 6) ECFCs (Table 1; Figs. 1a, S4) stimulated with increasing doses of BMP9 (0.2–10 ng/mL) for 6 h. Both CTL and MUT ECFCs exhibited a clear dose-dependent response to BMP9 (Fig. 3c); yet, no significant differences were observed between the two groups (Fig. 3c), which displayed identical half maximal effective concentrations (EC50: 376 pg/mL for CTL vs 404 pg/mL for MUT ECFCs; inset Fig. 3c). We also quantified the mRNA levels of ID1, a target gene known to be strongly induced by BMP9, and found no difference between CTL (n = 4) and MUT (n = 4) ECFCs (Table 1; Fig. 3d). To examine whether the similar Smad1/5 activation in MUT vs CTL ECFCs was due to a compensation in ALK1 protein levels in ALK1-mutated ECFCs, we assessed the levels of membranous ALK1 by flow cytometry analysis in five different MUT ECFCs (Table 1) in comparison to 3 CTL ECFCs. We found that ALK1-mutated ECFCs carrying a missense mutation had a slightly lower level of membranous ALK1 level than CTL ECFCs, and those carrying an ALK1 nonsense mutation displayed a 50% reduction in the level of cell surface ALK1 (Fig. 3e, f). Thus the similar Smad signaling response in CTL and ALK1-mutated ECFCs cannot be attributed to an elevated compensatory level of ALK1 in mutated ECFCs. Altogether, these results support that ALK1-mutated ECFCs maintain intact activation of the canonical Smad1/5 signaling pathway in response to BMP9 despite their reduced cell surface ALK1 levels.
Fig. 3
ALK1 heterozygosity in ECFCs does not impair the p-Smad1/5 response to BMP9. a, b 3 CTL and 6 MUT ECFCs (MUT-H1–H6) were stimulated with BMP9 (10 ng/mL) for 1 h then fixed and immunostained for phospho-Smad1/5 (p-Smad1/5). Cells were stimulated in duplicates and at least 16 different fields were imaged in each well. a Representative p-Smad1/5 immunostainings in 1 CTL and 1 MUT ECFC in the absence or presence of BMP9 for 1 h. The nuclei were counterstained using Hoechst 33342. b p-Smad1/5 fluorescence was quantified in the nuclei using IN Carta Image Analysis Software. Data presented are mean relative fluorescent intensity (RFU) ± SEM of three independent experiments. c 4 CTL and 6 MUT (MUT-H1–6) ECFCs were transiently transfected with pGL3(BRE)2-luc and pRL-TK-luc. Cells were then either non-treated or stimulated with increasing concentrations of BMP9 (0.2, 0.5, 1, 10 ng/mL) for 6 h. Firefly luciferase activities were normalized to renilla luciferase activities. Data shown are mean ± SD from 1 representative experiment of 4, and each point corresponds to one donor. The inset represents the calculated BMP9 EC50 for CTL and MUT ECFCs. d RT-qPCR quantification of ID1 mRNA expression normalized to HPRT level in 4 CTL and 4 MUT (MUT-H1–H4) ECFCs following an 18 h stimulation with 10 ng/mL BMP9. Data are mean ± SEM of three independent stimulations presented as ΔΔCT compared to CTL NS. e Flow cytometric analysis comparing cell-surface ALK1 levels in 3 CTL vs 5 MUT ECFCs either carrying an ALK1 missense mutations (MUT-H2, MUT-H4 and MUT-H6) or a nonsense mutations (MUT-H1 and MUT-H3). Isotypic control is illustrated in grey. One representative flow cytometry histogram of 3 is shown. f Quantification of the ALK1 cell-surface expression by flow cytometry (in percentage of in MUT ECFCs with missense or nonsense mutations compared to CTL ECFCs). Data are means ± SEM of three independent experiments. Two-way Anova followed by Sidak’s multiple comparisons tests were used for statistical analysis of b–d, except for the inset of c where Mann–Whitney test was used. Kruskal–Wallis test followed by Dunn’s multiple comparison’s test was used for f. For all panels, ns non-significant, *P < 0.05, **P < 0.01, ***P < 0.001 and ****P < 0.0001
PAH patient-derived lung ECs carrying ALK1 mutations display different transcriptomic profiles compared to CTLs at the basal stateHaving not detected any differences in the transcriptomic nor in the early p-Smad1/5 response to BMP9 or BMP10 in newborn-derived ALK1-mutated ECFCs, we investigated the transcriptomic response in HMVECs carrying ALK1-mutations derived from transplanted sick lungs of two PAH patients (MUT-P1 and -P2, Table 3). The two ALK1 mutations were missense mutations mapping to the kinase domain of ALK1 and were both confirmed as LOF mutations using the BRE luciferase assay (Fig. 4a).
Table 3 PAH patient characteristicsFig. 4
ALK1-mutated HMVECs display substantially different transcriptomic profiles compared to controls. a Relative BMP Response Element (BRE) luciferase activity measured in NIH-3T3 cells overexpressing either WT or mutant ALK1 plasmids identified in ALK1-mutated HMVECs (p.Gly319Arg, MUT-P1) and (p.Arg484Trp, MUT-P2) that are included in the HMVEC RNA-seq analysis. BRE luciferase activities were normalized to renilla luciferase activities. Data shown are mean ± SEM from of three independent experiments. b–f 3 CTL and 2 ALK1-mutated (MUT-P1 and MUT-P2) HMVECs were stimulated or not with BMP9 or BMP10 (10 ng/mL) for 18 h. The experiment was repeated three times after which bulk RNA-seq analysis was performed. b GSEA performed using hallmark gene sets. The bar plot represents the top significant gene set categories enriched in non-stimulated MUT HMVECs compared to nonstimulated (NS) CTL HMVECs. Each bar represents a hallmark gene set and bars are ordered from top to bottom by decreasing order of enrichment score (NES). c, d Volcano plot representations showing global changes in gene expression in CTL HMVECs after BMP9 (c) or BMP10 (d) stimulation. DEGs with high LFC and high statistical significance are annotated. DEGs annotated in blue correspond to DEGs identified in CTL ECFCs (Fig. 1d, e). e, f Scatter plots comparing log2 fold change (LFCs) of DEGs regulated by BMP9 (e) or BMP10 (f) in CTL HMVECs vs MUT HMVECs. Pearson correlation is reported
To delineate the transcriptomic signature of these cells, RNA-sequencing was performed, using the same experimental conditions described for ECFCs, i.e. using 3 CTLs and 2 ALK1-mutated HMVECs from PAH patients (MUT-P1 and -P2, Table 3) that were either stimulated overnight with BMP9 or BMP10 (10 ng/mL) or were left nonstimulated (NS). Hierarchical clustering identified four separate clusters: (1) NS and stimulated MUT-P2 samples, (2) NS and stimulated MUT-P1 samples, (3) NS CTLs and (4) BMP9 and BMP10-stimulated CTL samples (from left to right, Fig. S5). This clustering showed that, unlike MUT-H ECFCs, the transcriptomes of MUT-P HMVECs were clearly different from those of CTLs. Interestingly, MUT-P HMVECs from each patient were separated into two different clusters (Fig. S5), highlighting variability between the two patients, which might be related to different disease stage or different treatments. The differential expression analysis (lLFCl ≥ 1 and padj ≤ 0.05, Benjamini–Hochberg correction; Supplementary information SI3) comparing NS ALK1-mutated and NS CTL HMVECs identified 1261 protein-coding DEGs (Table 2), with both down-regulated (58%) and up-regulated (42%) genes. GSEA using hallmark gene sets in NS CTL vs ALK1-mutated HMVECs (Supplementary information SI4) revealed a positive enrichment of several gene sets involved in cell cycle (E2F targets, G2M checkpoint, MYC-targets and mitotic spindle) and a negative enrichment in genes related to various signaling pathways (KRAS signaling, WNT-beta-catenin signaling and TNFα signaling; Fig. 4b). Together, these data show that ALK1-mutated HMVECs derived from diseased lungs have a severely altered basal transcriptome compared to CTL HMVECs.
BMP9 and BMP10 induce cell type-specific transcriptomic reponsesWe then analyzed the BMP9 and BMP10 transcriptomic response in CTL HMVECs. Differential expression analysis identified 704 and 481 DEGs in BMP9- and BMP10-stimulated CTL HMVECs, respectively (Table 2). Although the number of DEGs was lower in BMP10 stimulated cells, the gene regulation patterns in BMP9- vs BMP10-stimulated CTL HMVECs were highly correlated (Pearson correlation coefficient = 0.844), further supporting, as for ECFCs (Fig. 1c), that BMP9 and BMP10 induce very similar transcriptomic responses in vitro under these experimental conditions. This similarity was also reflected in the volcano plots of BMP9- and BMP10-stimulated vs NS cells, which highlighted similar top dysregulated genes in terms of LFC or padj (Fig. 4c, d). Interestingly, by comparing BMP9/10-regulated genes in CTL HMVECs to those in CTL ECFCs, with the limitation that these data were derived from two independent RNA-sequencing, only a quarter of the DEGs was shared between ECFCs and HMVECs (27% for BMP9 and 26% for BMP10; Fig. S6a, b). ID1, SMAD6, SMAD7, ENG, KAT7, CLEC1B, CLEC3B, and APLN were identified as top targets in both ECFCs (Fig. 1d, e) and HMVECs (highlighted in blue in Fig. 4c, d). On the other hand, some top target genes were specific to HMVECs (e.g. LOX and NOG) (highlighted by an asterisk in Fig. 4c, d).
ALK1-mutated HMVECs display slightly different transcriptomic responses to BMP9 and BMP10 compared to CTL HMVECsWe next analyzed the BMP9 or BMP10 response in ALK1-mutated vs CTL HMVECs. BMP9 and BMP10 stimulation resulted in the regulation of 295 and 206 DEGs, respectively (Table 2). This corresponds to around 60% less DEGs compared to stimulated CTLs. This decrease in the number of DEGs suggests that ALK1-mutated HMVECs might possess a reduced capacity to respond to BMP9 or BMP10. When comparing the LFCs of each list of dysregulated genes in stimulated CTL vs MUT HMVECs, we obtained fairly high Pearson correlation coefficients (0.728 for BMP9 and 0.855 for BMP10; Fig. 4e, f). However, these correlations were lower than the ones obtained in ECFCs (Fig. 2a, b), suggesting that the BMP9 and BMP10 responses in ALK1-mutated HMVECs might be more affected than in ALK1-mutated ECFCs. Since we are simultaneously analyzing two variables (genotype and stimulation), and because there was a clear difference between CTL and MUT HMVECs at the basal level (Table 2), we performed a two-factor analysis as previously described [26] (See Materials and Methods section in Supplementary information) considering the two factors: genotype and BMP9 or BMP10 treatment (Supplementary information SI5). We found 44 protein-coding interaction term genes differentially regulated by BMP9 in CTL versus ALK1-mutated HMVECs and 15 in response to BMP10, which were all shared with BMP9 stimulation (marked by an asterisk, Benjamini–Hochberg adjusted for multiple comparisons, padj < 0.05; Fig. S7a). On the other hand, when applying the same kind of analysis on CTL and MUT ECFCs, we could not detect any interaction term genes, in accordance with the highly similar transcriptomic responses to BMP9/BMP10 between MUT and CTL ECFCs. Then, we hypothesized that this difference between MUT HMVECs and MUT ECFCs could be due to a further reduction in ALK1 level in MUT HMVECs below a critical threshold. Nevertheless, flow cytometric analysis revealed similar cell-surface ALK1 levels in MUT HMVECs compared to their control counterparts (Fig. S8).
Out of the 44 interaction term genes identified in MUT HMVECs, we selected 25 genes (highlighted in blue in Fig. S7a) for validation by RT-qPCR on independent BMP9 stimulations. The selection was based on genes showing only minimal interindividual heterogeneity between members of each group (CTL or MUT group; data not shown). The regulation trend of each of the 25 selected genes was confirmed by RT-qPCR (Fig. S7b), validating the robustness of the two-factor analysis.
LFNG (lunatic fringe) shows impaired regulation by BMP9 in ALK1-mutated HMVECs, ECFCs and HUVECsAmong the 25 genes validated, we focused on 6 (LFNG, JAG2, TNFRSF1B, SLC6A6, SOX13 and CEBPG) that were also identified as DEGs in response to BMP9 and BMP10 in CTL ECFCs with the same sense of regulation, making them good candidates to compare the effect of ALK1 mutation on gene regulation by BMP9 across the different EC models. The BMP9 regulation of all 6 genes was confirmed by RT-qPCR in CTL HMVECs (Figs. 5a, S9a), and consistent with the two-factor analysis, their induction was repressed in ALK1-mutated HMVECs, except for SLC6A6, whose difference between CTL and MUT HMVECs did not reach statistical significance (Fig. S9a). Due to the limited number of accessible PAH donors with ALK1 mutations, and to test whether LOF mutations in another component of the receptor complex would affect the response similarly, we tested BMPR2-mutated HMVECs derived from transplanted lungs of 3 PAH patients (Table 3), and found comparable regulation patterns to ALK1-mutated HMVECs for all 6 genes (Figs. 5a, S9a).
Fig. 5
Regulation of LFNG mRNA expression by BMP9 in CTL and ALK1-mutated HMVECs, ECFCs and HUVECs. a, b RT-qPCR quantification of the mRNA expression level of LFNG in 3 CTL, 2 ALK1-mutated (MUT-P1 and -P2) and 3 BMPR2-mutated HMVECs (MUT-P3–P5) (a), 3 CTL and 4 ALK1-mutated ECFCs (MUT-H1–H4) (b). LFNG mRNA expression level was normalized to HPRT mRNA expression and presented as ΔΔCt compared to mean CTL NS. Data shown are mean ± SEM of at least three independent stimulations. c Count plot representation showing the regularized log transformed counts of LFNG mRNA in CTL-P and ALK1-MUT-P HMVECs in nonstimulated (NS) and BMP9 or BMP10 stimulated cells. d Western blot analysis of 3 CTL (1–3) and 2 ALK1-mutated HMVECs (MUT-P1–2) that were either NS or stimulated for 24h with 10 ng/mL BMP9. Cell lysates were resolved by 4–20% SDS–PAGE and immunoblotted with antibodies against lunatic fringe or against HSP90 (loading control). The resulting blots are shown along with quantification of the mean lunatic fringe signal normalized to HSP90. e RT-qPCR quantification of the mRNA expression level of LFNG in 3 CTL (CTL-H′1–3) and 3 ALK1-mutated (MUT-H′3–5) HUVECs. LFNG mRNA expression level is normalized to HPRT mRNA expression and presented as ΔΔCT compared to mean CTL NS. Data shown are mean ± SEM of at least three independent stimulations. a, b, e, Two-way Anova followed by Sidak’s multiple comparisons test were used for statistical analysis of panels. ns non-significant, **P < 0.01 and ****P < 0.0001 vs NS and ##P < 0.01 and ####P < 0.0001 vs CTL. f 2 CTL HMVECs were treated either with scrambled siRNA (siScr) or two different concentrations of siRNA against ALK1 (siALK1 a and b) to generate a gradient of ALK1 expression and then stimulated with 10 ng/mL BMP9 for 18 h. LFNG mRNA expression normalized to HPRT mRNA level is presented as 2−ΔCt. Data shown are mean ± SD of 2 CTL HMVECs. Inset represents ALK1 mRNA expression presented as ΔΔCt compared to scrambled siRNA-transfected cells
Then, we tested the BMP9 regulation of these genes in CTL and ALK1-mutated ECFCs (n = 4) using RT-qPCR. BMP9 stimulation upregulated the mRNA expression levels of all six genes in CTL ECFCs (Figs. 5b, S9b). In accordance with the results of the two-factor analysis in ECFCs, the regulation of the aforementioned genes in ALK1-mutated cells showed either similar or slightly weaker regulation by BMP9 compared to CTLs (Fig. S9b), with the exception of LFNG (Fig. 5b). In HMVECs, using RT-qPCR, LFNG mRNA was strongly upregulated by BMP9 in CTLs (10.3 folds), but this upregulation was strongly reduced in ALK1-mutated (2.7 folds) and BMPR2-mutated HMVECs (2.6 folds; Fig. 5a), mirroring the result of the two-factor analysis generated from the RNA-seq data in these cells, as illustrated in the count-plot representation (Fig. 5c). The dysregulation of LFNG was also validated at the protein level following a 24 h stimulation of 10 ng/mL BMP9 in CTL and ALK1-mutated HMVECs (6.49 folds in CTL vs 1.17 folds in ALK1-mutated HMVECs; Fig. 5d). In ECFCs, while CTL cells displayed a significant upregulation of LFNG expression by BMP9 (5.1 folds), ALK1-mutated ECFCs presented a significantly weakened upregulation of LFNG (3.1 folds, n = 4; Fig. 5b). Together, these results support the hypothesis that LFNG transcriptional regulation by BMP9 could be affected by ALK1 heterozygosity, but did not allow to conclude on the other interaction term genes studied.
To further test this hypothesis, we studied the BMP9 response in human umbilical vein endothelial cells (HUVECs), another type of ECs that can be isolated from newborns. RT-qPCR for LFNG, JAG2, TNFRSF1B, SLC6A6, SOX13 and CEBPG were performed on 3 CTLs and 3 LOF ALK1-mutated HUVECs (CTL-H and MUT-H) stimulated or not with BMP9 for 18 h. These genes were all induced in response to BMP9 in CTL HUVECs, although SOX13 stimulation did not reach significance (Figs. 5e, S9c). As for HMVECs and ECFCs (Fig. 5a, b), we found a significant decrease in the level of induction of LFNG mRNA expression by BMP9 in ALK1-mutated HUVECs compared to CTLs (FC = 4.9 in CTL-H′ and 2.6 in MUT-H′; Fig. 5e). The BMP9 regulation of JAG2, TNFRSF1B and SLC6A6 mRNA expressions were also found to be significantly reduced in ALK1-mutated vs CTL HUVECs (Fig. S9c).
To directly relate the BMP9-induced upregulation of LFNG to functional ALK1 levels, we tested LFNG induction in CTL HMVECs whose ALK1 expression level was reduced using an siRNA approach. Reducing ALK1 mRNA levels in CTL HMVECs by 50 or 90% (siALK1 a and siALK1 b, respectively; inset in Fig. 5f) suppressed LFNG induction by BMP9 in a dose-dependent manner (Fig. 5f).
Together, these data show that ALK1 heterozygosity could impair the BMP9 regulation of the mRNA expression of LFNG and possibly other genes, depending on the endothelial cellular model.
留言 (0)