
記住我

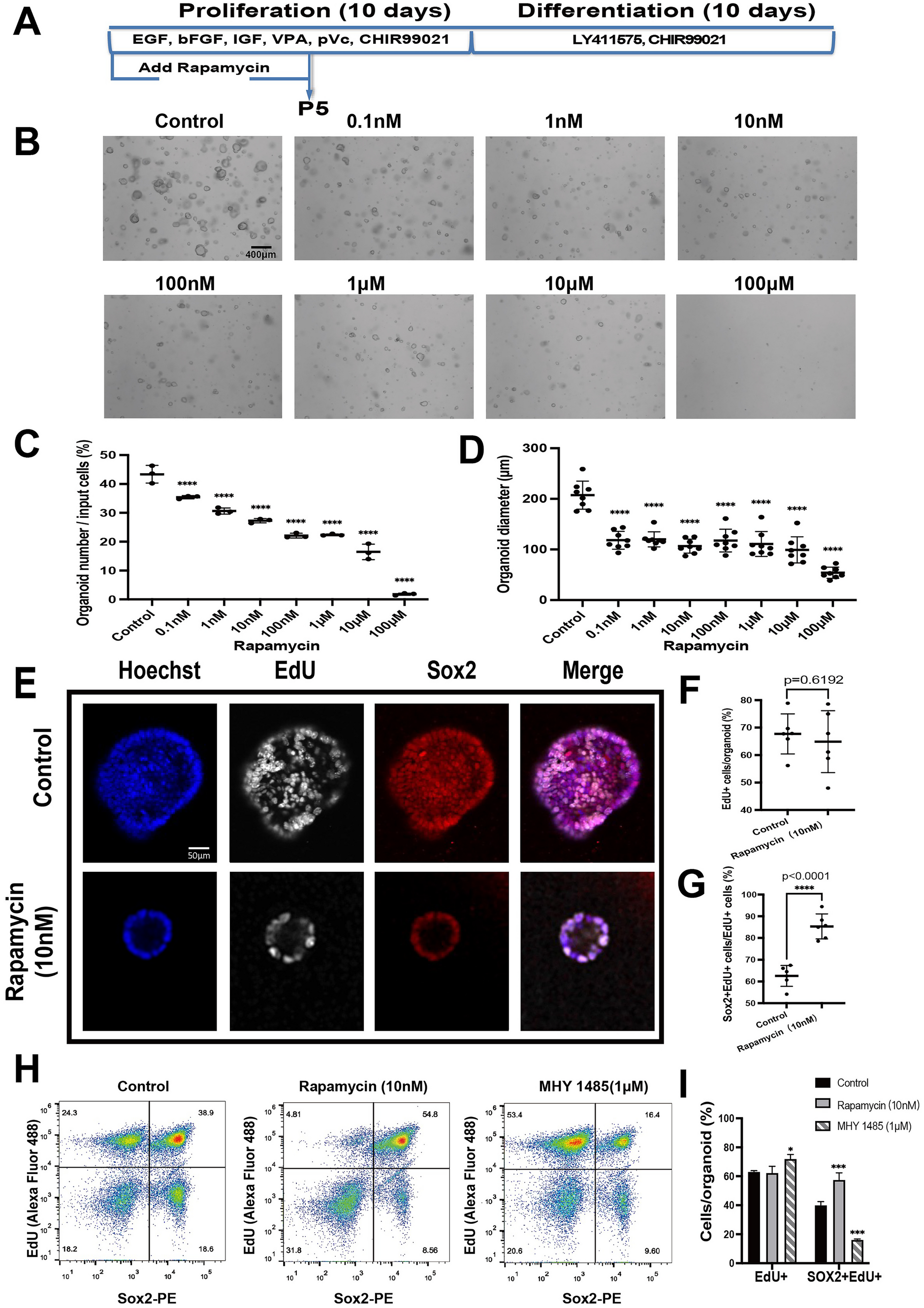
In this appendix, we take a closer look at the convolution equations (Eqs. 1 and 2), since the assumptions justifying these formulas were implicit in the main body of the paper.
We first start by considering the equation for the non-masked version of the CAP (Eq. 1), recalled here:
$$\begin CAP \, (t) = E *u_0 \, (t) \, . \end$$
This formula can in fact correspond to two different approaches, depending on how the unitary response is defined. If the unitary response is the same as the spike unit response, then \(E\) is the compound post-stimulus time histogram (PSTH) of all ANFs. This is consistent with several studies that use elaborate computational models of ANF activity to simulate compound PSTHs and then generate CAP waveforms [20, 46]. This differs however from our approach, as we actually never try to reproduce the compound PSTH. In this sense, our work is closer to a second approach [47, 48], where \(E\) is defined as an excitation spread over latencies with a one-to-one mapping of latencies and CFs. This approach necessarily requires that the spike-time jitter of ANFs tuned to a given CF is encompassed in the unitary response since it cannot be included in the excitation pattern.
To make the above distinction more explicit, let us consider \(nPST\), the compound PSTH of ANFs tuned to a single CF normalized by the total number of spikes. We also assume that only a limited segment of the cochlear partition contributes to the CAP waveform, so that \(nPST\) can be considered independent of CF. We can then write a double convolution equation for the CAP waveform:

(6)
where \(nPST, cPST\) stand for the normalized and compound PSTHs, and \(UR\) is the spike unit response. In this formula, we have used notations similar to Bappert et al. [48], which describes the double convolution approach in more detail. If \(nPST\) is left outside the function to the right of the convolution, the unitary response aligns with the spike unit response, corresponding to the first approach of simulating the compound PSTH. In our case, however, the unitary response \(u_0\) takes into account the spike unit response as well as the normalized spike histogram \(nPST\).
Now let us take a closer look at the equation for the masked version of the CAP, specifically the masking-release \(\Delta CAP(t)\), which is the real focus of this paper. Since we consider masking releases associated with the manipulation of a narrow spectral notch, the assumption that only a limited segment of the cochlear partition contributes to \(\Delta CAP(t)\) is always justified. We can therefore approximate \(\Delta CAP(t)\) with an equation similar to Eq. 6:

(7)
We recall that \(R(\tau )\) is the masking-release pattern and \(u\) is the unitary response – here, the zero subscript has been removed to distinguish the unitary response from the one in Eq. 6, defined differently. Again, \(u\) is considered as the compound of the spike unit response and the difference in the PSTH of a population of synchronized ANFs normalized with respect to the amount of masking (\(n\Delta PST\)). If, on the other hand, the unitary response was identical to the spike unit response \(UR\), the function to the left of the convolution would be the difference in the compound PSTH induced by masking (\(c\Delta PST\)). We are not interested, however, in the actual decomposition of \(u\), justifying that the simpler equation \(\Delta CAP = R *u\) is kept in the main body of the paper. It is worth noting that, in this equation, we assume that \(n\Delta PST\) is independent of the amount of masking. If a click probe of medium-to-high intensity is used, the individual PSTHs are characterized by a sharp predominant peak restricted on a short time interval [29]. As a result, the changes in the shape of \(n\Delta PST\) are expected to have a minimal effect on the CAP; however, the amount of masking applied to the PSTHs will have a significant impact on the CAP amplitude. Prior to any experiment, we tested whether this hypothesis was reasonable with a well-established computational model of ANF responses (BEZ model [38]). This analysis is left as supplementary material (Online Resource 1). As for the spike unit response \(UR\), studies have typically reported that it can be considered independent of the ANF best frequency or spontaneous rate [28, 49, 50].
Appendix B: Gammatone ModelThis appendix contains the computation of the average intensity at the output of a gammatone cochlear filter:
Note: In this paragraph, \(\tau\) does not have the same use as in the main part of the paper where it is a variable for latencies. Here, it refers to the time constant of the gammatones.
The k-th order gammatone, characterized by an envelope proportional to \(t_+^ e^\), is defined in the frequency domain (complex version, w.l.o.g.) by
$$\begin |w_(\omega )|^2 = 2k-2\\ k-1\end}\right) }^ 2^ \tau \left[ 1+\tau ^2 (\omega - 2\pi \, CF )^2\right] ^ \, . \end$$
The average quadratic response considering a single-band Gaussian noise masker is
$$\begin < A^2 > = S_0 2k-2\\ k-1\end}\right) }^ 2^ \pi ^ \tau \, \int _}-CF)}^}-CF)} \left[ 1+\tau ^2\omega ^2\right] ^ d\omega \end$$
$$\begin < A^2 > = S_0 2k-2\\ k-1\end}\right) }^ 2^ \pi ^ \, \int _( 2\pi \tau (f_}-CF))}^(2\pi \tau (f_}-CF))} \cos ^\theta \, d\theta \ . \end$$
The last integral is then computed by writing
$$\begin \cos ^\theta = 2^ \cdot \left[ \sum _^ 2k-2\\ l\end}\right) } 2 \cos ( (2k-2-2l) \theta ) + 2k-2\\ k-1\end}\right) } \right] . \end$$
In the case of a masker presenting multiple bands, the expressions for each band simply add up.
Note: The 10-dB bandwidth is related to \(\tau\) by \(BW_ \tau \pi = \left[ 10^ - 1 \right] ^\).
留言 (0)